There are a wide suite of conservation issues in the modern age (Gurevitch and Padilla, 2004; Giam et al., 2010; Bellard et al., 2012), each having a variety of often negative implications on global biodiversity, and each demanding attention (Sheil, 2001), research, and funding to address (or at the very least, to attempt to do so). One of the biggest conservation concerns, and one that is very often implicated in extinctions of species in modern times, is habitat loss (Simberloff, 1984; Tilman et al., 1994). Habitat loss causes a reduction in population size, leaving a species more prone to the effects of stochastic events (Burkey, 1995), and thus increasing the potentiality of its extinction. The loss of habitat in general tends to outweigh the effects of fragmentation of such habitat (Fahrig, 1997), and is aggravated by land conversion tending to be non-random (Seabloom et al., 2002), biased towards areas valued agriculturally or those with a particular geographic placement.
Panthera pardus is classified as a vulnerable predatory species according to the latest IUCN Red List (Stein et al., 2016). It has a wide distribution comprising a broad region in Africa (with the Sahara Desert excluded from its range), the Arabian Peninsula, southwest Asia, as well as a small population maintained in the Russian Far-East (Nowell and Jackson, 1996). While some estimates show its Southern African range as being in no immediate danger of a severe decline (Martin and de Meulenaer, 1988), there have been criticisms of these estimates (Norton, 1990), as there are marked reductions in areas of encroaching human settlement and other habitat conversions, with range decline for the species being as high as 37% in a period of 100 years (Ray et al., 2005).
P. p. pardusis an African sub-species of the leopard, with some 78% of the overall species range occupied by this variant (Jacobson et al., 2016). South Africa is responsible for some of the biggest habitat losses for the species, with leopards in unprotected areas being severely restricted in their occurrence (Skead, 2007). One case of this was noted in the Phinda-Mkhuze Complex, a small region lying along the eastern edge of South Africa, where the observed number of leopards was on average 11.11/100 km2 within the protected Mkhuze Game Reserve (core protected area), 7.17/100 km2 in the neighbouring Phinda Private Game Reserve (buffer protected area), and then only 2.49/100 km2 in the surrounding non-protected lands used for livestock farming, private game ranches and tribal land (Balme et al., 2010). With poaching and trophy hunting still being prominent factors driving leopard population in South Africa, there is an added potential risk of leopards being subjected to retaliatory farmer killings because of the real and perceived threats that these animals pose to livestock, as leopards tend to easily cross boundary fences (Balme et al., 2009; Chapman and Balme, 2010). Therefore, despite about 20% of South Africa currently providing suitable leopard habitat (Swanepoel et al., 2013), there is a need for a thorough investigation of the potential of the species to experience a further decline, as suggested by the decreasing population trend predicted by the IUCN (Stein et al., 2016).
An important factor to consider when determining the potentiality of a population decline is to examine the species’ physiological needs against the resources that the environment in which the species occurs is actually capable of providing at any given time (Wikelski and Cooke, 2006). If the surrounding environment falls short of meeting an animal’s physiological needs, the animal is faced with a decision to either relocate elsewhere in search of resources or to face death from starvation in its current habitat (Stephens, 2008). However, where threats such as poaching and legal hunting outside the habitat pose a life-threatening risk of their own (as earlier mentioned) – the surrounding matrix is in itself an unpredictable habitat – the animal may still face the same fate outside the poor habitat as it would by remaining there (Switzer, 1993). Therefore, using the physiological needs of species to determine the required energetic quality of the habitat to maintain a viable population is a useful technique in helping inform habitat management (particularly where such habitat is facing the potentiality of, for example, habitat loss (Fahrig, 2001)).
When assessing the energetic needs of a predator, it is of fundamental importance to consider the interactions that they have with their potential prey – a relationship that is instrumental in regulating and shaping both populations and communities at large (Fretwell, 1987). Not only does the predator-prey relationship affect prey numbers as per predator kills, but extensive phenotypic changes can be induced in the prey as a response (Werner and Peacor, 2003), and prey intimidation has an effect on their demographics comparable to those resulting from prey consumption (Preisser et al., 2005). In accounting for the predator-prey interaction that exists between leopards and their prey (mostly mammalian species with a weight range of 10 to 40 kg, as per Hayward et al., 2006), it is also important to consider the effects of spatial heterogeneity on modifying the functional response (Gorini et al., 2012). In a heterogeneous system that tends to persist in real-life scenarios, the leopard will face regular challenges not only in searching for and encountering prey, but also in actually killing and consuming it.
A factor complicating the matter even further in the modern times is human disruption: anthropogenic activities can directly influence the nature of the relationship between a predator and its prey, whether by controlling the numbers of the respective species, or by providing food subsidies (Rodewald et al., 2011). In the case of the leopard, subsidies may be provided accidentally, such as domestic livestock that the predator may take to hunting (Kissui, 2008). Being assisted by the natural tendency of leopards to roam widely in search of prey, the animals can easily become involved in tense human-animal interactions with the owners of private lands (as earlier mentioned). As such, in assisting the survival of the species, conservation efforts need to be adequately backed by knowledge of how much wild prey should be stocked in reserves, to allow leopard densities to stabilize in accordance with their main natural prey, as is common for predators (Karanth et al., 2004). This should also assist in reducing incidences of livestock killing. To determine this, a detailed analysis is needed of the difference between energetic consumption and expenditure of the animal, and how vulnerable it actually is to having its energy intake fall below its rate of utilization, upon which a steady decline in weight and activity levels ensues and the potential of death becomes a serious threat. This parameter is closely examined henceforth.
An energetic model for P. pardus was constructed, using standard work (W) equations (presented in kilojoule (kJ) measurement units), which relates directly to kinetic energy according to the work-energy principle. Input values for the model were sourced from a variety of literature, obtaining specific values on feeding, hunting and other behaviours as is accurate for the species. For the purposes of standardization, the p. pardus subspecies was chosen as the subject to allow for maximal consistency of the input values, given the well-defined genetic differences between the various subspecies (Miththapala et al., 1996).
Where required energetic values were not available specifically for the African leopard, the closest approximation was chosen, that is, first looking for values concerning the leopard, followed by a search for felids at large, etc. The model environment selected for the subject to operate in was the savanna biome of Southern Africa, more specifically the northern Kwa-Zulu Natal (due to the concentration of leopard studies in that region), obtaining the physiologically relevant seasonal temperatures, the diurnal/nocturnal differences as appropriate for the region, as well as other atmospheric properties such as average wind speed, from literature sources.
Following this, a statistically-average African leopard was defined, to allow for the refinement of the energetic results obtained. An average daily energetic gain was determined for the leopard subject, using the values obtained for prey consumption. On the opposite end of the scales, the basal metabolic energy consumption of the subject was determined, followed by heat-related costs to the organism (such as due to radiation, convection, etc.). Lastly, energy costs associated with obtaining prey were calculated. Summing all the determined energetic costs and weighing those up against the average energetic gains the leopard can be expected to obtain, allowed the determination of the extent of the difference between the energy gains and losses. The energetic surplus would be used by the organism to meet other, non-daily, needs such as breeding effort or emigration out of an area (Parker et al., 2009).
The results of the model provide another important finding: an estimation of how many hunting attempts on average before success are possible to warrant an organism’s continued survival. Using this value, various scenarios of catch probability were tested to determine how many hunting attempts were required before a catch was made under each case. Given that a higher prey density is strongly related to the incidence of a successful catch (Whitfield, 2003), catch probability served as a proxy for prey stocking rates in the environment tested. This allowed for the determination of the lowest hunting success probability (that is, prey stocking rates) at which the leopard would still be able to obtain a hunt before the average number of hunting attempts, as allowed by standard daily energy reserves, were exhausted.
One of the main factors involved in energy expenditure, is that required for the correct functioning of internal organs such as the heart, the lungs, liver, etc. Even while being within a state of relative physical inactivity, there is a continuous utilization of energy for the continuation of basal metabolism, characterized by catabolism of compounds with oxygen intake. The rate of oxygen consumption by the animal, the energetic output produced, as well as the rate of carbon dioxide release, is interrelated. If carbohydrates are subjected to oxidation, the intake of one litre of oxygen frees up 21.13 kJ of energy. If there is a similar oxidation of proteins and lipids, then 19.66 and 20.08 kJ of energy are released, respectively. For an adult animal on average, the hourly basal metabolism uses up 4.2 kJ for a kilogram of body weight.
Energetic balance analysis defines the following parameters for an adult leopard of mean statistical parameters in Southern African savanna: a body mass of 60 kg and a body surface area of 1.53 m2 (using Meeh coefficient of 10 for an average cat, given by Schmidt-Nielsen, 1984). P. pardus has an internal body temperature of about 39.86°C (Deka et al., 2012), while the ambient temperature in its savanna habitat fluctuates seasonally (Balme et al., 2007). Midday temperatures range from 23°C in July to 30°C in January (BirdLife South Africa, 2016), while night-time temperatures of 11°C in July and 20°C in January. Averaging, we obtain: 26.5°C in summer and 15.5°C in winter. The dermal layers of an animal tend to be a few degrees cooler than the rectal temperatures as recorded by Deka et al. (2012), with the epidermis being where internal and ambient temperatures meet (ambient temperatures averaging about 26°C in Southern African savanna). Given the aforementioned considerations, this study assumes an average epidermal temperature to be about 30°C for the African leopard (p. pardus).
Further, the study assumes that a statistically-average African leopard in African savanna has a daily meat consumption of 3.25 kg, since this is the median value of a leopard’s 1.6-4.9 kg meat/day consumption recorded in the literature (Bothma and Le Riche, 1986; Bailey, 1993; Stander et al., 1997). African leopards are known to strongly prefer killing impala and bushbuck, with an average body mass of 23 kg (Hayward et al., 2006). The energetic content of this game can be estimated at 8.5 kJ/g, since venison is considered to be a much leaner type of meat than that of cattle, with a common fat content being less than 3% (Schönfeldt, 1993; Hoffman, 2000). The quantity of energy obtained by an African leopard in a single statistically-average 24-h period therefore becomes:
WEO = 8.5kJ*(1000*3.25kg) = 27625 kJ.
Utilization of the energy source provides for sustenance of a leopard’s life until the next successful hunt. Loss of energy due to basal metabolism can be presented as:
WBM = 4.2 kJ*24 h*60 kg = 6048 kJ
Alongside the basal metabolism energetic expenditure, there are significant costs associated with heat exchange of the body with the exterior environment. In the complex process of maintenance of a heat balance, a major importance is the intricate regulation of heat loss (Berkovich, 1964). In physiology, bodily heat transfer can be viewed as the loss of heat, freed through activities associated with living, into a cooler environment (Ivanov, 1990). There are four key modes of heat transfer between an animal and its environment: radiation, convection, conduction and evaporation; the latter being dominant in case of overheating. However, when existing under conditions of a comfort temperature zone, the greatest exchange is provided by radiation and convection (Fanger, 1970). The total energy radiated from a unit of bodily surface area is directly proportional to the fourth power of the thermodynamic absolute temperature, as detailed by the Stefan-Boltzmann law. When there is but a small difference between the animal’s epidermal temperature and the ambient temperature (as is generally the case in the Southern African part of a leopard’s distribution), the equation for radiative heat loss can be presented in the following format:
WR = hrad*S*(δ1 - δ2)*t,
where hrad is the radiative heat transfer coefficient of WR (in m2/ °C), S is the leopard’s body surface area (in m2), δ1 is the leopard’s epidermal temperature, δ2 is the ambient temperature, and lastly, t is length of time over which radiation is being measured (in seconds).
By the Stefan-Boltzmann law, the radiative heat transfer coefficient hrad between two gray surfaces can be determined with the equation:
hrad = εσ(T12+T22)(T1+T2),
where ε is the emissivity of the leopard’s epidermis, σ is the Stefan-Boltzmann constant (σ = 5.67*10-8 W/m2.K4), T1 is the epidermal temperature of the leopard and T2 is the ambient temperature (both temperatures being absolute).
Emissivity varies with the radiation wavelength, but is close to unity at wavelengths greater than 5 μm (Ingram and Mount, 1975), therefore a value of 0.9 is here assumed for ε. T1 is 30°C and T2 is 26.5°C for daytime, with T2 becoming 15.5°C average at night (as explained previously). From the abovementioned, the calculation for hrad becomes:
(0.9) (5.67*10-8) (3032+299.52) (303+299.5) = 5.58 during the day, and (0.9)(5.67*10-8)(3032+288.52) (303+288.5) = 5.28 at night.
Therefore for WR, we now have:
WR = 5.58*1.53*(30-26.5)*3600 ss*12 h = 29.88 J/s *3600 seconds*12 h = 1290.85 kJ for 12 h of daytime; and WR = 5.28*1.53*(30-15.5)*3600 s*12 h = 117.14 J/s*3600 s*12 h = 5060.31 kJ for 12 h of nighttime. This assumes the yearly average of equal day and night length, and costs 1290.85+5060.31 = 6351.16 kJ for a statistically-average 24 h period.
Transfer of convection heat occurs between body surface and air temperature and its motion (Ingram and Mount, 1975). In calculation, the biggest difficulty takes place with determination of the size of hC (convective heat transfer coefficient), due to this value fluctuating greatly as a result of its dependency on factors such as air temperature, the shape of body form, its size, etc. The most profound influence on hC is had by wind speed, which sets the strength of the forced convective heat transfer. In the northern Kwa-Zulu Natal, wind speed averages at about 5 ms-1 (Weather, 2017). At this wind speed, the value of hC is about 690% greater than at the common indoor wind speed of 0.2 m.s-1 (Mitchell, 1974). In an animal, convection occurs within the layer of exposed fur, which provides some insulation against the wind. However, at wind speed of 5m.s-1, about half of this insulation will be lost (McArthur, 1981). Since no forced convective heat transfer coefficient has ever been determined for a wild felid species, the following is an approximation (determined for a sheep by Joyce et al. (1966)):
hC = 7.1v0.5,
where v is the wind speed (in m.s-1).
Substituting the wind speed of 0.2 m.s-1, we obtain a value of 3.18 for hC. In northern Kwa-Zulu Natal, considering the average wind speed frequent in the region, the value of hC will be 690% greater: 3.18*6.9 = 21.91. The precise quantity of convectional heat transfer can then be determined by the equation of Newton-Richman:
WC = hC*S*(δ1 - δ2)*t = 21.91*1.53*(30 - 26.5)*(3600 seconds*6 hours) = 2534.29 kJ for 6 h of daytime activity.
For nighttime, WC becomes:
21.91*1.53*(30-15.5)*(3600 s*6 h) = 10499.18 kJ.
If we assume that the African leopard is a predominantly nighttime hunter, performing most stalking and chasingactivities in the cooler temperatures, the animal will expend (2534.29*0.2)+(10499.18*0.8) = 8906.20 kJ. From existing literature, heat loss due to radiation and convection forms 73 to 88% of overall energetic heat losses (Ivanov, 1990); therefore it is best to assess the remaining heat-related energetic costs through relational means:
WE = [(WR + WC)/75]*25 = [(6351.16+8906.20)/75]*25 = 5085.79 kJ
From the aforementioned, the summative energetic costs attributed to heat transfer in general become:
Wheat = WR + WC + WE = 6351.16 + 8906.20 + 5085.79 = 20343.15 kJ.
Having accounted for both basal metabolism and heat transfer, it is important to consider the energy expenditure an African leopard is likely to incur while obtaining food. A leopard’s hunt consists of a number of stages (Stander et al., 1997): regular average-speed runs (10 km/h) to detect the presence of potential prey, a period of stalking and crouching, followed by a brief sprinting phase, during which speeds of up to 60km/h (Nowak, 1999) are reached.
In case of the attempt being unsuccessful, the hunting process repeats itself all over. On average, every 1 in 4/5 hunts are successful (Bailey, 1993; Stander et al., 1997). When a prey item is killed, the leopard is likely to attempt relocating it to a competitor-free zone, dragging it at speeds of about 6 km/h (equating to about 1.7 m/s). Although there are known cases of p. pardus feeding on the same carcass for a few days, they often lose their kill after the initial feeding, frequently to hyenas (Creel et al., 2001).
Utilizing the aforementioned information, we can formulate a model of a standard African leopard hunt. The summative 10 km/h runs constitute a distance of about 8 km of daily movement. During this time, it initiates a maximum of 5 sprints as part of its hunting attempts, which together cover a distance of 200 m at 60 km/h speeds, an average of 40 m covered per sprint (Bothma, 1998).
In this model, energy losses associated with conversion from average to maximum speeds and back are not considered. The leopard’s ideal prey of 23 kg weight is dragged at speeds of 6 km/h for an average of 320 m (Smith, 1978). Therefore, average daily runs:
W1 = mv12/2*t1 = 60kg*(2.82m/s)/2*2880 seconds = 677.38 kJ
The maximum 5 hunting sprints per day (final one being successful):
W2 = mv22/2*t2 = 60 kg*(16.62m/s)/2*12.05 s = 99.61 kJ
Relocation of the prey carcass to a safe feeding location:
W3 = (m1+m2)*v32/2*t3 = (60+23)*(1.72/2)*188 s = 225.48 kJ
Summative daily expenditure of kinetic energy on movement activities:
Wkinetic = W1 + W2 + W3 = 677.38 + 99.61 + 225.48 = 1002.47 kJ
Summing the leopard’s overall daily energetic costs, we obtain:
W = Wheat + Wkinetic + WBM = 20343.15 + 1002.47 + 6048 = 27393.62 kJ
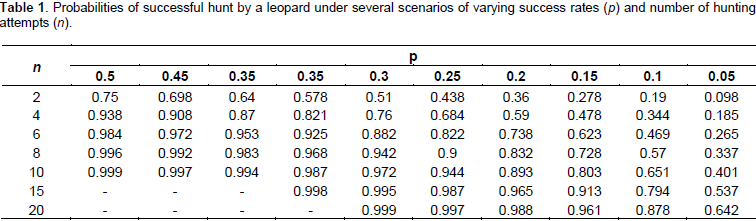
From the aforementioned, there is very little difference between the daily energy obtained (27625 kJ) and that used up on essential survival activities (27393.62 kJ). The small surplus of energy can either be used on particular activities which do not form part of daily routine (such as mating, territory defense, or unusually lengthy movement due associated with relocation), or can be retained for use the following day. Therefore for a statistically average African leopard, a sustained energetic balance is possible if, and only if, the biomass of prey is maintained at a level high enough to allow for successful food acquisition after at most the 5th hunting attempt. Let us consider the probability of such a situation. Marking every successful hunting sprint of a leopard with the variable p, the probability of a successful hunt after n attempts can be presented as:
R1,n = 1 – qn,
where q = 1 - p, and is the chance of failure.
Using this equation, the values of p can be tested from 0.5 to 0.05, alongside varying n values, the results of which are presented in Table 1. As can be deduced from Table 1, under a success rate probability of p = 0.5, practically four or five hunting attempts are required before a kill is almost guaranteed to be made (chances of a kill being above 0.9). Under conditions of p = 0.3, the number of attempts required for the same effect almost doubles; while at p = 0.1 there need to be at least 20 hunts to allow for at least one success, a hunting requirement that cannot be adequately met on average, if using the energetic balance determined earlier.