Full Length Research Paper

ABSTRACT
The key role of ectomycorrhizal (EcM) fungi in ecosystems functioning has been demonstrated worldwide. However, their diversity, spatial distribution, fruiting phenology and production as influenced by climatic parameters variability remain poorly understood in tropical African forests. Weekly surveys were conducted from April to early October 2014 at the Comoé National Park (CNP), Côte d’Ivoire (West Africa) in 09 permanent plots established in Isoberlinia doka (IW), Uapaca togoensis (UW) and Mixed (MW) woodlands. Non metric multidimensional scaling (NMDS) of EcM fungi abundance was run to assess the influence of environment tal parameters on fungi distribution using the package VEGAN. Hierarchical clustering based on dissimilarity and indicator species analysis were run to characterize fungi communities. Analyses were computed with the statistical program R. A total of 123 EcM fungi species belonging to 23 genera and 09 families were collected at CNP. Simpson diversity (1-D) and evenness were 0.97 and 0.54, 0.97 and 0.61, 0.96 and 0.52 for IW, MW and UW respectively. Yet, weekly-based species accumulation curves did not reach an asymptote. Stem density of U. togoensis Pax (UTDen) and I. doka Craib & Stapf were the most important tree parameters influencing EcM fungi distribution (respectively r2 = 0.92 / p-value = 0.002 and r2 = 0.83 / p-value = 0.018). Two sites groups were distinguished and four indicators species were identified.
Key words: EcM fungi, fruit bodies, diversity, indicator species.
INTRODUCTION
MATERIALS AND METHODS


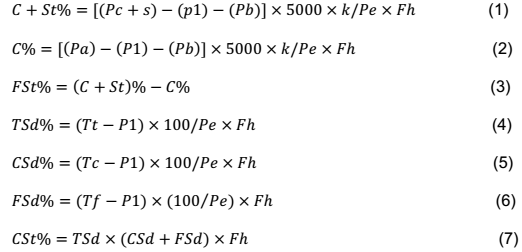

RESULTS




DISCUSSION
CONCLUSION
CONFLICT OF INTERESTS
ACKNOWLEDGEMENTS
SUPPLEMENTARY DATA




REFERENCES
|
AFNOR (1995). NF ISO 10694 (X31-409). Soil quality- Determination of organic and total carbon after dry combustion (elementary analysis). French norm AFNOR. |
|
|
AFNOR (1999). NFX 31-130 soil quality. Chemical methods, Determination of Cation Exchange Capacity (CEC) and Cation Extraction, AFNOR, Paris. |
|
|
AFNOR (2003). NFX 31-107 standard. Qualité du sol. Détermination de la distribution granulométrique des particules du sol. Méthode à la pipette. AFNOR, Paris. |
|
|
Andrew C, Lilleskov EA (2009). Productivity and community structure of Arbonnier M (2004). Trees, shrubs and lianas of West African dry zones. Margraf Publishers. |
|
|
Bâ AM, Duponnois R, Moyersoen B, Diédhiou AG, (2012). Ectomycorrhizal symbiosis of tropical African trees. Mycorrhiza. Springer-Verlag. |
|
|
Bahram M, Põlme S, Kõljalg U, Zarre S, Tedersoo L (2012). Regional and local patterns of ectomycorrhizal fungal diversity and community structure along an altitudinal gradient in the Hyrcanian forests of northern Iran. New Phytol. 193(2):465-473. |
|
|
Beeli M (1935). Amanita, Volvaria. Flore Iconographique des Champignons du Congo, 1:1-28. |
|
|
Boa ER (2004). Wild edible fungi: a global overview of their use and importance to people. Food & Agriulture Organization. |
|
|
Boddy L, Büntgen U, Egli S, Gange AC, Heegaard E, Kirk PM, Mohammad A, Kauserud H (2014). Climate variation effects on fungal fruiting. Fungal Ecol. 10:20-33. |
|
|
Braun-Blanquet J (1932). Plant sociology. The study of plant communities. The study of plant communities. 1st Ed. |
|
|
Büntgen U, Kauserud H, Egli S (2012). Linking climate variability to mushroom productivity and phenology. Front. Ecol. Environ. 10(1):14-19. |
|
|
Büntgen U, Peter M, Kauserud H, Egli S (2013). Unraveling environmental drivers of a recent increase in Swiss fungi fruiting. Glob. Change Biol. 19(9):2785-2794. |
|
|
Buyck B (1993). Russula I (Russulaceae). Flore illustrée des Champignons d'Afrique Centrale, 15:337-407. |
|
|
Buyck B (1994). Russula II (Russulaceae). Flore Illustrée des Champignons d'Afrique Centrale, 16:411-539. |
|
|
Buyck B (1997). Russula III (Russulaceae). Flore Illustrée des Champignons d'Afrique Centrale, 17:545-598. |
|
|
Buyck B, Thoen D, Watling R (1996). Ectomycorrhizal fungi of the Guinea-Congo region. Proc. R. Soc. Edinb. Section B. Bio. Sci. 104:313-333. |
|
|
Cao Y, Williams DD, Larsen DP (2002). Comparison of ecological communities: the problem of sample representativeness. Ecol. Monogr. 72(1):41-56. |
|
|
Ceulemans R, Janssens I, Jach M (1999). Effects of CO2Enrichment on Trees and Forests: Lessons to be Learned in View of Future Ecosystem Studies. Ann. Bot. 84(5):577-590. |
|
|
Chiarucci A, Bacaro G, Scheiner SM (2011). Old and new challenges in using species diversity for assessing biodiversity. Philos. Trans. R. Soc. Lond. B Biol. Sci. 366(1576):2426-2437. |
|
|
Chidumayo EN, Gumbo ENCDJ, Gumbo DJ (2010). The Dry Forests and Woodlands of Africa: Managing for Products and Services. Earthscan 305p. |
|
|
Colwell RK (2013). EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples. Version 9. |
|
|
De Cáceres M, Jansen F (2015). indicspecies: Relationship Between Species and Group of Sites, R Package Version 1.7. 5. |
|
|
De Cáceres M, Legendre P (2009). Associations between species and groups of sites: indices and statistical inference. Ecology, 90(12):3566-3574. |
|
|
De Kesel A, Codjia J, Yorou S (2002). Guide des Champinons Comestibles du Bénin. Centre International d'Ecodéveloppement Intégré (CECODI). Coco-Multimedia, Cotonou, République du Bénin, National Botanic Garden, Belgium. |
|
|
Dickie IA, Koele N, Blum JD, Gleason JD, McGlone MS (2014). Mycorrhizas in changing ecosystems. Botany, 92(2):149-160. |
|
|
Dissing H, Lange JMC (1963). Gasteromycetales. Ministère de l'agriculture, Jardin botanique de l'État. |
|
|
Duchaufour P, Blum WE (1997). Abrégé de pédologie: sol, végétation, environnement. Masson. |
|
|
Ducousso M, Louppe D, Ouattara N, Eyssartier G, Buyck B (1999). Des mycorhizes très diversifiées dans les jachères naturelles au nord de la Côte d'Ivoire. La jachère en Afrique tropicale: rôle, aménagements, alternatives, Dakar, Sénégal. |
|
|
Dufrêne M, Legendre P (1997). Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecol. Monogr. 67(3):345-366. |
|
|
ectomycorrhizal fungal sporocarps under increased atmospheric CO2 and O3. Ecol. Lett. 12(8):813-822. |
|
|
Eyi Ndong H, Degreef J, De Kesel A (2011). Champignons comestibles des forêts denses d'Afrique Centrale. In: Samyn Y, VandenSpiegel D, Degreef J (eds.) Taxonomie et identification. ABC Taxa. |
|
|
Gange AC, Gange EG, Sparks TH, Boddy L (2007). Rapid and Recent Changes in Fungal Fruiting Patterns. Science, 316(5821):71-71. |
|
|
Gardes M, Bruns TD (1993). ITS primers with enhanced specificity for basidiomycetes - application to the identification of mycorrhizae and rusts. Mol. Ecol. 2(2):113-118. |
|
|
Gerakis A, Baer B (1999). A Computer Program for Soil Textural Classification. Soil Sci. Soc. Am. J. 63(4):807-808. |
|
|
Heim R (1955). Les Lactaires d'Afrique intertropicale (Congo belge et Afrique Noire francaise). Bulletin du Jardin Botanique de l'État Bruxelles, 25:1-91. |
|
|
Heinemann P (1954). Boletineae. Flore iconographique des champignons du Congo 3:50-80. |
|
|
Heinemann P (1959). Cantharellineae. Flore iconographique des champignons du Congo 8:153-165. |
|
|
Heinemann P (1966). Hygrophoraceae, Laccaria et Boletineae II (compléments). Flore iconographique des champignons du Congo, 15:279-308. |
|
|
Henkel TW, Aime MC, Chin MM, Miller SL, Vilgalys R, Smith ME (2012). Ectomycorrhizal fungal sporocarp diversity and discovery of new taxa in Dicymbe monodominant forests of the Guiana Shield. Biol. Conserv. 21(9):2195-2220. |
|
|
Hennenberg KJ (2004). Vegetation ecology of forest-savanna ecotones in the Comoé National Park (Cote d'Ivoire): Border and ecotone detection, core-area analysis, and ecotone dynamics. Mathematical and Natural Sciences PhD Thesis. University of Rostock Fakultäder, Rostock, pp. 101. |
|
|
Hennenberg KJ, Goetze D, Kouamé L, Orthmann B, Porembski S, Austin M (2005). Border and ecotone detection by vegetation composition along forest-savanna transects in Ivory Coast. J. Veg. Sci. 16(3):301-310. |
|
|
Högberg P, Piearce G (1986). Mycorrhizas in Zambian trees in relation to host taxonomy, vegetation type and successional patterns. J. Ecol. 775-785. |
|
|
Hollister RD, Webber PJ, Bay C (2005). Plant response to temperature in northern Alaska: implications for predicting vegetation change. Ecology, 86(6):1562-1570. |
|
|
Horton TR, Bruns TD (2001). The molecular revolution in ectomycorrhizal ecology: peeking into the black-box. Mol. Ecol. 10(8):1855-1871. |
|
|
Hui W, Gel YR, Gastwirth JL (2008). lawstat: An R Package for Law, Public Policy and Biostatistics. 28(3):26. |
|
|
Hyvönen R, Ågren GI, Linder S, Persson T, Cotrufo MF, Ekblad A, Freeman M, Grelle A, Janssens IA, Jarvis PG, Kellomäki S, Lindroth A, Loustau D, Lundmark T, Norby RJ, Oren R, Pilegaard K, Ryan MG, Sigurdsson BD, Strömgren M, van Oijen M, Wallin G (2007). The likely impact of elevated [CO2], nitrogen deposition, increased temperature and management on carbon sequestration in temperate and boreal forest ecosystems: a literature review. New Phytol. 173(3):463-480. |
|
|
Ingleby K, Last F, Mason P (1985). Vertical distribution and temperature relations of sheathing mycorrhizas of Betula spp. growing on coal spoil. For. Ecol. Manage. 12(3):279-285. |
|
|
IPCC (2014). IPCC Fifth Assessment Synthesis Report - Climate Change 2014 Synthesis Report. Intergovernmental Panel on Climate Change (IPCC), pp.116. |
|
|
Janssens IA, Freibauer A, Schlamadinger B, Ceulemans R, Ciais P, Dolman AJ, Heimann M, Nabuurs GJ, Smith P, Valentini R, Schulze ED (2005). The carbon budget of terrestrial ecosystems at country-scale - a European case study. Biogeosciences 2(1):15-26. |
|
|
Kauserud H, Heegaard E, Semenov MA, Boddy L, Halvorsen R, Stige LC, Sparks TH, Gange AC, Stenseth NC (2010). Climate change and spring-fruiting fungi. Proc. R. Soc. Lond. B: Bio. Sci. 277(1685):1169-1177. |
|
|
Kauserud H, Stige LC, Vik JO, Økland RH, Høiland K, Stenseth NC (2008). Mushroom fruiting and climate change. Proc. Natl. Acad. Sci. 105(10):3811-3814. |
|
|
Körner C, Asshoff R, Bignucolo O, Hättenschwiler S, Keel SG, Peláez-Riedl S, Pepin S, Siegwolf RTW, Zotz G (2005). Carbon Flux and Growth in Mature Deciduous Forest Trees Exposed to Elevated CO2. Science 309(5739):1360-1362. |
|
|
Koulibaly A (2008). Caractéristiques de la végétation et dynamique de la régénération, sous l'influence de l'utilisation des terres, dans des mosaïques forêts-savanes, des régions de la Réserve de Lamto et du Parc National de la Comoé, en Côte d'Ivoire. PhD thesis, UFR Biosciences, Botanical Laboratory, University of Cocody, Abidjan, 111p. |
|
|
Krebs CJ, Carrier P, Boutin S, Boonstra R, Hofer E (2008). Mushroom crops in relation to weather in the southwestern Yukon. Botany, 86(12):1497-1502. |
|
|
Krebs CJ, Kenney AJ (2002). Programs for ecological methodology, 2nd Ed. version 6.1. Department of Zoology, University of British Columbia, Vancouver, B. C. Canada. |
|
|
Kruskal W.H. & Wallis W.A., 1952. Use of Ranks in One-Criterion Variance Analysis. J. Am. Stat. Assoc. 47(260):583-621. |
|
|
Lauginie F (2007). Conservation de la nature et aires protégées en Côte d'Ivoire. CEDA/NEI. |
|
|
Lawton R (1978). A study of the dynamic ecology of Zambian vegetation. J. Ecol. pp. 175-198. |
|
|
Maba DL, Guelly AK, Yorou NS, Verbeken A, Agerer R (2013). Two New Lactifluus species (Basidiomycota, Russulales) from Fazao Malfakassa National Park (Togo, West Africa). Mycol. Prog. 13(3):513-524. |
|
|
McGuire KL (2007). Common ectomycorrhizal networks may maintain monodominance in a tropical rain forest. Ecology, 88(3):567-574. |
|
|
Montoya JM, Raffaelli D (2010). Climate change, biotic interactions and ecosystem services. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 365(1549):2013-2018. |
|
|
Moore D, Robson GD, Trinci AP (2011). 21st Century Guidebook to Fungi with CD. Cambridge University Press. |
|
|
Morin X, Augspurger C, Chuine I (2007). Process-based modeling of species' distributions: what limits temperate tree species' range boundaries? Ecology 88(9):2280-2291. |
|
|
Nara K, Nakaya H, Hogetsu T (2003). Ectomycorrhizal sporocarp succession and production during early primary succession on Mount Fuji. New Phytol. 158(1):193-206. |
|
|
O'Dell TE, Ammirati JF, Schreiner EG (2000). Species richness and abundance of ectomycorrhizal basidiomycete sporocarps on a moisture gradient in the Tsuga heterophylla zone. Can. J. Bot. 77(12):1699-1711. |
|
|
Oksanen J, Blanchet FG, Kindt R, Legendre P, Minchin PR, O'Hara RB, Simpson GL, gy Package. R pSolymos P, Stevens MHH, Wagner H (2015). vegan: Community Ecoloackage version 2.0-10. P 274. |
|
|
Onguene N, Kuyper T (2002). Importance of the ectomycorrhizal network for seedling survival and ectomycorrhiza formation in rain forests of south Cameroon. Mycorrhiza, 12(1):13-17. |
|
|
Onguene NA (2000). Diversity and dynamics of mycorrhizal associations in tropical rain forests with different disturbance regimes in South Cameroon. Landbouwuniversiteit Wageningen (Wageningen Agricultural University). |
|
|
Onguene NA, Kuyper TW (2012). Habitat and diversity of ectomycorrhizal fungi in forests of South Cameroon. Cameroon J. Exp. Biol. 8(1):26-34. |
|
|
Orwin K.H., Kirschbaum M.U., St John M.G. & Dickie I.A., 2011. Organic nutrient uptake by mycorrhizal fungi enhances ecosystem carbon storage: a modelâ€based assessment. Ecol. Lett. 14(5):493-502. |
|
|
Pickles BJ, Egger KN, Massicotte HB, Green DS (2012). Ectomycorrhizas and climate change. Fungal Ecol. 5(1):73-84. |
|
|
Piepenbring M (2015). Introduction to Mycology in the Tropics. United States of America: APS PRESS, P 366. |
|
|
Poilecot P, Bonfou K, Dosso H, Lauginie F, N'dri K, Nicole M, Sangare Y (1991). Un écosystème de savane soudanienne: le Parc National de la Comoé (Côte d'Ivoire), IVC/87/007. UNESCO. |
|
|
Riviere T, Diedhiou AG, Diabate M, Senthilarasu G, Natarajan K, Verbeken A, Buyck B, Dreyfus B, Bena G, Ba AM (2007). Genetic diversity of ectomycorrhizal Basidiomycetes from African and Indian tropical rain forests. Mycorrhiza, 17(5):415-428. |
|
|
Sanon KB, Bâ AM, Dexheimer J (1997). Mycorrhizal status of some fungi fruiting beneath indigenous trees in Burkina Faso. For. Ecol. Manage. 98(1):61-69. |
|
|
Simard S. & Austin M., 2010. The role of mycorrhizas in forest soil stability with climate change. INTECH Open Access Publisher. |
|
|
Singh LP, Gill SS, Tuteja N (2011). Unraveling the role of fungal symbionts in plant abiotic stress tolerance. Plant Signal. Behav. 6(2):175-191. |
|
|
Smith SE, Read DJ (2008). Mycorrhizal Symbiosis. London, UK: Academic press, P 800. |
|
|
Tedersoo L, Bahram M, Jairus T, Bechem E, Chinoya S, Mpumba R, Leal M, Randrianjohany E, Razafimandimbison S, Sadam A (2011). Spatial structure and the effects of host and soil environments on communities of ectomycorrhizal fungi in wooded savannas and rain forests of Continental Africa and Madagascar. Mol. Ecol. 20(14):3071-3080. |
|
|
Tedersoo L, Nara K (2010). General latitudinal gradient of biodiversity is reversed in ectomycorrhizal fungi. New Phytol. 185(2):351-354. |
|
|
Van Der Heijden MGA, Horton TR (2009). Socialism in soil? The importance of mycorrhizal fungal networks for facilitation in natural ecosystems. J. Ecol. 97(6):1139-1150. |
|
|
Van Der Heijden MGA,Bardgett RD, Van Straalen NM (2008). The unseen majority: soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 11(3):296-310. |
|
|
Vargas R, Baldocchi DD, Querejeta JI, Curtis PS, Hasselquist NJ, Janssens IA, Allen MF, Montagnani L (2010). Ecosystem CO2 fluxes of arbuscular and ectomycorrhizal dominated vegetation types are differentially influenced by precipitation and temperature. New Phytol. 185(1):226-236. |
|
|
Vavrek MJ (2011). Fossil: palaeoecological and palaeogeographical analysis tools. Palaeontologia Electronica, 14(1)1. |
|
|
Verbeken A, Walleyn R (2010). Vol. 2: Monograph of Lactarius in tropical Africa. Meise: National Botanic Garden of Belgium. |
|
|
Walther GR, Post E, Convey P, Menzel A, Parmesan C, Beebee TJC, Fromentin JM, Hoegh-Guldberg O, Bairlein F (2002). Ecological responses to recent climate change. Nature, 416(6879):389-395. |
|
|
WMO W (2007). Scientific assessment of ozone depletion: 2006. World Meteorological Organisation, Global Ozone Research and Monitoring Project–Report, 50:572. |
|
|
Yorou NS (2010). Higher mushrooms. In: Sinsin B. & Kampmann D. (eds.). Biodiversity Atlas of West Africa. Cotonou & Frankfurt/Main. |
|
|
Yorou NS, Koné NGA, Guissou ML, Guelly AK, Maba LD, Ékué MRM, De Kesel A (2014). Biodiversity and Sustainable Use of Wild Edible Fungi in the Sudanian Centre of Endemism: A Plea for Valorisation. In: Bâ AM, McGuire KL, Diédhiou AG (eds.). Ectomycorrhizal Symbioses in Tropical and Neotropical Forests. pp. 241. |
|
|
Yorou S, De Kesel A, Sinsin B, Codjia J (2001). Diversité et productivité des champignons comestibles de la forêt classée de Wari-Maro (Bénin, Afrique de l'Ouest). Syst. Geogr. Plants 71:613-625. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


