ABSTRACT
Tree species composition and diversity were assessed in Agoro-Agu Central Forest Reserve located in Lamwo district-Northern Uganda. Fifteen transects, each 2 km long, were established in the reserve. A procedure of nested quadrats was employed for a systematic sampling of mature trees, saplings and seedlings at 100 m interval(s). Vegetation analysis was conducted in Species Diversity and Richness (SDRiv) software. A total of 100 species of trees belonging to 36 families were recorded. The dominant tree species, also with high Importance Value Indices were Combretum molle G. Don, Albizia grandibracteata Taub. and Vachellia hockii De Wild. Tree species diversity was higher in the intact part (H’=3.46) than in the degraded part (H’=2.86). Lower diversity in the degraded part of the reserve could be due to human disturbance and resulting conducive condition for the early establishment of pioneer species during succession. The occurrence of mixed species with indicator species for both savannah woodlands and Afromontane tree species indicates that Agoro-Agu is an enriched forest for enhanced conservation. Management options that limit continuous degradations such as the enforcement of environmental laws, tree enrichment planting, and ex-situ conservation should be undertaken to restore the status of trees in the degraded part of the reserve.
Key words: Afromontane, biodiversity, East Africa, forest conservation, national forestry authority and species richness.
Afromontane forests are forests that occupy the mountainous landscapes of East and West Africa (Mairal et al., 2017). In East Africa, Afromontane forests are widely scattered but geographically similar. Such forests often share similar features of mixed trees and plant species which are distinct from the surrounding lowlands (Gadow et al., 2016). The Afromontane altitudinal geographical limit falls within the range of 1000 to 2700 m above sea level (UNEP-WCMC, IUCN, 2020). Typically occurring above 1500 m in elevation, Afromontane forests extend from the Arabian Peninsula south along the rift to the Drakensberg Mountains in the east. The mountains are rich in biodiversity and are centers of endemism with many range-restricted species (Abiem et al., 2020).
Agoro-Agu Central Forest Reserve is located at the border between Uganda and South Sudan. It is the extension of Imatong Mountains into the Northern Region of Uganda, and basically lower than the southern part in South Sudan (Zhao et al., 2018). In Uganda, Afromontane forests include among others Bwindi forests and mid elevation forests at the border of Republic of South Sudan. The forest reserves of Imatong hills in South Sudan and Agoro-Agu in Northern Uganda are some of the outlying patches of the Eastern Arc Afromontane vegetation (Newmark and McNeally, 2018). Studies conducted in other Afromontane regions showed that such Afromontane outliers are hot spot areas with high diversity of flora and fauna but are often neglected (Kidane et al., 2019).
The high elevations in South Sudan that adjoins Agoro-Agu Central Forest Reserve in Northern-Uganda border are equally known to harbour a mixture of various kinds of forests including Afromontane forests, shrub lands, woody grasslands and bamboo - Sinarundinaria alpina (Zhao et al., 2018; Omoding et al., 2020). The distribution of East African Montane, its extension and elevations in South Sudan and Uganda border form part of the medium-altitude archipelago landscapes of the East African Afromontane for which Agoro and Agu hills are part of the landscape covered by Agoro-Agu Central Forest Reserve. It should be noted, however, that montane forests such as the hills of Agoro-Agu currently covered by the Agoro-Agu Central Forest reserve, have had very little attention in the past; yet they are becoming increasingly important for the conservation of fauna and flora (Omoding et al., 2020).
Agoro-Agu Central Forest Reserve has been reported to be extensively encroached with both temporary and semi-permanent housing, gardens, sambas and schools throughout the lower and medium altitude areas, particularly in the southern and eastern areas. Most of the disturbance and settlements were concentrated in the mid slope area where the landscape remains relatively flat. The human activities included settlements, cultivation, cutting of trees for fuel wood/building poles, timber, and rearing of domestic livestock including dogs, chicken, goats and cattle (NFA, 2018).
The situation was exacerbated by the presence of both the Lord’s Resistance Army (LRA) and the Sudanese People’s Liberation Army (SPLA) who operated within the reserve for many years. Despite the aforementioned, this forest reserve remains a source of building poles (especially bamboo), honey, bush meat, medicinal plants and firewood to the adjacent communities. The status quo to date indicates high level of encroachment from the resettling population as people return from captivity (Environmental Alert, 2017).
In the last 20 years or so, civil wars have negatively affected highlands and mid-altitudes forests in Northern Uganda including Agoro-Agu. The Agoro-Agu Forest Reserve suffered from persistent conflict in the mid-1980s, including the Lord’s Resistance Army civil war against Uganda’s Government and conflicts in neighbouring South Sudan. The Agoro-Agu Forest Reserve (264 km2) established in 1937, and gazetted as a Central Forest Reserve (CFR) in 1948 is part of a trans boundary protected area complex with South Sudan’s Imatong Forest Reserve. Apart from massive areas being converted to farmland, trees also got continuously cut for firewood and building poles by the resettling population (NFA, 2018).
To understand the rate of forest cover loss, a precise, fine scale demographic data would be needed to fully understand where returnees settled and how their land use practices have affected forest cover in Agoro-Agu Central Forest Reserve. Assembling information on tree species diversity and composition is not only key in understanding important aspects of tropical forest ecosystem, but can also help in determining the diversity and structure of the tropical forests ecosystems than any other living component of the system (Mori et al., 2017).
Furthermore, having a proper data on patterns of tree species diversity and composition is a prerequisite if any meaningful management interventions related to conservation of tropical forests are to be carried out (Gebeyehu et al., 2019). Apart from providing the necessary context for planning and interpreting long-term ecological research (Haruna et al., 2018), data from such a study would be equally relevant to Agoro-Agu Central Forest Reserve given the current pressure from resettling war returnees and the need to making choices and decisions for biodiversity conservation in this forest landscape.
Much as the tropical flora remains chronically understudied, thousands of plant and animal species in tropical regions provide a variety of non-timber products that are used by billions of people all over the world. Indeed conservation and long term utilization of these species require that they be harvested on a sustainable basis (Young et al., 2017). This implies that understanding tree species diversity and composition patterns in such a reserve would be of utmost interest to conservationists when interpreting long-term ecological data (Gonçalves et al., 2017) and monitoring changes in ecosystem health, as well as the composition of flora and fauna (Strassburg et al., 2019).
Policy makers, conservationists, the public and other professional (Planners, foresters, health workers, help researchers, etc.) would also need such information to them understand the implication of environmental and population trends including designation of appropriate conservation actions to be taken (Gavin et al., 2018).
Even if Uganda Forest Department surveyed and recorded 254 useful trees and shrubs in Agoro-Agu with moderate to high levels of biodiversity, the study was limited in scope due to the insecurity situation within the reserve (NFA, 2018). Difficulty of access to an area during war combined with no clear spatial or temporal definition for the extent of conflict also made it extremely challenging to conduct accurate and timely assessment of the impacts of wars on tree species composition. Satellite image analysis on the same forest reserve by Gorsevski et al. (2012) was also reported only on slight decreasing forest and increasing tree cover of the reserve with no details on tree diversity and composition.
These scenarios, thus, necessitated an assessment of tree species diversity and composition in the reserve. Such knowledge of floristic composition and diversity is useful for conservation by identifying ecologically and economically important plants and their diversities. The information on plant diversity is also required for facilitating proper planning and management of biodiversity, and ecosystem services provided in such a reserve. Indeed long-term biodiversity conservation also depends basically on the knowledge of the structure, species richness, and the ecological characteristics of vegetation (Ifo et al., 2016).
Location and size of the study area
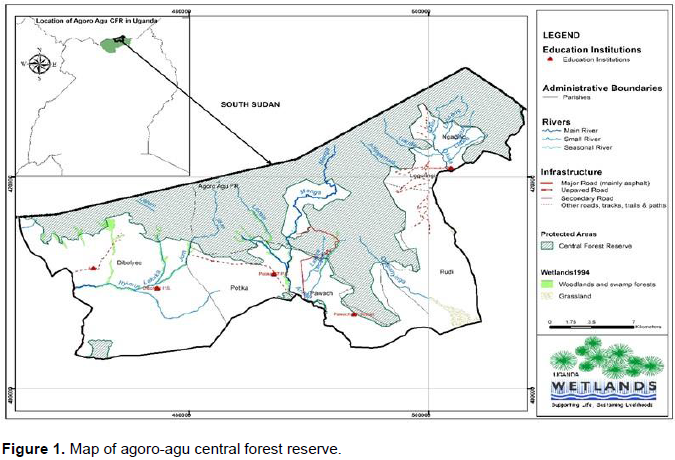
Agoro-Agu forest landscape falls within the Agoro-Agu Sector that covers 17 central forest reserves in Aswa river range. The Agoro-Agu Central Forest Reserve is a savanna woodland reserve considered among patches of diverse Eastern Afromontane landscape in the region; at the border between Uganda and South Sudan. It is the extension of Imatong Mountains into the Northern Region of Uganda, and basically lower than the southern part in South Sudan (Zhao et al., 2018). Agoro-Agu is located in Lamwo District, Northern Uganda between 3°40-3°53′ N and 32°42′-33°04 E (Figure 1). It has at altitudinal range of 1100 to 2700 masl, and at a distance of 463 km from Kampala city and covers a total of 26,508 ha (IUCN, 2015).
According to National Forestry Authority, the reserve covers a total area of 236 km2/26,508 ha (NFA, 2018). This constitutes 2.9% of the total tropical forest area coverage in Uganda. The forest reserve lies along the border between Uganda and Republic of South Sudan.
Description of the study site
Physiographically, Agoro-Agu Central Forest Reserve is characterized by hills, escarpments, valleys, relatively flat mid slopes and isolated ridges. The forest reserve is constituted by a series of small hills whose peaks are marked by Lumwaka hills (3100 m) in the North and Agu hills (1810 m) in the North-West and Agoro hills (2850 m) in the eastern part of the forest reserve (Olanya, 2020b).
These three hills stretch across the international border of Uganda and the Republic of South Sudan (Environmental Alert, 2017). Apart from being connected to the Imatong hills, other small hills in the reserve include: Talamudu, Lowa matidi, Lowa madit, Podolo, Loturtur, Lacer, Lamwo, Alila and Katomo hills (Zhao et al., 2018). Agoro-Agu Central Forest Reserve had for the last two decades served as a rebel hideout which rendered it inaccessible to the conservation authority (Omoding et al., 2020).
Vegetation characteristics
The vegetation of Agoro-Agu Central Forest Reserve has been broadly classified as dry Combretum savanna, forest-savanna mosaic and dry montane forest. Accordingly, land cover map for Agoro-Agu indicates that the vegetation of this area includes Afromontane forests, shrub lands, woody grasslands and bamboos (Zhao et al., 2018).
According to NFA Master Plan, about 100 km2 (38%) of the reserve is occupied by forest/savanna mosaic at high altitudes. About 71 km2 (29%) is occupied by Combretum-Acacia-Themeda savanna. Juniperus-Podocarpus trees typical of dry montane forest occupies 48 km2 (20%), of the reserve landscape while Cyperus papyrus Swamp; Acacia-Cymbopogon-Themeda grassland and Butyrospermum-Hyparrhenia savanna occupies 4, 2 and 2% of Agoro-Agu forest landscape, respectively. The 16 CFRs within the Agoro-Agu Landscape cover 65,548 ha under one Forest Management Planning Area (the Agoro-Agu Sector) which are managed with a common Forest Management Plan (NFA, 2018).
Climate and topography
The climate of Agoro-Agu Central Forest Reserve is tropical Afromontane and rainfall is fairly constant throughout the season with warm day time temperatures up to 35°C that cools to 15°C at night (at Katire, 800 masl). Precipitation increases with altitude, resulting in a transition of the montane forest zone to the alpine zone. Rainfall across the study region is seasonal with the rainy season beginning in late March and lasting until the end of October (Gorsevski et al., 2013). The mountains in Agoro-Agu Central Forest Reserve consist of granitic crystalline rocks, most of which are folded and foliated with soils largely falling within the following four categories: (i) dark cracking clays, (ii) non-cracking clays, (iii) red loam and ironstone soils, and (iv) hill or mountains soils (Uganda Land Use Dialogue, 2020).
Conservation issues
Agoro-Agu Central Forest Reserve is of relatively high conservation value for all taxa compared to other forest reserves in Northern Uganda. The forest is contiguous with the Imatong Mountains of south Sudan and known for their high rates of endemism and unique species composition. In northern Uganda, it is among the 15 forest reserves that were surveyed and reported to have had at least one species found in no other protected area in Uganda. Although a study commissioned by the Ugandan Forest Department recorded moderate to high levels of biodiversity; extensive encroachment evidenced in the lower and medium altitude areas raises conservation concern (NFA, 2018).
Data collection
Research design
The study followed a design by Delgado-Aguilar et al. (2017) in which the naturalness or the integrity of the ecosystem and human disturbance features were used as a basis for landscape description of intact (non-degraded) and disturbed (degraded) landscapes. The criterion therefore described intact area as an unbroken expanse of natural ecosystems within the reserve with no signs of significant human activity. According to Haurez et al. (2017), intact site usually consists of forested, treeless, grassland or non-forest ecosystems minimally influenced by human economic activity. Based on the two descriptions, two study sites were identified in the study area.
While the disturbed/degraded areas consisted of relatively flat mid slope areas with settlements, cultivation zones had significant human activities (e.g. fallow lands or previously cultivated lands and anthropogenic evidences like remnants of tree cuttings and stumps); the intact part had features similar to those described by Haurez et al. (2017), compared to the intact part of the forest reserve. This was based on the idea that the degraded section of the forest reserve constituted less than 5% of the total reserve area. The vegetation community landscape was furthered categorized and subdivided into three topographic positions that were defined as Ridge top, Mid slope and Valley bottom topographic positions following Mligo (2018).
Using the two categories of forest parts and three topographic positions of the landscape in the assessment of tree species diversity, structure and composition, a total of 12.7 ha of the forest reserve was surveyed consisting of 9.5 ha of the intact part and 3.2 ha of degraded part of the forest reserve. Efforts were made to select plot locations based on representation of possible aspect and slope topographic position combinations.
Reconnaissance study
Fieldwork started with a preliminary visit (reconnaissance) to the forest reserve. This was intended to plan the stratification of sample sites based on naturalness and intactness or extent of human disturbance on the forest reserve. The disturbance evidence considered included human activities like crop cultivation, tree stumps and settled areas. The non-degraded areas were considered to be at least 1 km away from the motorable paths and half a kilometer from the foot paths. This guided the laying of transects and establishment of plots during data collection (Solomon et al., 2018).
Transect establishment and plot lay out
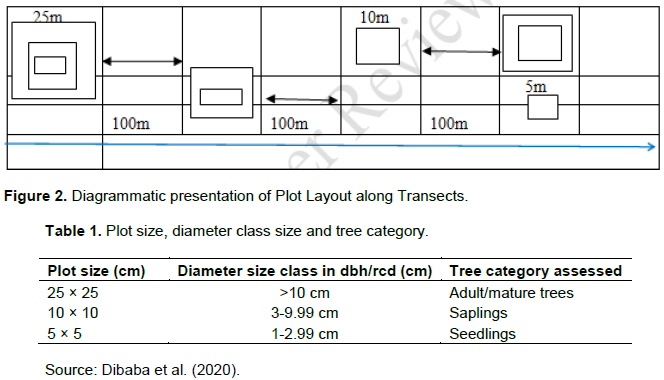
A total of 15 transects were laid throughout the study area; nine in the intact part and six in the degraded part of the forest reserve. Transects (2 km long) were laid and on each; nested quadrants (alternating on either side of each transect) were laid out systematically at 100 m intervals. The inter-transect distance was 600 m and the inter-plot distance was 100 m (Figure 2).
Global Positioning System (GPS), compasses and 50 m ribbon tape measure were used to mark the plots along transects. A compass bearing was used to indicate direction during plot setting. Within the 25 m × 25 m plot, smaller plots of 10 m × 10 m and 5 m × 5 m were nested for sampling mature trees, saplings and seedlings, respectively (Dibaba et al., 2020).
In this study, seedlings were considered as young trees with root collar diameter (rcd) between 1 and 2.9 cm and the data were obtained from the 5 m × 5 m plots; saplings were considered to be of diameter at breast height (Dbh) range from 3 to 9.9 cm and the data were collected from 10 m × 10 m plots. Mature trees were considered as trees with Dbh ≥10 cm and the data were recorded from 25 m × 25 m plots throughout the inventory (Dibaba et al., 2020). The total number of plots for the entire sampled area for both intact and degraded parts of the reserve was 203 of which 152 were in the intact and 51 were in the degraded part of the forest reserve.
Assessment of tree species composition and diversity
At each plot center, GPS coordinate readings were recorded. Following Dibaba et al. (2020), measurements of Dbh of mature trees and saplings were carried out at 1.3 m from ground surface and root collar diameters (rcd) for seedlings (Table 1).
For the tree trunk with buttresses near breast height, the Dbh were measured just above the buttress. Tree species were assessed in the 25 m × 25 m plots where the diameter at breast height (Dbh) of the trees were measured using either Vernier caliper (for smaller trees), tape measure (for very big trees) and recorded under different diameter class sizes (Table 1). Specimens of unfamiliar trees were collected and taken to Makerere University Herbarium for proper identification.
Data analysis
Vegetation data were entered and analysis conducted in Species Diversity and Richness (SDRiv) software (Kacholi, 2019; Birhanu et al., 2021).
Analysis of tree species composition
Tree species composition was considered to be an assemblage of tree species that characterized the forest vegetation (Ifo et al., 2016). Tree species encountered in each plot were counted, and the composition measures in Equations 1 to 5 were considered in the calculation of Important Value Species Indices (IVI): Density (D), Frequency (F), Relative Density (RD), Relative Frequency (RF) and Relative Dominance (RDn) following methods by Tolangara et al. (2019) and Reshad et al. 2020).
IVI= Relative frequency + Relative density + Relative dominance (6)
Every species was ranked according to their importance values and the species with the highest importance value in the stand was considered as trees with the highest value that existed in the greatest number or are of the greatest size, dominant species and may have the greatest effect on the community (Asigbaase et al., 2019). Thus, the index reflected the overall importance of each species within the community. This provided a basis for highlighting species of conservation significance (Gebrewahid and Abrehe, 2019).
Analysis of tree species diversity
Species diversity has two components: richness and evenness. Species richness is the number of species in a sample while evenness refers to their relative abundance. Species Diversity and Richness (SDRiv) software (Tecimen et al., 2017) was used to carry out the analyses of tree species diversity and richness indices for Degraded and Intact Forest Reserve. The same analysis was done for tree species diversity indices and richness along topographic positions (ridge top, mid-slope and slope bottom).
To assess the woody species diversity in the forest reserve, the Shannon-Wiener diversity index (an information statistic index, which assumes that all species are represented in a sample and that they are randomly sampled) was used (Joshi and Dhyani, 2019).
Its equation is:
H′ = −ΣPi×(ln Pi) (7)
where H′ = is the Shannon Diversity index, Pi = is the importance value of a species as a proportion of all species (that is, the proportion (n/N) of individuals of one particular species found (n) divided by the total number of individuals found (N), ln = is the natural log, Σ = is the sum of the calculations, and s is the number of species.
Shannon Tree Species Diversity and Richness (SDR) were calculated as follows (Equation 8):
where D= Simpson Diversity Index, ni is the number of individuals of ith species and N is the total number of individuals.
The Simpson index is a dominance index because it gives more weight to common or dominant species. In this case, a few rare species with only a few representatives will not affect the diversity.
Pooled analysis using rarefaction method
Rarefaction technique was also used to standardize samples from intact and degraded parts of the forest reserve; and also sample sizes along topographic locations (ridge top, mid-slope and slope bottom) that differed in terms of sizes. The standardization was derived from hyper geometric distribution whose outputs were hyperbolic curves depicting Species Accumulation Curves for the various sample plots (Jenerette et al., 2016).
Pairwise analysis using Sørensen and Jaccards Similarity Index (JSI)
Jaccard’s Similarity Index was used based on the presence/absence of species that were shared and uniqueness between samples of tree species (Rahman et al., 2019). For this study, analysis of similarity was carried out between intact verses degraded parts of the reserve; and between topographic positions.
Thus, in measuring the similarity indexes, Sørensen Similarity Index was calculated from (Equation 10):
Cs = 2a / (2a + b + c); and Jaccards Similarity Index was calculated from:
Cj = a / (a + b + c) (10)
where c = represented the number of species found in site A (that is, Intact part of the reserve); b = represented the number of species in site B (that is, degraded part of the reserve) and a = represented the number of species shared by the two sites, that is, intact and degraded parts of the reserve (Santini et al., 2017).
Tree species composition
The inventoried trees of Agoro-Agu Central Forest Reserve were composed of 100 species belonging to 36 families. The most abundant families in the intact part of the reserve were Fabaceae with 20 species, Moraceae and Euphorbiaceae with eight species each, and Anacardiaceae and Rubiaceae with five species each. These were followed by Combretaceae and Meliaceae each with four species, Canellaceae, Celastraceae and Sterculiaceae with three species each and the rest of the families had either two or one species each (Table 2).
While Fabaceae was the most diverse; followed by Moraceae, Euphorbiaceae, Anarcaidaiceae, Rubiaceae and Combretaceae with a minimum of 4 species each (in intact part), Fabaceae was the most diverse; followed by Combretaceae, Moraceae, Euphorbiaceae, and Anarcaidaiceae, with 3 species each in degraded parts of the reserve. The rest of the families had either one or two species only (Table 2).
Of the encountered 36 families, the first ten families (Table 2) contributed 74 and 64.2% of the total species in the intact and degraded parts of the reserve, respectively. The remaining 26 families accounted for only 26 and 35.8% of the total tree species in the intact and degraded parts of the reserve, respectively.
Eleven families that were exclusively observed in the intact part of the reserve included Ulmaceae, Podocarpaceae, Sterculiaceae, Sapindaceae, Polygalaceae, Phillanthoceae, Ochnaceae, Meliaceae, Apiaceae, Bignoniaceae, Celastraceae and Ebenaceae (Table 2).
Importance value indices of tree species
Important value indices (IVIs) provide knowledge on important species of the tree community in the reserve. The list of species and their IVIs for first 10 most important species are shown in Tables 3 and 4.
Based on IVI, the most important species in the intact part of the reserve were Combretum molle, Acacia hockii, Albizia grantibracteata and Entada abyssinica; while Combretum collenum and Terminalia glaucescens were the least ranked species among the first 10 species in the intact part of the reserve (Table 3).
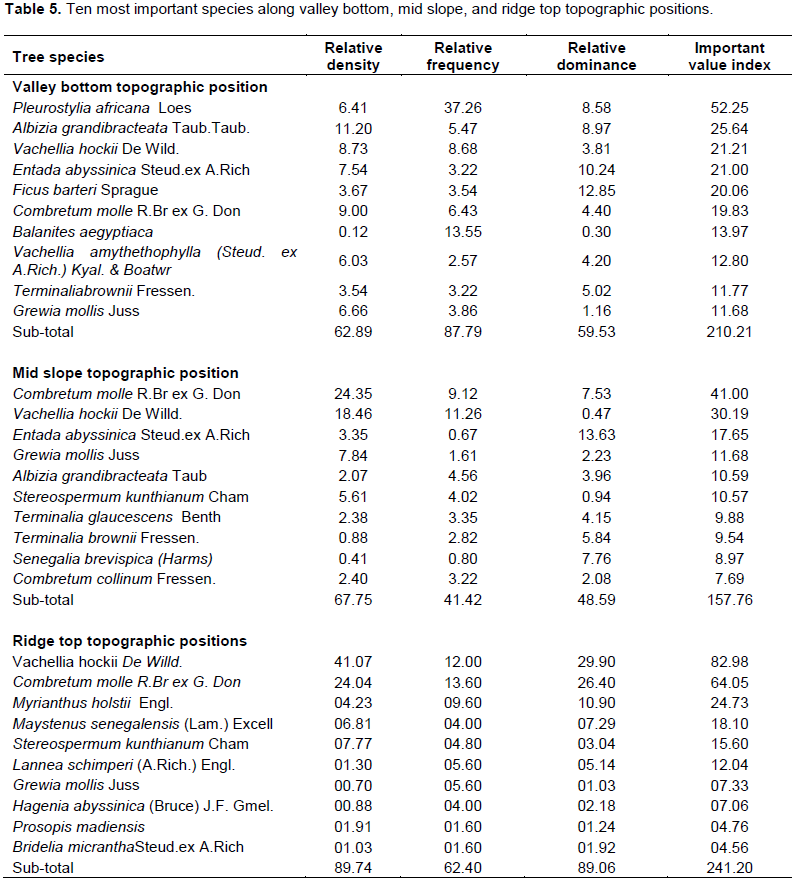
On the other hand, C. molle, A. hockii, T. glaucescens, A. grandibracteata, Grewia mollis and Ficus sycamore were some of the most important species in the degraded part of the reserve (Table 4).
C. molle and A. hockii had the highest IVIs, while Vitex doniana and Stereospermum kunthianum had the least IVIs (Table 4).
In general, the IVIs for the 10 species represented 52.8 and 70.7% of the total Importance Value Indices for the intact and degraded parts of the reserve, respectively. Along topographic gradient, A. hockii, Pleurostylia africana and C. molle had the highest Importance Value Indices (IVIs) for Mid slope, Valley bottom and Ridge top topographic positions, respectively.
The summary for the species in order of their decreasing IVI is provided in Table 5a to c. While A. hockii had the highest IVI (82.98) followed by C. molle (64.05) in the bottom Ridge top topographic positions, P. africana (52.25) followed by A. grandibracteata (25.64) and C. molle (41.00) followed by A. hockii (30.19) had the highest IVIs in the Valley bottom and Mid slope topographic positions, respectively (Table 5a to c).
Tree species diversity
Tree species richness
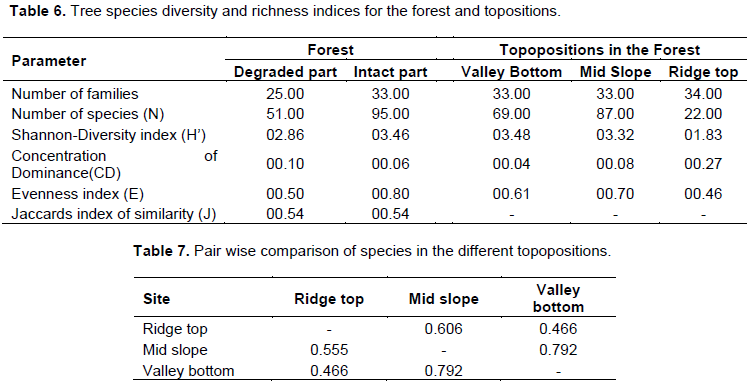
One hundred tree species were encountered throughout the study area. The intact part of the reserve had 95 tree species and was richer than the degraded part with only 51 tree species. Along topographic positions (valley bottom, mid slope and ridge top), mid slope was the richest, followed by valley bottom and ridge top with 87,69 and 22 tree species, respectively (Table 6).
The species diversity given by Shannon-Diversity and Richness index was higher in the intact part of the reserve (H’=3.46) than in the degraded part (H’=2.86). The tree species diversity declined from valley bottom with (H’=3.48) to ridge top with (H’=1.83).
Tree species evenness
Pielou evenness index was highest for the intact part of the reserve (E=0.80) and lowest for degraded part (E=0.50). Similarly, mid slope had the highest evenness (E=0.70) and ridge top had the least evenness (E=0.46) among topopositions. The value for concentration of dominance was lowest for intact portion (0.06) and highest for the degraded (0.10) part of the reserve.
Along the topographic gradient, evenness value increased from valley bottom (0.04) to ridge top (0.27). While all values for concentration of dominance (cd) showed reverse trend as compared to Shannon diversity indices, species diversity and concentration of dominance appeared inversely related (Table 6).
Tree species similarity
The Jaccards Similarity Index showed an average of 50% similarity in species composition between the intact and degraded parts of the reserve. Along topographic positions, high similarity values were observed between mid- slope and valley-bottom; the least comparison was observed between ridge-top and other topographic positions (Table 7).
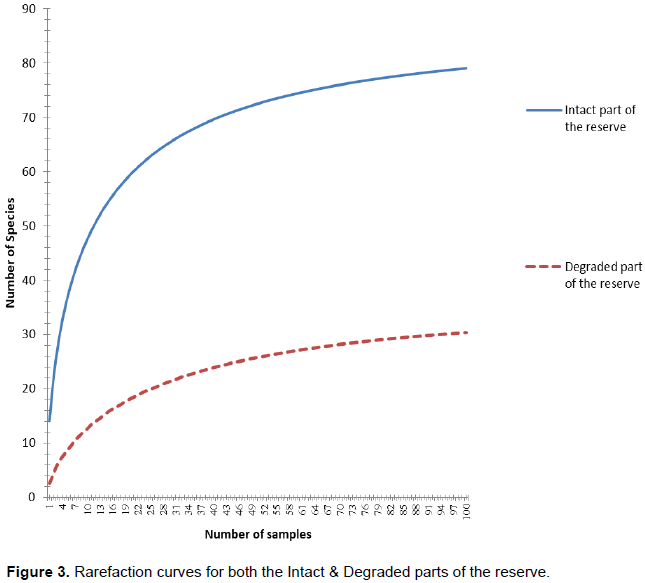
The rare faction and assemblages of the intact and degraded parts of the reserve including topopositions showed similar hyperbolic curves (asymptote) with maximum plateau shapes for all samples for both degraded and intact parts of the reserve (Figure 3).
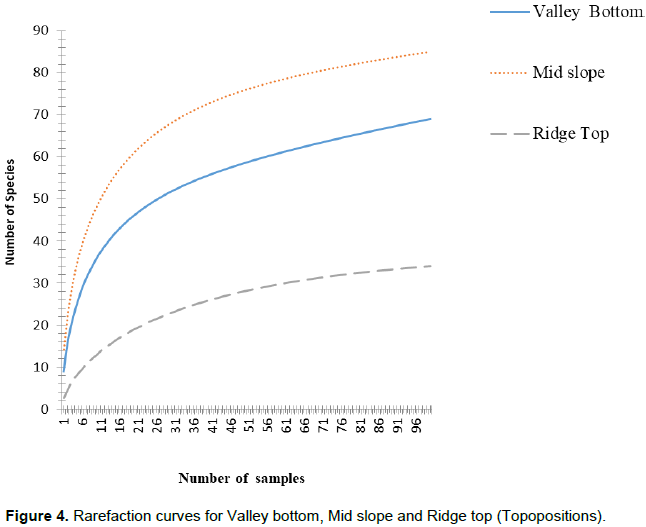
Similarly, along topographic position, the generated curves had hyperbolic shape which flattens out and eventually begins to level off. This is an indicator that the sampling captured maximum proportion of the species richness (Figure 4).
Tree species composition
The most common tree families recorded in the study included Euphorbiaceae, Moraceae, Combretaceae and Fabaceae which were well represented in both intact and degraded parts of the reserve. According to Ifo et al. (2016), such families are common in most savannah woodland mosaics in Africa and are typical of the Sudano-Sahelian zone. The dominance by Combretaceae, Euphorbiacea and Fabaceae are therefore good indicators for Sudano-sahelian vegetation composition (Dangulla et al., 2020).
On the contrary, families that included Ulmaceae, Podocarpaceae, Sterculiaceae, Sapindaceae, Polygalaceae, Phillanthoceae, Ochnaceae, Meliaceae, Apiaceae, Bignoniaceae, Celastraceae and Ebenaceae were not encountered in the degraded part of the reserve. It is possible that such families contributed most of the bigger trees that had been harvested for fuelwood, building poles and expansion of cultivation lands leading to compositional differences (Tenzin and Hasenauer, 2016). The most probable reasons for the harvest of such tree species could include among others factors like agricultural encroachment and increased demand for fuelwood (Ministry of Water and Environment, 2017; Kalema and Hamilton, 2020). Indeed Agoro-Agu Central Forest Reserve had been inhabited by the displaced people who exploited it for building poles, firewood, medicine, and bush meat (Omoding et al., 2020).
Extraction of these resources could have caused habitat fragmentation and loss in this protected area, altering its structure and functioning (Olanya, 2020a).
The encountered tree families that consisted of species such as Podocarpus latifolia, Lovoa trichilioides (African Walnut), Lovoa swynnertonii, Mimusops bagshawei, Pterygota mildbraedii, and Hagenia abyssinica are typical forest species. This could be attributed to (among other factors), the absence of single mature tree remnants to serve as a seed bank (Martínez?Ramos et al., 2016). Although, other scholars have maintained that the absence of such species may be due to micro-site factors like moisture, soil characteristic and landscape position (Fazlollahi et al., 2017), it is also possible that clearance of mature trees that serve as seed bank can affect recruitment from seeds (Chen et al., 2017). The removal of such seed sources could have certainly led to local extinction of certain tree species and subsequently altered tree species composition (Aynekulu et al., 2016; Ranjan, 2018).
Tree Importance Value Indices
In forest ecological studies, IVI indicates the ecological importance of a species in a community and provides an overview of the social structure of a species. IVIs also indicate dominance of species in mixed population and give a total picture and knowledge about important species and their composition in such a forest reserve (Siraj and Zhang, 2018). Since IVI is an important parameter that reveals the ecological significance of species in a given ecosystem, it reflects the combined effect of species density, frequency and dominance (Demies, 2019). Moreover, it enables prioritizing species for conservation interventions such that species with lowest IVIs might benefit from conservation and management interventions. Tree species such as C. molle, A. grandibracteata and A. hockii are species with high IVIs and constitute the dominant species in both portions of the reserve. Some of these species recorded with high IVIs are multipurpose in nature and are valued by many communities in Africa for fuel and medicines.
For example, C. molle and Terminalia species are documented as important medicinal species not only by communities in Uganda but also other parts in Africa (Hegazy et al., 2020). A. hockii is valued for construction purposes and medicine in treating tuberculosis and allied diseases (Nabasumba et al., 2016) in south Western Uganda. According to Asigbaase et al. (2019), such trees with high IVIs have more ecological advantage and require less conservation management efforts. This implies that conservation measure on the contrary should now be shifted to trees with low IVIs.
In this study, L. trichilioides and L. swynnertonii, which recorded low IVIs are listed as vulnerable and endangered. While species like P. latifolia and Strombosia scheffleri are considered nationally threatened species (MTWA, 2018); and L. swynnertonii is considered internationally threatened species (Migliore et al., 2020). Indeed some of these species with low IVI values in this reserve are included in the 2017 IUCN Plant Red list either as vulnerable or endangered; mainly due to high exploitation rates (IUCN, 2020) and being subjected to heavy felling (Mengich et al., 2020). Such important trees with low IVIs, would thus, require more attention and prioritization because they are probably less adaptive with least ecological advantage (Asigbaase et al., 2019).
Other species with low IVIs that are also enlisted under IUCN Red list as threatened or endangered species and that need to be considered for conservation are M. bagshawei, Polyscias fulva, P. mildbraedii and a synonym of H. abyssinica. Afzelia africana, L. trichiloides and Securidaca longipedunculata are also among the species of special conservation interest. This observation defines the ecological significance and the urgency needed in developing mechanisms for the restoration and conservation of Agoro-Agu Central Forest Reserve.
In order to understand where returnees are resettling and how their land use practices are affecting forest cover, a fine scale demographic data combined with a well-executed socio-economic surveys also would be necessary (Salazar et al., 2018). This is quite important given that these forests may not only provide timber and firewood, but is part of a larger water catchment system. The results suggest that the species having low IVI value should be prioritized for conservation. Accordingly, these lower IVIs indicate that these woody species are threatened and need immediate conservation measure (Demie, 2019). Since IVI is used for prioritizing conservation of species, species with low IVI index values should be given higher priority in conservation than those with high IVIs.
Tree species diversity
In a survey covering 12.7 ha, 5,936 stems of trees belonging to 100 tree species and 36 families have been encountered. The value for richness(R) and species diversity (H’) varied among sites (intact and degraded parts of the reserve) and along the topographic gradient as provided in Table 6. The variation in species richness and diversity in such sites could be attributed to physical heterogeneity (Shirima et al., 2016) and anthropogenic perturbation (Yuan et al., 2016). The intact part registers higher species richness and diversity (Table 6). This could be probably correlated with greater diversity and community stability in such a reserve.
According to Ifo et al. (2016) high species diversity indicates a highly complex community, for a greater variety of species that allows for a larger array of species interactions. The diversity index in intact part was significantly higher than degraded part perhaps due to the high number of families, number of genera, and number of species that were recorded in the intact compared to the degraded part of the reserve. As has also been reported by Arroyo?Rodríguez et al. (2017), the development and changes of floristic composition and diversity of plants during early stages of succession could have been mostly influenced by secondary succession processes and fallow periods in the intact and degraded parts of the reserve, respectively.
On the contrary, the degraded part of the reserve has lower species richness and diversity. This could probably be attributed to the impact of anthropogenic perturbations such as firewood harvesting, felling, lopping and clearing of land for cultivation and settlement (Ifo et al., 2016). Such disturbances are likely to impact on tree species richness and diversity of the reserve. In some cases, such disturbances could reduce not only the number of species in the ecosystem but also the number and extent of places where species coexist. According to Wegasie et al. (2018), the effects of such disturbance may even be more on certain tree species depending on the purpose for which the trees are needed.
In addition to anthropogenic disturbance, suppression by the pioneer species such as C. molle, A. hockii and A. grandibracteata may also not be ignored (Swinfield et al., 2016). According to Muluneh et al. (2021), pioneer species usually establish very early in succession and are able to suppress other species very rapidly. The recruitment of such species may probably account for the observed variation in species richness and diversity in both intact and degraded part of the reserve. As has also been reported by Yahya et al. (2019), the absence of certain species like P. latifolia, M. bagshawei, P. fulva, Pterygota mildbraedii, H. abyssinica, A. africana, and L. trichiloides from the degraded part of the reserve could be attributed to dominance by the pioneer species such as the genus Combretum.
Indeed, the genera Combretum and Acacia are recorded as light tolerant and have characteristics of dominating the previously disturbed savannah woodlands while also suppressing the regeneration of other tree species (Sangeda and Maleko, 2018). Apart from these, other pioneers species that are capable of growing to forest canopy within a decade to become important timber species have to be assessed and protected.
Although Shannon-Wiener’s diversity index is the most preferred index among the other diversity indices and values are between 0.0 and 5.0 with results generally in the range 1.5 to 3.5 and very rarely exceeding 4.5 (Deo et al., 2016), the overall diversity index value of 3.40 for Agoro-Agu Central Forest Reserve is equally considered high. This is so because the value is higher than the diversity values for other woodland reserves in East Africa such as the ones for Maruzi Hill (Kiyingi et al., 2010), Miombo Forest Reserves (Gonçalves et al., 2017; Jew et al., 2016), Ruvu South Forest Reserve (Mligo, 2018), Nongeni Forest Reserve (Kacholi, 2019), Taita Hills (Wekesa et al., 2019) and Kitui woodlands (Kiruki et al., 2017). It is also still higher and compares well above with the diversity results from studies in other East African Reserve such as Mabira (Weldemariam et al. 2017) and Southern Rangelands Forests of Kenya (Jawuoro et al., 2017).
Since a reserve with a diversity value (H’) greater than two (H’ ≥ 2) can be regarded as highly diverse in terms of species (Demies et al., 2019; Ifo et al., 2016), it implies that Agoro-Agu Central Forest Reserve with a diversity of 3.40 harbours rich tree diversity and high species richness. The reason for higher diversity values in the present study may be due to the protected status and a wider range of geographic factors like altitude, aspect, and productive soil (Bhat et al., 2020). Subsequently, more conservation efforts are urgently needed to buffer such a reserve from further degradation and habitat loss.
According to Heywood (2019), conservation and protection efforts should be prioritised in areas experiencing a high rate of habitat loss such as Agoro-Agu Central Forest Reserve. This is expected to enhance the natural ecological restoration and regeneration of species at risks of local extinction in the degraded part of the reserve. One such effort may involve establishment of buffer zones in order to help conserve species richness and diversity; and also to regulate the utilization of tree resources by local dwellers (Siraj et al., 2018). There is also a need to enforce the provision of environmental awareness in the local communities on the importance of forests and build-up a “we feel” for the communities, which will actually promote responsible management, protection, utilization and conservation of such species (Kacholi, 2019).
Tree species evenness
The general tree species evenness as a measure of equitability of species distribution in Agoro-Agu CFR is 0.80. This is a bit higher than that of the degraded part (0.52). This implies that up to 80% of the tree species are fairly and evenly distributed in the intact part of the reserve; an indication that every encountered species was distributed evenly within the plant community in the intact part of the forest (Mekonen et al., 2015). According to Ifo et al. (2016), such high a value of species evenness can be attributed to less competition for space among tree species in intact part of the reserve.
Along topopositions, maximum value of species evenness of 0.70 is witnessed in the mid-slope and least (0.50) in the ridge top (Table 6). This is probably caused by favourable environmental conditions in the mid slope as oppose to harsher environmental conditions (shallower soils, strong winds, low moisture) that usually characterize the ridge top. The existence of such extreme conditions is likely to affect the distribution of species in such a ridge top with limited space due to rock outcrops (Moradi et al., 2020). However, given the recent rise in population for returnees/influx of South Sudan refugees and the heavy dependence of people on the forests for food, fuel and construction materials, future planned development should consider how additional human pressure could impact long-term sustainability of the region.
On the other hand, the fact that parts of the reserve largely left intact during LRA and SPLA wars had high species diversity is encouraging. Accordingly these findings should be used to boost the creation of a National Park to protect key vegetation types and their associated biodiversity in Agoro-Agu and neighbouring forest reserves (Corrigan et al., 2018). Finally, by the refugees leaving the encroach part of the reserves and getting settled outside Agoro-Agu Central Forest Reserve, is a positive sign that a reduction in population pressure can allow for the recovery of forests in this area (Watson et al., 2018); potentially paving the way for a trans-boundary park with related conservation benefits (Trogisch and Fletcher, 2020; Pouya and Pouya, 2018) between Uganda and South Sudan.
In some cases, natural regeneration is associated with rural outmigration (returnees) or remittance economies. Since reestablishing native forest cover does not have to require mass exodus of families and decline of rural livelihoods or traditions, new ways of thinking about how natural regeneration (coupled with other solutions), may promote a rural resurgence where communities and local economies thrive along with expansion of native forests should be taken up by concerned authorities. It should, however be noted that, one challenge for policy initiatives that promote natural regeneration is to address the social costs and drivers of rural outmigration. This is so, because, enhancing natural regeneration of native forests is not a viable option for forest restoration if these changes fail to provide benefits for rural residents and forests are short-lived (Chazdon and Brancalion, 2019).
Subsequently, in the new era of restoration, rural livelihoods should be re-envisioned through new opportunities created by growing native forests and trees in agricultural landscapes. For example, community based rural development projects should be implemented with the support of rural organizations and decision makers at local and regional levels (Hissa et al., 2019). This can make rural communities become stewards of community-managed forests that provide local, regional and global benefits. When done, this can promote rural resurgence based on proactive and integrated land management and landscape-scale restoration, where forests and new generations of people have room to grow and prosper together (Chazdon et al., 2020).
The outlined research issues emphasize that understanding the relationship between armed conflicts and land-change will require interdisciplinary research efforts, bringing together specialists from forestry, agricultural, geographical and environmental sciences; in addition to political sciences, and anthropology. It is unlikely that any research discipline alone will be able to uncover and delineate the causal linkages between armed conflict and biodiversity conservation. This implies that working together should be beneficial to improving our understanding of the potential for scientific and conservation engagement to mitigate negative biodiversity impacts in war zones, and the importance of the postwar period for incorporating biodiversity priorities into reconstruction and recovery efforts (Hanson, 2018).
The tree composition consisting of mixed species with indicator species for both savannah woodlands (C. molle, G. mollis, A. grandibracteata and T. glaucescens) and Afromontane/medium altitude tree species (Podocarpus spp., L. trichilioides (African Walnut) and L. swynnertonii) is an indication that Agoro-Agu Central Forest Reserve is an enriched forest reserve with a mixture of medium altitude Afromontane and savannah woodland trees.
The absence of trees species such as Podocarpus spp., Strombosia scheffleri, L. swynnertonii, M. bagshawei, P. fulva, P. mildbraedii, Hagenia abyssinica, A. africana, and L. trichiloides, S. longipedunculata is an sign of low regeneration potential of such species in the intact part as oppose to the degraded part of the reserve. This could probably be attributed to either an unidentified threats or loss of adult seeding trees and change in species representation which after anthropogenic disturbance is dominated by pioneer species.
The presence of locally threatened species like Podocarpus spp. and S. scheffleri; and internationally threatened species like L. swynnertonii and L. trichilioides display the uniqueness of Agoro-Agu Central Forest Reserve and its potential as biodiversity hotspot for conservation.
The development and changes of floristic composition and diversity of plant seedlings and saplings during early stages of secondary succession process could have been mostly influenced by secondary succession processes and fallow period in various ages of secondary forests in the intact and degraded parts respectively. Such information on the composition and diversity of different species are useful for predicting future trends in the vegetation succession, especially on secondary succession of degraded parts; and ought to be ascertained.
Understanding the composition and diversity of plant regeneration at early stages of secondary succession on the degraded parts also generates useful information for enhancing biodiversity conservation, and social and economic values for future forest.
The total absence of P. latifolia, S. scheffleri, L. swynnertonii, M. bagshawei, P. fulva, P. mildbraedii and H. abyssinica from the degraded part of the reserve raises issues of conservation concern and a need for long term monitoring or research on the life-cycle, propagation (both sexual and asexual) methods and regeneration potentials of these woody species in order to understand their population dynamics.
Considerable management options that limits continuous degradations such as enforcement of environmental laws, tree enrichment planting and ex-situ conservation should be undertaken to restore particularly the status of trees in the degraded part of the reserve.
Agoro-Agu Central Forest Reserve with a diversity of 3.40 harbours rich tree diversity and high species richness warranting more conservation efforts aimed at buffering such a reserve from further degradation and habitat loss. Ideally, conservation and protection efforts should be prioritised in areas experiencing a high rate of habitat loss in order to augment the natural ecological restoration and regeneration of species at risks of local extinction in the degraded part of the reserve. One of such efforts may involve establishment of buffer zones in order to help conserve species richness and diversity; and also to regulate the utilization of tree resources by surrounding local dwellers.
Agoro-Agu forest landscape is huge with several forest tenures including central forest reserves, community forests, local forest reserves that should be conserved and sustainably managed with an immense potential for scale up of Community Forest Management (CFM) within its landscape.
The authors declare that there has been no conflicting/ competing interest, financial or personal relationships in relation to this manuscript that may have inappropriately influenced the writing and publication of this article.
This work was funded by USAID through the Wildlife Conservation Society. The authors wish to recognize the logistics and technical support by the Project Manager (Beatrice Kyasimire) at the Wildlife Conservation Society. The author is grateful to the communities of Agoro-Agu who willingly participated in the interviews and the National Forestry Authority for granting permission that enable the study to be conducted in this forest reserve.
REFERENCES
|
Abiem I, Arellano G, Kenfack D, Chapman H (2020). Afromontane forest diversity and the role of grassland-forest transition in tree species distribution. Diversity 12(1):30.
Crossref
|
|
|
|
Arroyo?Rodríguez V, Melo FP, Martínez?Ramos M, Bongers F, Chazdon RL, Meave JA, Tabarelli M (2017). Multiple successional pathways in human?modified tropical landscapes: new insights from forest succession, forest fragmentation and landscape ecology research. Biological Reviews 92(1):326-340.
Crossref
|
|
|
|
|
Asigbaase M, Sjogersten S, Lomax BH, Dawoe E (2019). Tree diversity and its ecological importance value in organic and conventional cocoa agroforests in Ghana. PloS One 14(1):e0210557.
Crossref
|
|
|
|
|
Aynekulu E, Aerts R, Denich M, Negussie A, Friis I, Demissew S, Boehmer HJ (2016). Plant diversity and regeneration in a disturbed isolated dry Afromontane forest in northern Ethiopia. Folia Geobotanica 51(2):115-127.
Crossref
|
|
|
|
|
Bhat JA, Kumar M, Negi AK, Todaria NP, Malik ZA, Pala NA, Kumar A, Shukla G (2020). Species diversity of woody vegetation along altitudinal gradient of the Western Himalayas. Global Ecology and Conservation 24:e01302.
Crossref
|
|
|
|
|
Birhanu L, Bekele T, Tesfaw B, Demissew S (2021). Relationships between topographic factors, soil and plant communities in a dry Afromontane forest patches of Northwestern Ethiopia. PLoS One 16(3):e0247966.
Crossref
|
|
|
|
|
Chazdon R, Brancalion P (2019). Restoring forests as a means to many ends. Science 365(6448):24-25.
Crossref
|
|
|
|
|
Chazdon RL, Lindenmayer D, Guariguata MR, Crouzeilles R, Benayas JM, Chavero EL (2020). Fostering natural forest regeneration on former agricultural land through economic and policy interventions. Environmental Research Letters 15(4):043002.
Crossref
|
|
|
|
|
Chen W, Zhong J, Sun S, Xie Z, Zhou Y (2017). Fuelwood collection depresses the seed-dispersal service provided by rodents. Forest Ecology and Management 406:53-60.
Crossref
|
|
|
|
|
Corrigan C, Bingham H, Shi Y, Lewis E, Chauvenet A, Kingston N (2018). Quantifying the contribution to biodiversity conservation of protected areas governed by indigenous peoples and local communities. Biological Conservation 227:403-412.
Crossref
|
|
|
|
|
Dangulla M, Abd Manaf L, Ramli MF, Yacob MR (2020). Urban tree composition, diversity and structural characteristics in North-western Nigeria. Urban Forestry and Urban Greening 48:126512.
Crossref
|
|
|
|
|
Delgado-Aguilar MJ, Fassnacht FE, Peralvo M, Gross CP, Schmitt CB (2017). Potential of TerraSAR-X and Sentinel 1 imagery to map deforested areas and derive degradation status in complex rain forests of Ecuador. International Forestry Review 19(1):102-118. Demie G (2019). Woody Species Diversity and Composition of Dry Woodland Vegetation in West Shewa, Central Ethiopia: Implications for Their Sustainable Management. American Journal of Agriculture and Forestry 7(6):282-289.
Crossref
|
|
|
|
|
Demies M, Samejima H, Sayok AK, Noweg GT (2019). Tree diversity, forest structure and species composition in a logged-over mixed dipterocarp forest, Bintulu, Sarawak, Malaysia. Science and Technology 6(1-2):102.
|
|
|
|
|
Deo GR, Banoo S, Tehmeena M, Suniti D, Ankit K, Vipin V (2016). Diversity of benthic macro-invertebrates in four tributaries of River Narmada in the central zone, India. International Journal of Life Sciences 4(1):107-115.
|
|
|
|
|
Dibaba A, Soromessa T, Kefalew A, Addi A (2020). Woody Species Diversity, Vegetation Structure, and Regeneration Status of the Moist Afromontane Forest of Agama in Southwestern Ethiopia. International Journal of Ecology 2020:10.
Crossref
|
|
|
|
|
Environmental Alert (2017). Integrating Pro-Poor and Human Rights-Based Approaches in Collaborative Forest Management Processes a Case study of Agoro-Agu Forest Landscape CFM Process, November 2017. 18 p.
|
|
|
|
|
|
|
Fazlollahi MM, Jalali SG, Kooch Y, Theodose TA (2017). Tree species composition, biodiversity and regeneration in response to catena shape and position in a mountain forest. Scandinavian Journal of Forest Research 32(1):80-90.
Crossref
|
|
|
|
|
Gadow KV, Zhang G, Durrheim G, Drew D, Seydack A (2016). Diversity and production in an Afromontane Forest. Forest Ecosystems 3(1):1-12.
Crossref
|
|
|
|
|
Gavin MC, McCarter J, Berkes F, Mead ATP, Sterling EJ, Tang R, Turner NJ (2018). Effective biodiversity conservation requires dynamic, pluralistic, partnership-based approaches. Sustainability 10(6):1846.
Crossref
|
|
|
|
|
Gebeyehu G, Soromessa T, Bekele T, Teketay D (2019). Species composition, stand structure, and regeneration status of tree species in dry Afromontane forests of Awi Zone, northwestern Ethiopia. Ecosystem Health and Sustainability 5(1):199-215.
Crossref
|
|
|
|
|
Gonçalves FM, Revermann R, Gomes AL, Aidar MP, Finckh M, Juergens N (2017). Tree species diversity and composition of Miombo woodlands in South-Central Angola: A chronosequence of forest recovery after shifting cultivation. International Journal of Forestry Research, Vol. 2017. Article ID 6202093: 13.
Crossref
|
|
|
|
|
Gorsevski V, Geores M, Kasischke E (2013). Human dimensions of land use and land cover change related to civil unrest in the Imatong Mountains of South Sudan. Applied Geography 38:64-75.
Crossref
|
|
|
|
|
Gorsevski V, Kasischke E, Dempewolf J, Loboda T, Grossmann F (2012). Analysis of the impacts of armed conflict on the Eastern Afromontane forest region on the South Sudan-Uganda border using multitemporal Landsat imagery. Remote Sensing of Environment 118:10-20.
Crossref
|
|
|
|
|
Hanson T (2018). Biodiversity conservation and armed conflict: a warfare ecology perspective. Annals of the New York Academy of Sciences 1429(1):50-65.
Crossref
|
|
|
|
|
Haruna H, Aliko AA, Zakari SM, Omeiza AH (2018). Quantitative analysis of plant species diversity in Kano zoological garden. Bayero Journal of Pure and Applied Sciences 11(1):208-213.
Crossref
|
|
|
|
|
Hegazy AK, Hosni HA, Lovett-Doust L, Kabiel HF, Badawi ES, Mwavu EN (2020). Indigenous knowledge of wild plants collected in Darfur, Sudan. Ethnobotany Research and Applications 19:1-19.
Crossref
|
|
|
|
|
Heywood VH (2019). Conserving plants within and beyond protected areas-still problematic and future uncertain. Plant Diversity 41(2):36-49.
Crossref
|
|
|
|
|
Hissa HR, Alves Filho NT, Costa M, Strauch G, Bassi L, de Assis RL (2019). Sustainable rural development in Rio de Janeiro state: the Rio rural program. In Strategies and Tools for a Sustainable Rural Rio de Janeiro. Springer, Cham. pp. 23-39.
Crossref
|
|
|
|
|
Ifo SA, Moutsambote JM, Koubouana F, Yoka J, Ndzai SF, Bouetou-Kadilamio LN, Mampouya H, Jourdain C, Bocko Y, Mantota AB, Mbemba M (2016). Tree species diversity, richness, and similarity in intact and degraded forest in the tropical rainforest of the Congo Basin: case of the forest of Likouala in the Republic of Congo. International Journal of Forestry Research Article ID 7593681:12.
Crossref
|
|
|
|
|
International Union for Conservation of Nature (IUCN) (2015). Socio Economic Baseline Report for Agoro-Agu forest landscape. Available options for REDD+ implementation in Uganda. Kampala-Uganda.
|
|
|
|
|
IUCN (2020) The IUCN Red List of Threatened Species. Version 2020-1.
|
|
|
|
|
Jawuoro SO, Koech OK, Karuku GN, Mbau JS (2017). Plant species composition and diversity depending on piospheres and seasonality in the southern rangelands of Kenya. Ecological Processes 6(1):1-9.
Crossref
|
|
|
|
|
Jenerette GD, Clarke LW, Avolio ML, Pataki DE, Gillespie TW, Pincetl S, Nowak DJ, Hutyra LR, McHale M, McFadden JP, Alonzo M (2016). Climate tolerances and trait choices shape continental patterns of urban tree biodiversity. Global Ecology and Biogeography 25(11):1367-1376.
Crossref
|
|
|
|
|
Joshi RK, Dhyani S (2019). Biomass, carbon density and diversity of tree species in tropical dry deciduous forests in Central India. Acta Ecologica Sinica 39(4):289-299.
Crossref
|
|
|
|
|
Kacholi DS (2019). Assessment of Tree Species Richness, Diversity, Population Structure and Natural Regeneration in Nongeni Forest Reserve in Morogoro Region, Tanzania. Tanzania Journal of Science 45(3):330-345.
|
|
|
|
|
Kalema J, Hamilton A (2020). Field guide to the forest trees of Uganda: for identification and conservation. CAB International, UK.
|
|
|
|
|
Kidane YO, Steinbauer M.J, Beierkuhnlein C (2019). Dead end for endemic plant species? A biodiversity hotspot under pressure. Global Ecology and Conservation 19: e00670.
Crossref
|
|
|
|
|
Kiruki HM, van der Zanden EH, Gikuma-Njuru P, Verburg PH (2017). The effect of charcoal production and other land uses on diversity, structure and regeneration of woodlands in a semi-arid area in Kenya. Forest Ecology and Management 391:282-295.
Crossref
|
|
|
|
|
Kiyingi I, Kidiya JM, Gwali S, Okullo P, Byabashaija DM (2010). Tree species composition, structure and utilisation in Maruzi Hills Forest Reserve in Uganda. Southern Forests 72(2):113-117.
Crossref
|
|
|
|
|
Mairal M, Sanmartín I, Herrero A, Pokorny L, Vargas P, Aldasoro JJ, Alarcón M (2017). Geographic barriers and Pleistocene climate change shaped patterns of genetic variation in the Eastern Afromontane biodiversity hotspot. Scientific Reports 7(1):1-13.
Crossref
|
|
|
|
|
Martínez?Ramos M, Pingarroni A, Rodríguez?Velázquez J, Toledo?Chelala L, Zermeño?Hernández I, Bongers F (2016). Natural forest regeneration and ecological restoration in human?modified tropical landscapes. Biotropica 48(6):745-757.
Crossref
|
|
|
|
|
Mengich EK, Macharia JM, Mitloehner R, Too DK, Muturi GM (2020). Diameter distribution of indigenous trees as indicator of adapted species in semi-arid rangelands of Kenya. Journal of Tropical Forest Science 32(2):144-153.
Crossref
|
|
|
|
|
Migliore J, Lézine AM, Hardy O (2020). The recent colonization history of the most widespread Podocarpus tree species in Afromontane forests. Annals of Botany Oxford University Press (OUP) 126(1):73-83.
Crossref
|
|
|
|
|
Ministry of Water and Environment (2017). Forest Investment Program for Uganda. Ministry of Water and Environment. Government of the Republic of Uganda, Kampala, Uganda.
|
|
|
|
|
Mligo C (2018). The plant species composition, diversity and natural regeneration of indigenous trees in the disturbed Ruvu South Forest Reserve, Tanzania. Tanzania Journal of Science 44(3):46-60.
|
|
|
|
|
Moradi H, Fattorini S, Oldeland J (2020). Influence of elevation on the species-area relationship. Journal of Biogeography 47(9):2029-2041.
Crossref
|
|
|
|
|
Mori AS, Lertzman KP, Gustafsson L (2017). Biodiversity and ecosystem services in forest ecosystems: a research agenda for applied forest ecology. Journal of Applied Ecology 54(1):12-27.
Crossref
|
|
|
|
|
Ministry of Wildlife, Tourism and Antiquities, MTWA (2018). Red List of Threatened Species of Uganda, 2018. Ministry of Wildlife, Tourism and Antiquities (MTWA) Kampala, Uganda.
|
|
|
|
|
Muluneh MG, Feyissa MT, Wolde TM (2021). Effect of forest fragmentation and disturbance on diversity and structure of woody species in dry Afromontane forests of northern Ethiopia. Biodiversity and Conservation 30(6):1753-1779.
Crossref
|
|
|
|
|
Nabasumba D, Eilu G, Bahati J, Kamwesigye G (2016). Utilisation and conservation status of indigenous woody plant species in a sedentary pastoral production system in south-western Uganda. Uganda Journal of Agricultural Sciences 17(2):125-137.
Crossref
|
|
|
|
|
Newmark WD, McNeally PB (2018). Impact of habitat fragmentation on the spatial structure of the Eastern Arc forests in East Africa: Implications for biodiversity conservation. Biodiversity and Conservation 27(6):1387-1402.
Crossref
|
|
|
|
|
NFA (2018). National Forestry Authority. Revised Forest Management Plan for Agoro-Agu Sector Central Forest Reserves for the Period: 1st July 2018-30th June 2028; National Forestry Authority: Kampala, Uganda, 2018.
|
|
|
|
|
Olanya BP (2020a). Diversity, Population Structure and Regeneration Status of Indigeous Woody Species in and Around Agoro-Agu Central Forest Reserve, Uganda. Journal of Ecosystem and Ecography 10:266.
|
|
|
|
|
Olanya BP (2020b). Environmental Determinants of Woody Species Distribution Patterns in Agoro Agu Central Forest Reserve, Northern Uganda. Journal of Ecosystem and Ecography 10(1-8):268.
|
|
|
|
|
Omoding J, Walters G, Andama E, Carvalho S, Colomer J, Cracco M, Eilu G, Kiyingi G, Kumar C, Langoya CD, Nakangu Bugembe B (2020). Analysing and Applying Stakeholder Perceptions to Improve Protected Area Governance in Ugandan Conservation Landscapes. Land 9(6):207.
Crossref
|
|
|
|
|
Pouya S, Pouya S (2018). Planning for Peace; Introduction of Transboundary Conservation Areas. Kocaeli Journal of Science and Engineering 1(2):33-41.
Crossref
|
|
|
|
|
Rahman IU, Hart R, Afzal A, Iqbal Z, Ijaz F, Abd_Allah EF, Ali N, Khan SM, Alqarawi AA, Alsubeie MS, Bussmann RW (2019). A new ethnobiological similarity index for the evaluation of novel use reports. Applied Ecology and Environmental Research 17(2):2765-2777.
Crossref
|
|
|
|
|
Ranjan R (2018). What drives forest degradation in the central Himalayas? Understanding the feedback dynamics between participatory forest management institutions and the species composition of forests. Forest Policy and Economics 95:85-101.
Crossref
|
|
|
|
|
Reshad M, Muhammed M, Beyene A (2020). Distribution and Importance Value Index of Woody Species Under Different Successional Stages at Jello-Muktar Dry Afromontane Forest, South-eastern Ethiopia. American Journal of Agriculture and Forestry 8(1):1-8.
Crossref
|
|
|
|
|
Salazar A, Sanchez A, Villegas JC, Salazar JF, Ruiz Carrascal D, Sitch S, Restrepo JD, Poveda G, Feeley KJ, Mercado LM, Arias PA (2018). The ecology of peace: preparing Colombia for new political and planetary climates. Frontiers in Ecology and the Environment 16(9):525-531.
Crossref
|
|
|
|
|
Santini L, Belmaker J, Costello MJ, Pereira HM, Rossberg AG, Schipper AM, Ceau?u S, Dornelas M, Hilbers JP, Hortal J, Huijbregts MA (2017). Assessing the suitability of diversity metrics to detect biodiversity change. Biological Conservation 213:341-350.
Crossref
|
|
|
|
|
Shirima DD, Totland Ø, Moe SR (2016). The relative importance of vertical soil nutrient heterogeneity, and mean and depth-specific soil nutrient availabilities for tree species richness in tropical forests and woodlands. Oecologia 182(3):877-888.
Crossref
|
|
|
|
|
Siraj M, Zhang K (2018). Structure and natural regeneration of woody species at central highlands of Ethiopia. Journal of Ecology and the Natural Environment 10(7):147-158.
Crossref
|
|
|
|
|
Siraj M, Zhang K, Xiao W, Bilal A, Gemechu S, Geda K, Yonas T, Xiaodan L (2018). Does participatory forest management save the remnant forest in Ethiopia? Proceedings of the National Academy of Sciences, India Section B: Biological Sciences 88(1):1-14.
Crossref
|
|
|
|
|
Strassburg BB, Beyer HL, Crouzeilles R, Iribarrem A, Barros F, de Siqueira MF, Sánchez-Tapia A, Balmford A, Sansevero JB, Brancalion PH, Broadbent EN (2019). Strategic approaches to restoring ecosystems can triple conservation gains and halve costs. Nature Ecology and Evolution 3(1):62-70.
Crossref
|
|
|
|
|
Swinfield T, Afriandi R, Antoni F, Harrison RD (2016). Accelerating tropical forest restoration through the selective removal of pioneer species. Forest Ecology and Management 381:209-216.
Crossref
|
|
|
|
|
Tecimen HB, Sevgi O, Akkaya M, Sevgi E, Hancer ÇK, Cakir EA (2017). Comparison of species richness and diversity at natural stands and plantations of stone pine (Pinus pinea L.). Pakistan Journal of Botany 49(5):1743-1748.
|
|
|
|
|
Tenzin J, Hasenauer H (2016). Tree species composition and diversity in relation to anthropogenic disturbances in broad-leaved forests of Bhutan. International Journal of Biodiversity Science, Ecosystem Services & Management 12(4):274-290.
Crossref
|
|
|
|
|
Tolangara A, Ahmad H, Liline S (2019). The Composition and Important Value Index of Trees for Wildlife Feed in Bacan Island, South Halmahera. In IOP Conference Series: Earth and Environmental Science 276(1):012037).
Crossref
|
|
|
|
|
Trogisch L, Fletcher R (2020). Fortress tourism: exploring dynamics of tourism, security and peace around the Virunga trans boundary conservation area. Journal of Sustainable Tourism 8:1-21.
Crossref
|
|
|
|
|
Uganda Land Use Dialogue (2020).
View 24th May 2020.21.25pm.
|
|
|
|
|
UNEP-WCMC, IUCN (2020). Protected Planet. The World Database on Protected Areas (WDPA); UNEP-WCMC and IUCN: Cambridge, UK, 2020.
|
|
|
|
|
Watson JE, Evans T, Venter O, Williams B, Tulloch A, Stewart C, Thompson I, Ray JC, Murray K, Salazar A, McAlpine C (2018). The exceptional value of intact forest ecosystems. Nature Ecology and Evolution 2(4):599-610.
Crossref
|
|
|
|
|
Wegasie MO, Eldegard K, Klanderud K, Tewolde-Berhan S, Giday K, Totland Ø (2018). Locally endangered tree species in a dry montane forest are enhanced by high woody species richness but affected by human disturbance. Journal of Arid Environments 158:19-27.
Crossref
|
|
|
|
|
Wekesa C, Kirui BK, Maranga EK, Muturi GM (2019). Variations in forest structure, tree species diversity and above-ground biomass in edges to interior cores of fragmented forest patches of Taita Hills, Kenya. Forest Ecology and Management 440:48-60.
Crossref
|
|
|
|
|
Weldemariam EC, Jakisa ES, Ahebwe DA (2017). Implication of forest zonation on tree species composition, diversity and structure in Mabira Forest, Uganda. Environment, Earth and Ecology 1(1).
Crossref
|
|
|
|
|
Yahya N, Gebre B, Tesfaye G (2019). Species diversity, population structure and regeneration status of woody species on Yerer Mountain Forest, Central Highlands of Ethiopia. Tropical Plant Research 6(2):206-213.
Crossref
|
|
|
|
|
Young NE, Romme WH, Evangelista PH, Mengistu T, Worede A (2017). Variation in population structure and dynamics of montane forest tree species in Ethiopia guide priorities for conservation and research. Biotropica 49(3):309-317.
Crossref
|
|
|
|
|
Yuan ZY, Jiao F, Li YH, Kallenbach RL (2016). Anthropogenic disturbances are key to maintaining the biodiversity of grasslands. Scientific Reports 6(1):1-8.
Crossref
|
|
|
|
|
Zhao Y, Feng D, Jayaraman D, Belay D, Sebrala H, Ngugi J, Maina E, Akombo R, Otuoma J, Mutyaba J, Kissa S (2018). Bamboo mapping of Ethiopia, Kenya and Uganda for the year 2016 using multi-temporal Landsat imagery. International Journal of Applied Earth Observation and Geoinformation 66:116-125.
Crossref
|
|