Full Length Research Paper

ABSTRACT
The reddish egret (Egretta rufescens) is the rarest heron in North America and much remains to be learned about in the ecology of the species. The reddish egret is a foraging habitat specialist and relies on shallow coastal ecosystems. There is a paucity of information on foraging habitat requirements and the availability of foraging habitat throughout the annual cycle. Characteristics of foraging habitat at locations within the Laguna Madre, Texas where reddish egrets were observed foraging were measured. These characteristics were used to conduct a geospatial analysis that estimated the spatial and temporal distributions of foraging habitat in the Laguna Madre across 120 weeks from 2012 to 2014. Reddish egrets (n = 372) foraged in an average water depth of 10.1 ± 0.68 cm and in areas with average seagrass cover of 12.3 ± 2.74%. Approximately, 75 000 ha of foraging habitat were delineated to be available in the Laguna Madre across the study period; of this, 4 003 ha were available ≥ 50% of the time. The amount of available foraging habitat was relatively high during the spring and summer, and decreased by ~50% during winter. This model-based approach can be used throughout the species’ range to examine foraging habitat availability which is a current conservation need according to the Reddish Egret Conservation Action Plan.
Key words: Egretta rufescens, foraging habitat, Laguna Madre, reddish egret, Texas.
INTRODUCTION
The reddish egret (Egretta rufescens) is the rarest Ardeidae in North America and is highly restricted geographically (International Union for Conservation of Nature, 2012). It is a coastal wetland specialist found along the Gulf of Mexico, the Caribbean and Bahamas, the Atlantic Coast of Florida, and the Pacific Coast of Mexico (Lowther and Paul, 2002). The reddish egret was nearly extirpated early in the 20th century due to plume hunting, and today there is an estimated 2000 breeding pairs in the United States with nearly half occurring in Texas (Lowther and Paul, 2002; Green, 2006). Because of its narrow habitat requirements and limited distribution, the reddish egret was listed as a species of concern by the U.S. Fish and Wildlife Service. It is also listed as near threatened on the International Union for Conservation of Nature Red List (2012).
Foraging habitat of the reddish egret typically consists of shallow coastal flats and lagoons where it actively forages for small fish such as sheepshead minnow (Cyprinodon variegatus), longnose killifish (Fundulus similis) and pinfish (Lagodon rhomboids) (Lowther and Paul, 2002). Because the reddish egret is restricted to coastal areas, it is sensitive to disturbances and alterations within these habitats. The rapidly growing human population along the Texas Coast has had major impacts on coastal habitats (Moulton et al., 1997). The Gulf Intracoastal Waterway is frequently dredged and new sites are needed for dredge disposal. Placement of dredge material can alter tidal flat habitats by covering seagrass meadows that provide habitat for fish, impacting water quality, and changing the hydrology of foraging areas for waterbirds (Farmer, 1991). Therefore, an understanding of the spatial and temporal dynamics of foraging habitat will enable managers to reduce negative impacts to these areas and support foraging requirements for this unique species. In fact, the recently developed Reddish Egret Recovery Plan (Wilson et al., 2014) specifically states the identification of key foraging areas as one of three primary goals. Consequently, the goal of this study was to develop a spatial model that delineates the temporal distribution of foraging habitat for the reddish egret in the Laguna Madre of Texas and to test the performance of the model with an independent sample of locations of foraging reddish egrets marked with GPS equipped satellite transmitters.
METHODOLOGY
Study area description
The Laguna Madre is located along the lower Texas Coast (Figure 1) and supports the largest concentration of reddish egrets in North America (Green, 2006). It is a large, hypersaline lagoon stretching ~185 km in length with an average width of 7 km, and relatively shallow water depths, averaging 1 m deep (Tunnell, 2002). Tides in the Laguna Madre are mainly wind-driven and can cause water levels to fluctuate by 0.6 to 0.9 m (Tunnel, 2002). Seagrass meadows and wind tidal flats are the primary habitat types found in the Laguna Madre. Shoalgrass (Halodule wrightii) and manateegrass (Cymodocea syringodium) are the dominant species in the upper and lower Laguna, respectively (Quammen and Onuf, 1993). Islands are abundant and widespread throughout the Laguna Madre, most of which were formed by placement of dredge material during construction and routine maintenance dredging of the Intracoastal Waterway (Tunnell, 2002). Some of these islands have become occupied by colonial nesting waterbirds, including the reddish egret.
Foraging observations
Habitat of foraging reddish egrets observed throughout the annual cycle (about every 2 weeks) from March 2008 to April 2010 were sampled. The Laguna Madre was divided into six zones for sampling purposes, and each survey day we systematically searched a zone by boat for foraging reddish egrets. Foraging individuals were observed for 20 min or until foraging ceased. Observations were made from >150 m to reduce any influence on foraging individuals. Color morph and age (hatch-year or after hatch-year) were recorded for each individual observed. Plumage characteristics and bill color were used to identify juvenile birds (Cézilly and Boy, 1988; Farmer, 1991). Dark morph hatch-year individuals are mostly gray with little if any distinction in color between the head and neck, and the rest of the body. The plumage of white morph hatch-year birds is completely white like the adults. However, hatch-year birds of both color morphs lack the plumes and bicolored bill of adults (Lowther and Paul, 2002). Birds were identified as either foraging in groups or solitarily (referred to as foraging class). A group was ≥ 2 egrets foraging with < 30 m of spacing between individuals during most of the observation period (Bates and Ballard, 2014). Coordinates at the center of the foraging site for each sampled individual were determined with a GPS unit, and a transect was established along the path of the foraging egret. A 1-m quadrat was used to measure percent seagrass coverage and seagrass species composition at ≥ 5 points at 25-m intervals along the transect. Water depth was measured at each quadrat sampling point. We averaged across sampling points on each transect to determine average water depth and percent seagrass cover for each bird. t-tests were used to examine differences in water depth and percent seagrass cover between ages, color morphs, and foraging class (solitary or group) as foraging success or habitat use by reddish egrets have been shown to differ between ages (Bates and Ballard, 2014), color morphs (Green 2005) and foraging classes (Bates and Ballard 2014). Habitat data collected during observations was used to develop this study model and examine foraging habitat availability for subsequent time periods.
Model building
Optimal water depth for foraging reddish egrets was determined to be 1 to 20 cm based on ~ 90% of the sample foraging within this range. A raster dataset of bathymetry (60 x 60 m grid size) for the Laguna Madre (United States Army Corp of Engineers, unpublished data), Digital Elevation Model (3 x 3 m grid size; United States Geological Survey, 2009), and tide readings from four tide gauges in the Laguna Madre (National Oceanic and Atmospheric Administration Tides and Currents, 2014) were used in ArcGIS 10.1 ModelBuilder (Environmental Systems Research Institute, 2011) to ModelBuilder (Environmental Systems Research Institute, 2011) to determine weekly changes in water depth and extent of available foraging habitat during January 2012 to April 2014. The bathymetry dataset did not extend completely into the tidal flats so the Digital Elevation Map, created from Lidar Imagery (10 x 10 m grid size) was merged with the bathymetry dataset by creating a 300-m buffer on the landward border of the bathymetry polygon and a 300-m buffer on the border of the Digital Elevation dataset where it met the Laguna Madre. Next, 10 000 random points were generated in the bathymetry buffer polygon and 10 000 random points in the Digital Elevation dataset buffer polygon. The values of the bathymetry layer and the Digital Elevation layer to the random points were extracted, then the two random point datasets were merged into one layer and an Inverse Distance Weighted analysis was performed to interpolate the values between the two layers to provide a fluid merging process of the two raster datasets. The three datasets (bathymetry, digital elevation and the created dataset interpolated from random points) were mosaicked into one raster layer at a 3 x 3 m resolution. The combined dataset resulted in a gradual merging of values between the bathymetry data and the Digital Elevation Map data over the areas that were not covered by either dataset. Observations of foraging reddish egrets in the Laguna Madre indicated that forging was most intense from sunrise to 10:30 (Bates, 2011). Therefore, 08:30 (median time) was chosen as a reference time to interpolate the tide throughout the Laguna Madre. One weekday (Wednesday) was randomly chosen and tide gauge readings from that day at 08:30 were used each week for the 120 weeks from 4 January 2012 to 16 April 2014 to estimate weekly water depth throughout the Laguna Madre. Readings from the tide stations in the Laguna Madre were used along with the inverse distance weighted tool in ArcGIS 10.1 to interpolate the tide. The interpolated tide was then combined with the merged bathymetry–DEM dataset using the raster calculator tool to obtain water depth throughout the Laguna Madre. The resulting data layer was reclassified giving water depth between 1 and 20 cm, a value of one, and all other depths a value of zero.
The Coastal Bend Texas Benthic Habitat dataset for the Laguna Madre was also used to help delineate foraging habitat based on benthic characteristics (National Oceanic and Atmospheric Administration Ocean Service, Coastal Services Center, 2008). Benthic habitat types classified as suitable reddish egret foraging habitat were based on our results of seagrass coverage from sampled foraging locations, as well as habitat descriptions by Farmer (1991) and Lowther and Paul (2002). Suitable habitat types were given a value of one and included unconsolidated bottom, patchy seagrass, bivalve reef, land and unknown benthic. All other habitat types including continuous seagrass and emergent marsh were coded as zero, indicating that they were unsuitable foraging habitat. The recoded benthic habitat dataset was then multiplied by the reclassified water depth layer to create new layers containing values of zero (unavailable) and one (available) for each week across the 120-week period. The new layers were then converted to polygons to allow calculation of the area of suitable foraging habitat during each week.
Model assessment
To assess the accuracy of this study model, GPS locations of 8 adult reddish egrets marked with satellite transmitters during January 2012 – April 2014 were used. Transmitters were programmed to record locations at 08:00 and 09:00 each morning and 04:00 and 05:00 each evening, which are peak foraging hours (Bates, 2011). We used locations from every day at each of the 4 times (n = 6 385) to assess the model’s ability to delineate foraging habitat availability across the 120 weeks. The authors then used locations from only Wednesdays at 08:00 and 09:00 (n = 770) to
correspond with each day and assess performance of the model at a finer temporal scale. They considered locations ≤ 50 m from available habitat as ‘in’ available habitat to account for GPS accuracy (±18 m) and inherent, unknown error in extrapolation of bathymetry and tide data. ArcGIS 10.1 was used to measure the distance from foraging locations to available foraging habitat for each week.
RESULTS
Foraging habitat of 372 reddish egrets that were observed foraging throughout the Laguna Madre were sampled; 211 were observed foraging solitarily, and 161 were observed foraging in groups (n = 21). Average water depth used by foraging reddish egrets was 10.1 ± 0.68 cm with no differences (P > 0.05) between ages, color morphs or foraging classes (Table 1). Approximately, 91% of reddish egrets foraged in water depths of 1 to 20 cm. Average seagrass coverage along transects was 12.3% (± 2.74), and 67% of egrets foraged in ≤ 10% seagrass coverage. There were no differences (P > 0.05) in percent seagrass coverage between ages, color morphs or foraging classes (Table 1).
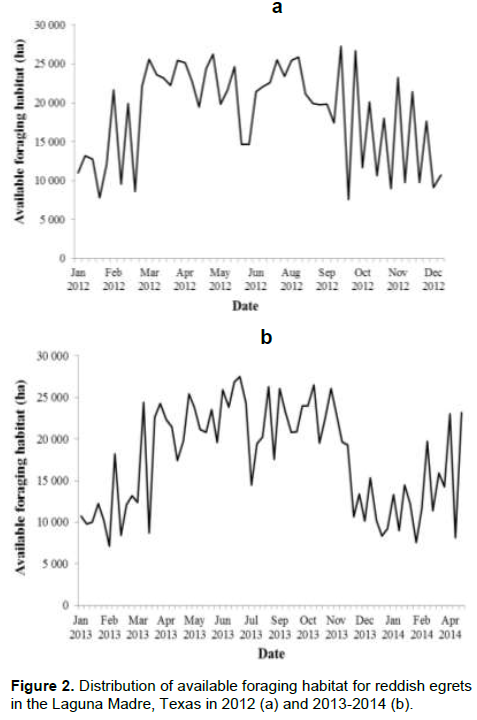
The Laguna Madre encompasses 215 223 ha of which 123 395 ha were considered potential foraging habitat based solely on benthic habitat characterization (prior to adding water depth to the model). Once water depth was added to the model, available foraging habitat was reduced considerably and varied throughout the annual cycle with a total of 74 985 ha of foraging habitat available at some point during the 120-week period. In 2012, the amount of foraging habitat was relatively low and highly variable from January to mid-March; availability became much higher and remained relatively stable from mid-March through late September, then was again relatively low, but highly variable from October to December (Figure 2a). The lowest amount of available foraging habitat occurred in March (8 608 ha) and peaked at 27 247 ha in late September. In 2013, the overall pattern was similar with relatively large amounts of foraging habitat during April–October, and lower amounts with greater variability during winter (Figure 2b). Foraging habitat peaked at 27 534 ha in July and reduced to 7 099 ha in February. On average, available foraging habitat throughout winter was about 50% of that available during summer.
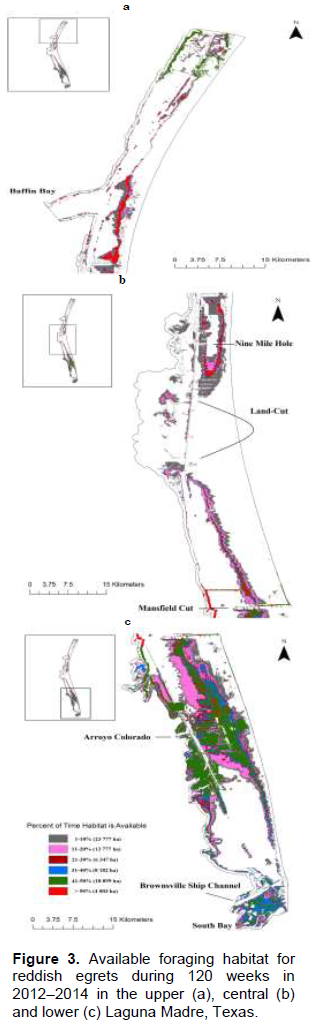
The availability of a given foraging area fluctuates; the longest duration specific areas available was 82 weeks (2.52 ha). Only 4 003 ha were available ≥ 50% of the time as compared to 51 208 ha available ≥ 10% of the time. Areas containing the greatest amount of foraging habitat in the upper Laguna Madre (Figure 3a and b) were located east of the mouth of Baffin Bay and south to Nine Mile Hole. Large areas of foraging habitat in the lower Laguna Madre were located on the east side, south of the Mansfield Cut, north and south of the Arroyo Colorado on the east side, and south of the Brownsville Ship Channel in South Bay (Figure 3b and c).
It was found that 90% of the GPS locations of foraging reddish egrets fell within our modeled available foraging habitat during the cumulative 120 week period, providing compelling evidence that our model performed well in delineating cumulative foraging habitat across the annual cycle. Less agreement (51%) was found between GPS locations of foraging reddish egrets as compared to corresponding estimates of foraging habitat during each week of the 120 week period. Locations falling outside the estimated foraging habitat deviated with an average of 407 m (median = 180 m) from the foraging habitat boundary and generally were shoreward. About 9% of locations were greater than 500 m from available habitat and in general were on the western part of the Laguna Madre towards the mainland and west of the land-cut. These areas had a much more patchy distribution of foraging habitat relative to other areas.
DISCUSSION
This study’s foraging habitat model appeared to perform well in estimating the distribution of foraging habitat for reddish egrets in the Laguna Madre, as there was 90% overall agreement with an independent dataset of 6 385 foraging locations from individuals marked with satellite transmitters. The weekly locations (n = 770) were less accurate, with half occurring within available habitat; yet the average distance the locations were from available habitat was relatively small. Potential causes of the reduction in agreement between GPS locations of foraging reddish egrets and the modeled foraging habitat include birds not actually foraging during the time the location was recorded (e.g., flying between foraging locations), inherent error in geospatial datasets and wind-driven water level changes in shallow flats not being accurately recorded by tide gauge stations. However, the influences of these potential sources of variability on modeled foraging habitat that became apparent at finer time scales appear to be less evident when modeled over longer time periods. This model showed that the availability of foraging habitat in the Laguna Madre varied both temporally and spatially. Spatial and temporal fluctuations of foraging habitat in wetland ecosystems is not uncommon; Arengo and Baldassarre (2002) found that size, quality and duration of available foraging habitat for American flamingos (Phoenicopterus ruber ruber) varied considerably within and among foraging sites in Yucatán, Mexico. Spatial and temporal variability of wading bird foraging habitat is common due to tidal cycles and variable precipitation (Kahl, 1964; Kushlan 1981; Gonzalez, 1997). Tides in the Laguna Madre are primarily wind driven (Tunnell, 2002); thus, any change in wind speed and direction can cause fluctuations in foraging habitat. The amount of available foraging habitat for reddish egrets was much greater during spring and summer, and large peaks of available habitat occurred within this time period. This period coincides with the breeding season, when energy requirements are high relative to other periods of the annual cycle (Lowther and Paul, 2002).
The amount and location of foraging habitat during the breeding season are important factors to consider when managing this species. The location and size of breeding colonies are often directly related to the availability of nearby foraging habitat (Gibbs et al., 1987; Gibbs, 1997). Green Island in the lower Laguna Madre typically contains the largest abundance of breeding reddish egrets, as well as all other herons and egrets, along the Texas Coast (Texas Colonial Waterbird Society, unpublished data). This island is located proximal to large areas of foraging habitat identified by our model and that are known to be used by reddish egrets nesting on Green Island (Farmer, 1991). Protection of this foraging habitat is likely an important factor in the maintenance of the colony. Likewise, spatially linking foraging habitat to other colonies throughout the Laguna Madre could be beneficial in understanding resources that are critical for sustaining breeding colonies and help reduce adverse impacts from anthropogenic influences (e.g., dredge disposal). More information is needed to examine how reddish egrets utilize foraging habitat in relation to breeding colonies and the distances reddish egrets can travel to foraging sites without reproduction being affected.
The timing of reproduction results in both post-breeding adults and young of the year having large amounts of stable foraging habitat during early fall. Bates and Ballard (2014) found groups of foraging adult and juvenile reddish egrets just after the breeding season from August to October. Farmer (1991) observed a similar pattern of group foraging by hatch-year reddish egrets in fall. Hatch-year reddish egrets are often less efficient foragers and may not be as skilled at identifying quality foraging areas (Bates and Ballard, 2014). The abundance and spatial stability of foraging habitat in fall probably results in hatch-year egrets spending less time and energy searching for quality foraging sites than if there was high temporal variation in availability, and allows them to spend more time foraging to meet their energy demands.
The decline in availability of foraging habitat from December to March corresponds with the time when reddish egrets depart the Laguna Madre during winter and arrive during spring (Farmer, 1991). There is a partial migration of reddish egrets in late autumn, but how far south they migrate is unclear (Paul, 1991). Limited banding data suggest reddish egrets migrate into Mexico and even El Salvador and Guatemala in search of foraging habitat (Paul, 1991). Recently, adult reddish egrets equipped with satellite transmitters were shown to migrate into Mexico during the end of November and beginning of December, returning to the Laguna Madre in March-April (Ballard, unpublished data). Partial migration often occurs when resources become limited and are unable to support the entire population (Ketterson and Nolan, 1976; Lundberg, 1988). The timing of departure from the Laguna Madre during early November by migrant reddish egrets coincides with the decrease in available foraging habitat, indicating that the amount of winter foraging habitat in the Laguna Madre may not be sufficient to maintain the large concentration of reddish egrets in this system.
An initial consideration when prioritizing foraging area for protection should be the temporal extent of its availability. Our habitat model indicated that no foraging habitat pixels (from the final raster image) were available for the entire two-year period and only a small fraction was available more than half of the time. The highly dynamic nature of foraging habitat availability in the Laguna Madre is primarily the result of wind-driven tides (Tunnel, 2002). Because of this high degree of spatial variation in where foraging habitat is located at any time during the annual cycle, it would be required to consider large areas of foraging habitat when devising conservation plans to ensure that enough is available at any given time to sustain the reddish egret population.
Foraging habitat assessment at both smaller spatial and temporal scales within the Laguna Madre may be beneficial in understanding how these habitat dynamics influence reddish egrets. For example, foraging habitat availability during the nestling period influences the provisioning rate by adults, thereby indirectly impacting fledging success. This was the case in the Florida Everglades where nest success of great egrets and white ibises (Eudocimus albus) is lower in years with below average prey availability as compared to years with high prey availability (Herring et al., 2010). Prey availability was not examined in this study; however, sheepshead minnow has been identified as a major prey item of reddish egrets breeding in Texas (Holderby et al., 2014) and the habitat of sheepshead minnow largely coincides with the foraging habitat of reddish egrets, that is, shallow water with minimal vegetation. Therefore, foraging habitat availability is likely related to prey availability.
The authors were able to delineate foraging habitat over a large spatiotemporal scale and to identify important foraging areas for protection and conservation, which is a current goal of the Reddish Egret Conservation
Action Plan (Wilson et al., 2014). This model may also be used in future analyses to examine shifts in foraging habitat and the potential impact of sea level rise on foraging habitat availability. Further, it can be used quite effectively to plan dredging operations in the Laguna Madre, by identifying areas where deposition of dredge material will have minimal impacts on foraging habitat for reddish egrets and other waterbirds.
CONFLICT OF INTEREST
The authors have not declared any conflict of interest.
ACKNOWLEDGEMENTS
This project was funded by the U.S. Fish and Wildlife Service Regions 2 and 4 Migratory Bird Programs, the Walter Fondren III Fellowship in Shorebird and Wading Bird Research at the Caesar Kleberg Wildlife Research Institute, the Rachel and Ben Vaughan Foundation, and the Fort Worth Zoo. The authors thank E. Redeker for providing input for the model development and three anonymous reviewers for their constructive comments. This is manuscript # 16-133 of the Caesar Kleberg Wildlife Research Institute.
REFERENCES
|
Arengo F, Baldassarre GA (2002). Patch choice and foraging behavior of nonbreeding American Flamingos in Yucatán, Mexico. Condor 104:452-457. |
|
|
Bates EM (2011). Foraging ecology of reddish egrets in the Laguna Madre of Texas. M.S. Thesis, Texas A&M University-Kingsville. |
|
|
Bates EM, Ballard BM (2014). Factors influencing behavior and success of foraging reddish egrets (Egretta rufescens). Waterbirds 37:191-202. |
|
|
Cezilly F, Boy V (1988). Age related differences in foraging Little egrets, Egretta garzetta. Colon. Waterbirds 11:100-106. |
|
|
Environmental Systems Research Institute (ESRI) (2011). ArcGIS v. 10.3. ESRI, Redlands, California. |
|
|
Farmer M (1991). Reddish Egrets of the lower Laguna Madre, Texas. United States Fish and Wildlife Service, Alamo, Texas. P 54. |
|
|
Gibbs JP (1997). Determinants of the size and location of Great Blue Heron colonies. Colon. Waterbirds 20:1-7. |
|
|
Gibbs JP, Woodward S, Hunter ML, Hutchinson AE (1987). Determinants of Great Blue Heron colony distribution in coastal Maine. Auk 104:38-47. |
|
|
Gonzalez JA (1997). Seasonal variation in the foraging ecology of the Wood Stork in the southern Llanos of Venezuela. Condor 99:671-680. |
|
|
Green MC (2005). Plumage dimorphism in the reddish egret: does plumage coloration influence foraging habitat use and tactics? Waterbirds 28:519-524. |
|
|
Herring G, Gawlik DE, Cook MI, Beerens JM (2010). Sensitivity of nesting great egrets (Ardea alba) and white ibises (Eudocimus albus) to reduced prey availability. Auk 127:660-670. |
|
|
Holderby Z, Hill A, Palacios E, Green MC, Amador E, de Dios C (2014). Comparisons of reddish egret (Egretta rufescens) diet during the breeding season across its geographic range. Waterbirds 37:136-143. |
|
|
International Union for Conservation of Nature (IUCN) (2012). The IUCN red list of threatened species, v. 2014.3. BirdLife International, Gland, Switzerland. |
|
|
Kahl Jr. MP (1964). Food ecology of the Wood Stork (Mycteria americana) in Florida. Ecol. Monogr. 34:98-117. |
|
|
Ketterson ED, Nolan Jr. V (1976). Geographic variation and its climatic correlates in sex-ratio of eastern wintering Dark-eyed Juncos (Junco hyemalishyemalis). Ecology 57:679-693. |
|
|
Kushlan JA (1981). Resource use strategies of wading birds. Wilson Bull. 93:145-163. |
|
|
Lowther PE, Paul RT (2002). reddish egret (Egretta rufescens), The Birds of North America Online (A. Poole, Ed.). Ithaca: Cornell Lab of Ornithology |
|
|
Lundberg P (1988). The evolution of partial migration in birds. Trends Ecol. Evol. 3:172-175. |
|
|
Maheswaran G, Rahmani AR (2002). Foraging behaviour and feeding success of the Black-necked Stork (Ephippiorhynchus asiaticus) in Dudhwa National Park, Uttar Pradesh. India J. Zool. 258(2):189-195. |
|
|
Moulton DW, Dahl TE, Dall DM (1997). Texas coastal wetlands: status and trends, mid-1950s to early 1990s. U.S. Department of the Interior, Fish and Wildlife Service, Southwestern Region, Albuqerque, New Mexico. P 32. |
|
|
Paul RT (1991). Status report: Egretta rufescens (Gmelin) – Reddish Egret. U.S. Fish and Wildlife Service, Region II, Albuquerque, New Mexico and U. S. Fish and Wildlife Service, Office of Ecological Services, Houston, Texas. |
|
|
Quammen ML, Onuf CP (1993). Laguna Madre: seagrass changes continue decades after salinity reduction. Estuaries 16:302-310. |
|
|
Tunnell Jr. JW (2002). Geography, Climate, and Hydrography. Pages 7-27 in The Laguna Madre of Texas and Tamaulipas. (JW Tunnell Jr. and FW Judd, Eds.). Texas A&M University Press, College Station, Texas. |
|
|
Wilson TE, Wheeler J, Green MC, Palacios E (2014). Reddish Egret Conservation Action Plan. Reddish Egret Conservation Planning Workshop, October 2012. Corpus Christi, TX. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


