ABSTRACT
The study was aimed at contributing to sustainable management of Ajei Highlands Watershed Community Forest, one of the vulnerable and highly threatened biological hotspots remaining within the Cameroon western highlands through an analysis of its woody species diversity and stand structure. The results of inventory of its 12 gallery forest patches through 17 permanent sample plots (20 m x 50 m) with multiple spatial scales subplots (1, 10 and 100 m2) using Whittaker plot sampling method showed that a total of 3644 individuals were recorded (124 species showing exclusive affinity to forest patches, 90 genera and 46 families). Diversity indices were 4.34 and 0.93 for Shannon-Weaver and Simpson indexes, respectively, indicating a highly diverse forest. The mean density was 1820 ± 24 stems/ha and the basal area 27.72 ± 0.04 m2/ha; with mean and maximum diameter of 9.3 and 140 cm, respectively. Species importance value index ranged from 0.35 to 35.19%, the most dominant species being Macaranga occidentalis, Xymalos monospora and Rauvolfia vomitoria. Euphorbiaceae, Apocynaceae, Moraceae and Rubiaceae were the most important families. Although this forest presented a high diversity and a certain floristic richness; few potentially commercial timber species were identified. Management options are prescribed within the framework of a participative management plan promoting watershed protection and habitat restoration, targeting livelihoods sustenance through agroforestry and reforestation activities favoring local tree species.
Key words: Woody diversity; watershed, gallery forest, community forest, Cameroon.
More than 1.6 billion people depend on varying degrees on forests for their livelihood (SCBD, 2009) and forests play a key role in the economic development of many countries. According to FAO (2015), global forest cover has drastically decreased from 4128 million hectare in 1990 to 3999 million hectare in 2015, corresponding to a drop from 31.1 to 30.6% (1.5% loss). The fragmentation and loss of tropical forest through deforestation and land degradation is the greatest threat to ecological services provided by these forests, such as biodiversity conservation (Hansen et al., 2020). This fragmentation process has resulted in increased riparian forest formation along watercourses across tropical forest highland regions serving as watersheds; and is considered as global biological hotspots, which have supported local populations with the provision of timber and non-timber forests products (Natta, 2003). The rapid change in land use for local subsistence in the Western highland region of Cameroon has resulted to an unprecedented destruction and fragmentation of riparian areas of gallery forests resulting in a few scattered forest patches on the landscape (Momo Solefack et al., 2018). These highland regions are rich in endemic plants and animal species (Momo Solefack et al., 2016; Wouokoue et al., 2017). The sub-montane and afromontane forests of Cameroon are sources of the most important fresh water streams in the country, particularaly in the western highlands, Noun, Benoue, Djerem, Lom, and Sanaga rivers for the Adamawa Massif (Lambi, 2001). Referring to high value conservation forests, gallery forests are classified as endangered ecosystems because they are marginal among the wooded vegetation and also because of their high degradation status.
Gallery forests in tropical savanna landscapes are mostly narrow strips of forest associated with creeks and rivers, in an otherwise unforested landscape (Veneklaas et al., 2005). Gallery forests in savannas represent one of the few examples of naturally fragmented tropical forests. They are generally rich in woody plant species typical of continuous forests and are postulated to have provided refugee for tropical forest species in areas deforested during the Pleistocene drought (Meave and Kellman, 1994). This forest type is important to the streams/river course; and also the biodiversity it harbours providing fertile moist soils, wood, water, shelter and non-timber forest products used by the populations to meet their needs (Ajonina and Tomedi, 2014).
Despite their recognized status, biodiversity potentials and their ecosystem services, gallery forests have remained unmanaged and ignored as key ecosystems in biodiversity conservation. This reason could be attributed to insufficient scientific knowledge on their potential to support local populations, and their contribution to climate change mitigation; thus, hindering their consideration in global conservation programmes. To combat these challenges, conservation options like community forestry encouraged the decentralisation and management of forest portions by the local population in order to support community development and biodiversity conservation. In several countries of the world, community forestry resulted in more sustainable management of forests when communities found it in their interests to conserve them (Maryudi et al., 2012; Kimengsi et al., 2019).
In a bid to redress threats of forest cover loss and promote community participation and alleviate poverty, the Cameroonian Forestry, Wildlife and Fisheries law, enacted in 1994, enshrined the concept of community forest, corresponding to a maximum area of 5000 ha, granting local community’s access, to use and manage substantial portion of the non-permanent forest estate (MINFOF, 2009). The Ajei community forest in North West Region of Cameroon was established in January 2014. This community forest is one of the remaining forest patches in the lower ridge of the Bamenda highland region and serves as an important watershed for the streams that flow into the River Momo feeding the Mamfe-Cross River Basins (Ndip et al., 2018); and also providing drinking water for the surrounding communities (AHWaCoFoMB, 2016). However, the area is under threats with challenging human activities and natural phenomena, including: Shifting cultivation, slash and burn agriculture, settlement, bush fires, and encroachment by grazers and farmers (Temgoua et al., 2018). Temgoua et al. reported an ongoing deforestation with forest cover loss of about 240 ha in 30 years (1988-2018). Understanding tree species composition, diversity, and structure is a vital source of information in assessing sustainability of any forest, conservation of species, and management of the ecosystems at large (Ssegawa and Nkuutu, 2006; Addo-Fordjour et al., 2009). However, no study has yet been carried out in this forest to assess its floristic composition and diversity. This paper aims to contribute to the sustainable management of the Ajei highland watershed community forest by providing a sound scientific basis that maintains its ecological integrity through the analysis of its floristic woody diversity and stand structure.
Study site
The study was carried out in the Ajei upland watershed community forest, located in Ngie subdivision, Momo division in the North West Region of Cameroon (Figure 1). This forest is located between latitudes 5.920° and 5.970°N and longitudes 9.840° and 9.890°E and covers a surface area of 1739.3 ha. The study site falls in the transitional zone between the wet tropical or equatorial climate in south Cameroon and the Sudanese climate of North Cameroon. Annual precipitation ranges from 2200 to 3000 mm from March through October with an average of 240 rainy days and an average temperature of 21oC (minimum 15 and maximum 30°C). The topography and relief ranges between 1500 m above sea level, cumulating at the highest peak of Ndek Akorbiri (2040 m) being typical of an area underlain by granitic basement rocks. Soils derived from this parent rock material are generally poor, being rapidly depleted of fertility through cropping and the erosive action of rain (AHWaCoFoMB, 2016). The vegetation is montane forests with interconnecting savanna habitats. Within the savanna, gallery forests occur as discontinuous strips of closed canopy woodland along the main streams. The Ajei upland watershed community forest is made up of 12 forest patches, some of the last remaining forest patches of the Bamenda Highlands. Plant species are typical assemblages of this montane landscape (AHWaCoFoMB, 2016).
Data collection
Data collection consisted of an inventory using a network of nested sampling units (plots, interplots, subplots and quadrats) in a transect arrangement following the Whittaker Plot Sampling Method (Shmida, 1984). The Whittaker plot sampling method was elaborated, adapted and implemented at multiple spatial scales (1 and 10 m2) on 1000 m2 permanent sample plots (Figure 2).
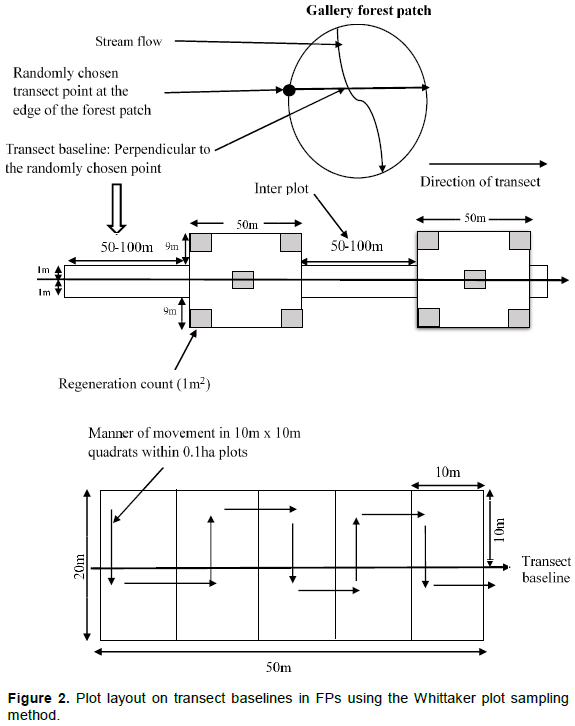
Plots were laid in all the 12 forest patches of the community forest spaced within a 2-m wide belt transect running perpendicular with the use of a compass from a randomly chosen GPS point close to the centre along one edge of forest patch crossing the stream to the other edge of the same forest patch. The transects were opened and pegged with polyvinyl chloride (PVC) plastic pipes in each forest patch also serving to demarcate the edges of the plots. Each transects and forest patch was identified with a signpost. At every 50- or 100-m interval (depending on the width of the patch) along the 2-m belt transect (inter-plot distance), permanent sample plots (PSPs) of dimension 20 m x 50 m (1000 m2) were systematically designed for the vegetation inventory (tree identification, measurement and tagging). The permanent sample plots were further sub-divided into ten 10 m x 10 m (100 m2) temporal subplots to ease data collection and access on the 1000 m2 plots (0.1 ha). On the inter plots (2 m × 50 m: 100 m2, or 2 m × 100 m: 200 m2) trees with dbh ≥ 10 cm in diameter were measured at 1.3 m (diameter at breast height); tagged and the point of measurement accurately marked out with red paint; while on plots (20 m × 50 m), diameter measurements were taken from trees with dbh ≥ 2.5 cm. At the edges and middle of each sample plot, 1 m × 1 m quadrats (1 m2) were established principally for regeneration count (seedlings and saplings dbh < 2.5 cm). At the end, 17 permanent sampling plots (each 20 m × 50 m) were established along 12 transects of varying length between 100 to 500 m within the 12 gallery forest patches that make up the community forest.
Each tree species sampled were noted from which the woody species checklist of Ajei community forest was established. Species that could not be identified while in the field were sampled, pressed and preserved for further identification in the herbarium of the Limbe Botanic Garden. The species were identified according to the Angiosperm Phylogeny Group (APG) IV (2016) classification.
Data analysis
Data analysis was performed with Microsoft Excel 2016 of Microsoft office and Palaeontology Statistics (PAST) software package version 2.
Floristic composition and diversity
Floristic characterization was done by determining parameters such as: Diversity indices, relative density and dominance of species and families. Species richness corresponds to the number of species in a community or stand (Magurran, 2004). The number of families and the number of species present in the community forest were determined, as well as the number of individuals of each species.
The Shannon-Weaver Diversity Index (H') is a measure of the potential for interaction between the species that make up a community. This index takes into account the number of species present and the distribution of individuals within those species.
H'= -∑ (ni/N) ln (ni/N)
where ni is the number of individuals of a given species i and N the total number of individuals. The Simpson's diversity index (D') is the probability that two randomly selected individuals are of different species. It is represented by the reciprocal of the Simpson's index (D). The maximum diversity is represented by the value 1 and the minimum diversity by the value 0.
D'=1 - ∑ (ni/N)2
The generic diversity index (IG) was calculated following the formula:
IG = δp/G
Where δp is the number of species and G the number of genera of the study population.
It helps to characterise the plant population stability (Sonke, 1998). A low value, close to 1 of this coefficient, indicates a high diversity of the flora (Ake Assi, 1984).
To describe the ecological importance of families and species within the total flora, the family importance value index (FIV; Mori et al., 1983) and the Importance value index (IVI); Curtis and McIntosh, 1950) were calculated using:
FIV = rDe + rDo + rDi
where rDe is the relative density (number of individuals of the family considered in relation to the total number of individuals × 100), rDo is the relative dominance (basal area of the family considered in relation to the total basal area of the stand × 100), and rDi is the relative diversity (number of species of the family considered in relation to the total number of species × 100).
IVI = rDe + rDo + rFr
Where rDe is the relative density (number of individuals of the species / the total number of individuals × 100), rDo is the relative basal area (basal area of the species / the total basal area of the stand × 100); and rFr is the relative frequency (frequency of a species / total of all frequencies × 100).
Stand structural parameters
Vegetation structure was determined by parameters such as density, basal area and distribution of individuals by diameter classes (size class distribution) commensurate with six distinct tree utility classes: <1 cm (seedlings); ≥1 cm to <2.5 cm (small saplings); ≥2.5 cm to <5 cm (medium sized saplings); ≥5 cm to <10 cm (small trees or posts); ≥10 cm to <30 cm (medium sized trees or poles); ≥30 cm to <50 cm (large trees or standards); and ≥50 cm (giant trees or veterans) (Barden et al., 1987).
Density and basal area were estimated using the formulas given by Kent and Coker (1992).
Density (D) is the number of individuals per hectare. It was calculated by converting the total number of individuals encountered in all plots to equivalent number per hectare, following this formula:
ð· = N/S
With D the density (stems ha-1), N is the number of stems present on the considered surface and S the area considered (ha).
Basal area (BA) provides information on the area occupied by tree sections at 1.30 m from the ground. The formula is:
BA=π/4 ∑ (Di²)
Where BA is basal area (m2 ha-1) and Di is diameter (m) for each measured section (i).
Principal coordinates analysis was performed with Palaeontology Statistics (PAST) software to explore and to visualize similarities or dissimilarities of different forest patches.
Diversity, floristic composition and richness
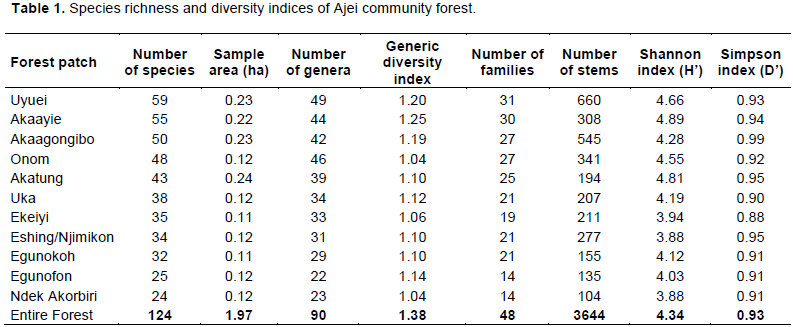
In the 1.97 ha surveyed, a total of 3644 individuals were recorded, belonging to 124 species, 90 genera and 46 families (Table 1). About 84.67% of species were identified to species level, 12.09% identified to generic level, 2 species identified to family level and 1.26% (2 species) remain unidentified. From the floristic inventory, 4 life forms were identified. This includes: Trees, shrubs, lianas and palms. Trees were the most abundant with 70 species (56.5%), followed by shrubs with 47 species (37.9%), lianas with 5 species (4.0%) and palms with 2 species (1.6%).
The Shannon-Weaver diversity index (H’) varied from 3.88 to 4.89 between different forest patches with a value of 4.34 for the entire forest, reflecting a high diversity of our study site. Simpson’s diversity index was 0.93.
Table 1 show that the number of species per forest patches varies from 24 to 62, the richest patch being Asoh and the poorest Ndek Akorbiri. Values of the Shannon diversity index were highest in Akaayie and Akatung forest patches (H’= 4.89 and 4.81, respectively) and lower in Eshing/Njimikon and Ndek Akorbiri (H’=3.88). Simpon’s diversity index was higher in Akaagongibo (0.99).
The relationship between the number of species and genus expressed by the generic diversity index was 1.32. This value shows an average of less than two species by genus. The number of genera having more than one species is few (13). The genus Psychotria was the most diverse with 9 species, followed by the genera Ficus (6 species) and Cola (3 species).
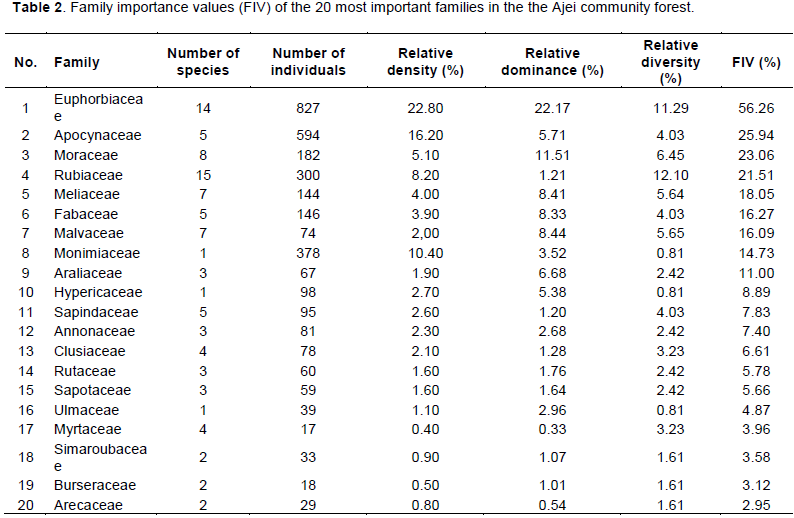
In terms of number of species per family, Rubiaceae was the best represented family with 15 species, followed by Euphorbiaceae (14), Moraceae (8), Meliaceae and Malvaceae (7), and Fabaceae and Sapindaceae with 5 species each. Families with one species were very common (32 families), while 8 families had between 2 and 4 species. The family importance values (FIV) of 20 most important families are presented in Table 2.
The five most important families were Euphorbiaceae (56.26%), Apocynaceae (25.94%), Moraceae (23.06%), Rubiaceae (21.51%), and Meliaceae (18.05%). 17 families had a FIV less than 3%.
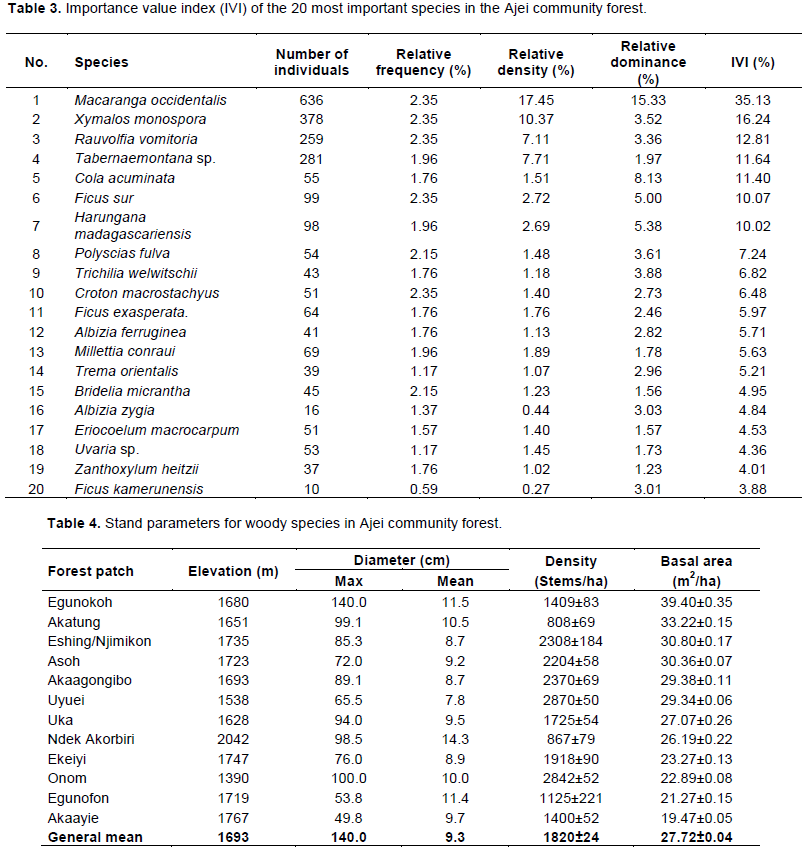
It appears from Table 3 that the species with higher importance value index were Macaranga occidentalis (35.13%), Xymalos monospora (16.24%), Rauvolfia vomitoria (12.81%), Tabernaemontana sp. (11.64%) and Cola acuminata (11.40%). These first five species represent about 41% of the individuals counted. A high number of species were represented by three or fewer individuals in the overall sample of the 12 patches taken together (40 species). The complete list of species is presented in Appendix Table 1.
Stand structural analysis
Density and basal area
Analysis based on all woody plants of at least 1 cm dbh showed the stem density of 1820 ± 24 ha-1 (Table 4). When considering only individuals with dbh ≥10 cm, the stem density was 693 stems ha-1. The smallest density (808 stems ha-1) was found in Akatung and the highest (2870 stems ha-1) in Onom. Basal area ranged from 19.4 m2 ha-1 (Akaayie) to 39.4 m2 ha-1 (Egunokoh) with a general mean of 27.72 ± 0.04 m2 ha-1 for the entire forest. Basal area depends not only on the trees’ density but also on the diameter of the trees. The mean diameter varied from 7.8 cm in Uyuei to 14.3 cm in Ndek Akorbiri with a general mean diameter of 9.3 cm for the entire forest. The maximium sized trees varied from 94 cm in Uka to 140 cm dbh in Egunokoh.
Diameter class abundance and distribution
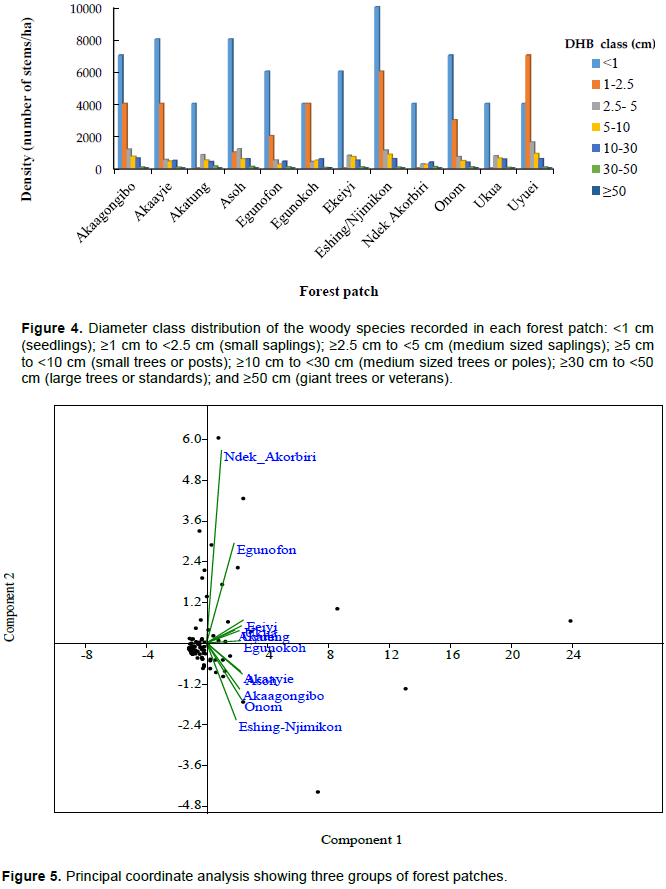
Concerning the diameter class abundance and distribution (Figure 3), the densities were highest for stems with diameter less than 1cm (6235 stems ha-1) (seedling category), followed by the diameter class 1 to 2.5cm (2941stems ha-1) (sapling category); meaning more regeneration. Woody vegetation has few trees with diameter larger than 50 cm (1 stem ha-1) (timber-sized trees). Stem densities were reduced with increase in diameter classes conforming to the characteristic inverted-J curve reflecting constant regeneration over time. Considering the diametric class distribution of each forest patch, the population size class frequency still exhibits a tendency towards an inverted-J curve distribution. In some forest patches (Uka, Ekeiyi, Ndek Akorbiri and Akatung) there is an absence of trees of diameter 1 to 2.5 cm; that is, sapling category (Figure 4). Only 2 forest patches (Onom and Egunokoh) showed the presence of trees of dbh ≥30 cm (reaching timber size).
The ordination by Principal Coordinate Analysis (Figure 5) indicated three groups of forest patches. Distances were expressed by similarity index of different forest patches in terms of species abundance, and showed a detectable gradient in the structure and floristic composition of the vegetation, reflecting variations in stem density and the abundance of certain species. The changes were interpretable in terms of topography and previous land use activities with Ndek Akorbiri (2040 m) and Egunofon (1811.18 m) distinctive at higher altitude. These forest patches have experienced little or no human intervention (particularly, the slash and burn agriculture practice), but are facing the consequences of bush fires from grazing activities, which annually consume the understorey situated behind only large old-growth trees, and few stems are left to succeed.
Diversity, floristic composition and richness
The Ajei community forest showed a floristic richness of 124 tree species, belonging to 90 genera and 48 families. In Cameroon, only a few known studies (Ndamason, 2016; Momo Soléfack et al., 2018) have been carried out to evaluate the vegetation composition of the gallery forests. For this reason, the results of this study cannot be compared with a wide range of other similar studies in Cameroon. The number of woody species found in this study is higher than that of 25 species found by Ndamason (2016) in Ijim Rigde gallery forest of Bamenda Highlands and also greater than that of 60 species found by Momo Soléfack et al. (2018) in the Koupa Matapit gallery forest. Fousseni et al. (2014) identified 61 tree species in a gallery forest in the sudanian savannah ecosystem of Togo. In the gallery forests found along
streams in the Mountain Pine Ridge Savannah of Belize, Kellman et al. (1998) found 66 tree species and in Muvumba gallery forest in Rwanda, ACNR (2011) identified 70 species. However, the number of trees species found in this study is very low compared to that found in riparian forests of Benin (224 species) by Natta (2003) and of Burkina Faso (196 species) by Sambaré et al. (2011).
The estimation of abundance and richness of plants can vary enormously depending upon whether inventorying included stems of small diameter (dbh < 2.5 cm) or not. In this study, small diameters minimum dbh of 1 cm was considered; whereas, in the above cited studies, the authors considered a dbh ≥ 10 cm, except Sambaré et al. (2011) who considered a dbh ≥ 5 cm. The other reasons for the high diversity of woody plants from one location to another may be related to the intensity and frequency of floods, variation in topography, variations in climate and disturbances regimes imposed on the riparian forest by upland environment (Naiman et al., 2008).
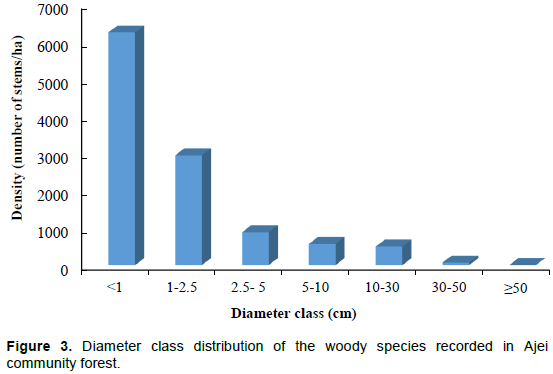
The Shannon-Weaver diversity index (H’) ranged from 3.88 to 4.89 in different forest patches with a value of 4.34 for the entire forest, reflecting the high diversity within this study site. Forest communities considered rich are characterized by a Shannon diversity value of about 3.5 or higher (Kent and Coker, 1992). The high diversity is also confirmed by a Simpson’s diversity index close to 1.0 (0.94); the low generic diversity (1.32), and the ocurrence of families and genera with a single species. Such high diversity seems also to derive from a great abundance of rare species: 40 species were represented by three or fewer individuals.
Regarding the family importance value, the most important families were Euphorbiaceae, Apocynaceae, Moraceae, Rubiaceae and Meliaceae. In term of diversity (number of species per family), Rubiaceae, Euphorbiaceae, Moraceae, Meliaceae, Malvaceae and Fabaceae were the best represented families. Except for the Meliaceae family, Natta (2003) and Momo Soléfack et al. (2018) have also found these families among the most represented in Benin and in Koupa Matapit gallery forests, respectively. In Burkina-Faso, the Fabaceae and Rubiaceae were the most tree species-rich families in gallery forests of the Hippopotamus Pond Biosphere Reserve (Bélem and Guinko, 1998). According to Gentry (1988) three families (Rubiaceae, Annonaceae and Euphorbiaceae) are always among the ten most species-rich families in Africa, Asia and the Neotropics.
The gallery combined various savannah and forest species. The most important species with an Importance value index (IVI) greater than 10% were Macaranga occidentalis, X. monospora, R. vomitoria, Tabernaemontana sp. and C. acuminate. IVI is used for prioritizing species conservation, whereby species with low IVI value need high conservation priority compared to the ones with high IVI (Gotelli and Colwell, 2011). In the Ajei upland community forest, 38 species had an IVI less than 0.5%, each. Therefore, conservation priority should be given to those species having low importance value index.
Species frequencies ranged from 8.33 to 100%, and species can be divided into three main groups based on forest patch affinity:
1. Species that occur in all the twelve forest patches and can be described as ubiquitous: There are 6, representing 4.03% of the total species (Macaranga occidentalis, Croton macrostachyus, Ficus sur, Psychotria dorotheae, Rauvolfia vomitoria and Xymalos monospora).
2. Non-regular species: They are 79 (representing 63.70% of species), each of which is present in at least two different forest patches.
3. Rare, forest patch specific, or occasional species: They are characterized by their belonging to a single particular patch; they are 39 species (31.45%).
Stand structure of the gallery forest
The overall mean tree density for Ajei upland community forest was 1820 trees ha-1. When considering only individuals with dbh ≥10 cm, the stem density was 693 stems ha-1. This number is comparable to the range of 641 to 715 stems ha-1 found by Kellman et al. (1998) in gallery forest in the Mountain pine ridge (Belize). Sambaré et al. (2011) found tree densities ranged from 233 to 727 stems ha-1 in gallery forests in Burkina Faso. This density is also within the range of 167 to 1947 individuals per hectare reported by Gentry (1982) for Neotropical forests sampled by different methods.
Values of basal areas found in this study (19.4 to 39.4 m2 ha-1) are nearly similar to those of riparian forests founnd in Burkina Faso (8.25 to 31.02 m² ha-1), but are lower than those found in Benin (35.6 to 45.6 m²·ha-1). Despite possessing stem densities that are higher than average, the basal areas of gallery forests are lower than average for other tropical forests. This can be explained by the stem sizes that tend to be smaller in gallery forests. It may be a supportive stabilisation adaptation to an unstable upland environment yet to be confirmed by level of shoot-root ratio.
The diameter class exhibited a tendency towards an inverted J-curve distribution, showing an important dynamic of tree species through a high potential of natural regeneration. The high number of individual trees with a small diameter highlights that this ecosystem is dominated by young trees. This gallery forest can be characterized as a low-biomass community with many small-stemmed trees, compared to the continuous upland tropical rain forests. Similar results were found in gallery forests in others parts of Africa (Natta, 2003; Sambaré et al., 2011, Fousseni et al., 2014).
The inverted-J curve distribution in Ajei upland community forest with the presence of more individuals in the diameter classes of ≥2.5 show signs of disturbances and can be explained by three principal causes:
1. The annual transhumance phenomenon in the area by the Bororos and locals accompanied by uncontrolled seasonal bush fires in the savannah in the dry seasons, stretching and consuming the understorey of forest patches;
2. The formation of grazing tracks through the forest patches (common to most patches, but particular to Uka, Egunofon, Akatung and Onom) and;
3. The slash and burn agriculture practice of farmers.
All these challenges restrict the growth and lowers the life span of individuals resulting in a continuous annual regeneration process with fewer individuals having the opportunity to recruit into higher diameter classes. The lack of individuals in the larger size classes cannot really be due to illegal logging of bigger trees by the locals for timber and construction purposes (Temgoua et al., 2018), but by the fact that the forest has limited species that grow larger than these diameters. In all, it could be concluded that that grazing and shifting cultivation activities pose a lot of threats to the sustainability of the forest patches and the entire highland watershed community forest. These phenomena were also described by Fousseni et al. (2014) as major threats in riparian forests of Togo.
Concerning the altitude gradient, the ordination by Principal Coordinate Analysis confirmed the diversity and richness of the lower and middle and lower altitude classes (between 1300 and 1900 m) highest amongst the other elevation classes. This decrease can be explained by changes in biophysical factors, altitude and the combination of other human factors including previous land use. These findings corroborate Wouokoue et al. (2017) who also indicated more species distributed in the lower zones in the Bamboutos Mountain in West region of Cameroon.
Implications for conservation and forest management
In Cameroon the aim of community forestry is to increase the level of involvement and participation of local communities in the conservation and sustainable management of natural resources (Minang et al., 2019). It has a dual objective of improving the standard of living of village populations and conserving biodiversity. Although other resources such as non-timber forest products and wildlife are also important for the populations, the most exploited resource in community forests in Cameroon is timber (Cuny, 2011). From the analysis of the floristic diversity of the community forest of Ajei, it appears that this forest is very poor in commercial timber species. Only four species identified are on the list of potentially commercial forest species established by the Cameroonian forest administration (MINFOF, 2014). These are Entandrophragma angolense, Carapa grandiflora, Albizia ferruginea and Albizia zygia. From the above species, E. angolense is the only one listed among species that are easy to commercialize due to the existence of a large market and demand (MINFOF, 2019). The three other species are among species that can be used, but currently have low commercial value. It is then obvious that the primary objective of the management of the Ajei community forest cannot be the exploitation of timber as is the case for most of the community forests in Cameroon. For example, in Cobaba community forest en Eastern Cameroon, Temgoua et al. (2020) recorded 65 commercial timber species.
One of the options for managing the Ajei community forest could be payment for ecosystem services through its carbon sequestration and watershed protection potential. The study will therefore need to be complemented by an assessment of carbon stocks. Such payments for ecosystem services initiatives, implemented by World Wide Fund for nature (WWF) in some community forests in eastern Cameroon, have had satisfactory and encouraging results for local development (WWF, 2016; Guechoung, 2018). Conservation efforts of the Ajei community forest should be tilted towards upland watershed forest ecosystems as they constitute an essential resource to the sustainability of endemic highland biodiversity and fresh water systems which supply down slope community with drinking water.
Some tree species of economic value providing non-timber forest products were identified in the Ajei community forest. These include Cola acuminata, Dacryodes edulis, Carapa grandiflora, Persea americana and Cola nitida. These species must be valued for the improvement of people’s livelihoods. It is therefore recommended to encourage and promote the planting of these species around compounds and in farmland. This would support the economic development of the Ajei community, while at the same time supporting water catchment protection, carbon sequestration and climate change mitigation. This could be initiated through small scale agroforestry, woodlots, or tree planting projects by providing seedlings of desired species to the local community. Salami et al. (2002) showed this is quite plausible especially among women folk.
Vegetation disturbances related to slash and burn agriculture, livestock and grazing were noted. These land uses are often conflicting with frequent farmer-grazer conflicts and grazing-water catchment conflicts, especially in the dry season, within the Community Forestry and Family Heritage Zones; given, that the various forests patches have been managed by particular families for decades. Therefore it sounds as though it may be critically difficult to shift the management to new land use policies under Community Forestry - most family settlements around the patches to which they have ancestral attributes and where they are custodians see the community forestry initiative by the government as an attempt to alienate them from their land use rights. Reconciling the rising needs for sustainable exploitation for livelihood security of a local population (coupled with ownership and ancestral rights to the forest patches) on one hand, and biodiversity conservation (with other national environmental benefits as major feeder connecting streams to major hydrological basins) on other hand; have been amongst the challenges faced by the Ajei Forest Community Management Board (AHWaCoFoMB). The Board urgently needs support to address, and successfully implememt, the future participative management plan. The plan should address conservation issues relating to several pressing needs including: Demarcation, fire control, habitat regeneration through viable agroforestry practices, woodlots plantation establishment and continuous environmental education, and finally, sensitization and awareness generation among the population.
This study showed that the gallery forests of Ajei highland community forest have a diversified woody flora. A total of 124 woody plant species were recorded, belonging to 90 genera and 47 families. Despite the high diversity level, the forest is still very poor in commercial timber species. Therefore, this community forest cannot be valorized through timber exploitation as is the case of the majority of community forests in Cameroon. Other options of valorization such as payment for ecosystem services through its carbon sequestration and watershed protection potential need to be explored. A good participative management and monitoring plan for these gallery forests is encouraged as a management tool for the Ajei Forest Community Management Board (AHWaCoFoMB) to help enhance the biodiversity conservation and carbon sink potential of these sites. The study needs to be complemented by an assessment of carbon sequestration potential and the contribution of these gallery forests to climate change mitigation.
The authors have not declared any conflict of interests.
The authors thank the Cameroon Wildlife Conservation Society and Forests and Wetlands Consulting Firm, which provided technical and material support for the realization of this work and Ajei Highlands Watershed Community Forest Management Board (AHWaCoFoMB); especially, the Board Chairman (Dr. Williams Ambeh), the Ajei Cultural and Development Association President (Dr. Joe Njweipi), the village traditional authority (Chief Mbacham IV), and various family heads and forest guides who provided guided forest walks, local species identification and their kind hospitality.
REFERENCES
|
ACNR (2011). Muvumba gallery forest conservation project. Technical report. Association pour la Conservation de la Nature au Rwanda (ACNR).
|
|
|
|
Addo-Fordjour P, Obeng S, Anning AK, Addo MG (2009). Floristic composition, structure and natural regeneration in a moist-semi deciduous forest following anthropogenic disturbances and plant invasion. International Journal of Biodiversity and Conservation 1(2):21-37.
|
|
|
|
|
AHWaCoFoMB (2016). Participatory socio-economic, zoological, and botanical inventory of Ajei highland watershed community forest. Ajei Highlands Watershed Community Forest Management Board.
|
|
|
|
|
Ajonina GN, Tomedi EM (2014). Aquaforests and Aquaforestry: Africa. In Encyclopedia of natural resources: Land. Taylor and Francis: New York, pp. 16-38. http://dx.doi.org/10.1081/E-ENRL-120047504
Crossref
|
|
|
|
|
Ake Assi L (1984). Flora of the Ivory Coast: a descriptive and biogeographical study with some ethnobotanical notes. Ph.D.Thesis, Université d'Abidjan, Côte d'Ivoire.
|
|
|
|
|
Angiosperm Phylogeny Group (APG) IV (2016). An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Botanical Journal of the Linnean Society 181(1):1-20.
Crossref
|
|
|
|
|
Barden JA, Halfacre RG, Parrish DJ (1987). Plant Science. McGraw-Hill, Inc. New York.
|
|
|
|
|
Bélem OM, Guinko S (1998). La biodiversité des galeries forestières du Burkina Faso: Cas de la réserve de la biosphère de la mare aux hippopotames. In Bâ AT, Madsen JE, Sambou B (Eds.), Atelier sur flore, végétation et Biodiversité au Sahel. Aarhus University Press, Aarhus Denmark, AAU Reports 39:75-86.
|
|
|
|
|
Cuny P (2011). State of the community and communal forestry in Cameroon. Tropenbos International Congo basin program, Netherlands.
|
|
|
|
|
Curtis JT, McIntosh RP (1950). The interrelations of certain analytic and synthetic of phytosociological characters. Ecology 31:434-455.
Crossref
|
|
|
|
|
FAO (2015). Global forest resource assessment. Second edition. Food and Agricultural Organisation of the United Nations, Rome, Italia.
|
|
|
|
|
Fousseni F, Marra D, Wala K, Batawila K, Xiuhai Z, Chunyu Z, Akpagana K (2014). Basic overview of riparian forest in Sudanian savanna ecosystem: Case study of Togo. Revue d'Écologie 69:24-38.
|
|
|
|
|
Gentry AH (1982). Patterns of Neotropical plant species diversity. Evolutionary Biology 15:1-84.
Crossref
|
|
|
|
|
Gentry AH (1988). Changes in plant community diversity and floristic composition on environmental and geographical gradients. Annals of the Missouri Botanical Garden 75:1-34.
Crossref
|
|
|
|
|
Gotelli NJ, Colwell RK (2011). Estimating species richness. In Magurran AE and McGill BJ, Editors. Biological diversity: Frontiers and measurement and assessment. Oxford University Press, New York USA.
|
|
|
|
|
Guechoung KP (2018). Payment for Environmental Services Initiative on Community Forests in the Ngoyla Mintom Forest Massif. State of art and perspectives. FASA, University of Dschang, Cameroon.
|
|
|
|
|
Hansen C, Wang L, Song XP, Tyukavina A, Turubanova S, Potapov PV, Stehman SV (2020). The fate of tropical forest fragments. Science Advances 6(11):eaax8574.
Crossref
|
|
|
|
|
Kellman M, Tackaberry R, Rigg L (1998). Structure and function in two tropical gallery forest communities: Implications for forest conservation in fragmented systems. Journal of Applied Ecology 35:195-206.
Crossref
|
|
|
|
|
Kent M, Coker P (1992). Vegetation description and analysis: A practical approach. John Wiley and Sons Ltd, England.
|
|
|
|
|
Kimengsi JN, Bhusal P, Aryal A, Fernandez MVBC, Owusu R, Chaudhary A, Nielsen W (2019). What (de)motivates forest users' participation in co-management? Evidence from Nepal. Forests 10(6):512.
Crossref
|
|
|
|
|
Lambi CM (2001). The impact of human activity on land degradation in some highland regions of Cameroon: Implications for development. In Lambi CM (Ed.), Environmental Issues: Problems and Prospects. Unique Printers, Bamenda.
|
|
|
|
|
Magurran AE (2004). Measuring biological diversity. Oxford and Victoria: Blackwell Publishing Malden.
|
|
|
|
|
Maryudi A, Devkota RR, Schusser C, Yufanyi C, Salla M, Aurenhammer H, Rotchanaphatharawit R, Krott M (2012). Back to basics: Considerations in evaluating the outcomes of community forestry. Forest Policy and Economics 14(1):1-5.
Crossref
|
|
|
|
|
Meave J, Kellman M (1994). Maintenance of rain forest diversity in riparian forests of tropical savannas: Implications for species conservation during Pleistocene drought. Journal of Biogeography 21:121-135.
Crossref
|
|
|
|
|
Minang PA, Duguma LA, Bernard F, Foundjem-Tita D, Tchoundjeu Z (2019). Evolution of community forestry in Cameroon: an innovation ecosystems perspective. Ecology and Society 24(1):1.
Crossref
|
|
|
|
|
MINFOF (2009). Manual of procedures for the attribution and norms for the management of community forests. Second Edition. Ministry of Forests and Wildlife, Yaoundé, Cameroon.
|
|
|
|
|
MINFOF (2014). Standard for operational inventory and geo-referencing of stems. Ministry of Forestry and Wildlife, Yaoundé, Cameroon.
|
|
|
|
|
MINFOF (2019). Management and pre-investment inventory guidelines. Ministry of Forests and Wildlife, Yaoundé, Cameroon.
|
|
|
|
|
Momo SMC, Temgoua LF, Ngueguim JR, Nkongmeneck BA (2016). Comparison of plant communities between primary and secondary tropical forests of Mount Oku, Cameroon. Journal of Ecology and the Natural Environment 8(10):163-174.
Crossref
|
|
|
|
|
Momo SMC, Temgoua LF, Fongnzossie E, Djouda Zangmene R (2018). Vegetation and functional spectra of the forest gallery of Koupa Matapit (West-Cameroon). International Journal of Tropical Geology, Geography and Ecology 42(1):147-158.
|
|
|
|
|
Mori SA, Boom BM, Carvalino AM, Dos Santos TS (1983). The ecological importance of Myrtaceae in an eastern Brazilian wet forest. Biotropica 15:68-70.
Crossref
|
|
|
|
|
Naiman RJ, Latterell JJ, Pettit NE, Olden JD (2008). Flow variability and the biophysical vitality of river systems. Comptes Rendus Geoscience 340:629-643.
Crossref
|
|
|
|
|
Natta AK (2003). Ecological assessment of riparian forests in Benin: Phytodiversity, phytosociology and spatial distribution of tree species. Ph.D. Thesis Wageningen University. Netherlands.
|
|
|
|
|
Ndamason S (2016). Assessment of floristic diversity and carbon stocks in relation to stream. FASA, University of Dschang, Cameroon.
|
|
|
|
|
Ndip EY, Agyingyi CM, Nton ME, Oladunjoye MA (2018). Review of the geology of mamfe sedimentary basin, S W Cameroon, Central Africa. Journal of Oil, Gas and Petrochemical Sciences 1(1):35-40.
Crossref
|
|
|
|
|
Salami AT, Ekande O, Usongo PA (2002). The role of women in an environmentally friendly agro-system: Case study of Ngie Clan, North West Province of Cameroon. African Journal of Environmental Assessment and Management 4(2):39-474.
|
|
|
|
|
Sambaré O, Bognounou F, Wittig R, Thiombiano A (2011). Woody species composition, diversity and structure of riparian forests of four watercourses types in Burkina Faso. Journal of Forestry Research 22:145-158.
Crossref
|
|
|
|
|
SCBD (2009). Sustainable forest management, biodiversity and livelihoods: A good practice guide. Secretariat of the convention on biological diversity (SCBD). Montreal Canada.
|
|
|
|
|
Shmida A (1984). Whittaker's plant diversity sampling method. Israel Journal of Botany 33(1):41-46.
|
|
|
|
|
Sonke B (1998). Etudes floristiques et structurales des forêts de la Réserve de Faune du Dja (Cameroun). Thèse de doctorat, Université Libre de Bruxelles, Belgique.
|
|
|
|
|
Ssegawa P, Nkuutu DN (2006). Diversity of vascular plants on Ssese Islands in Lake Victoria, Central Uganda. African Journal of Ecology 44:22-29.
Crossref
|
|
|
|
|
Temgoua LF, Ajonina G, Woyu HB (2018). Land use and land cover change analysis in Ajei upland watershed community forest, North West Region, Cameroon. Journal of Geoscience and Environment Protection 6(9): 83-99.
Crossref
|
|
|
|
|
Temgoua LF, Momo Solefack MC, Nyong PA, Tadjo P (2020). Floristic diversity and exploitable potential of commercial timber species in the Cobaba community forest in Eastern Cameroon: Implications for forest management. Forest Science and Technology 16(2):56-67.
Crossref
|
|
|
|
|
Veneklaas EJ, Fajardo A, Obregon S, Lozano J (2005). Gallery forest types and their environmental correlates in a Colombian savanna landscape. Ecography 28:236-252.
Crossref
|
|
|
|
|
Wouokoue TJB, Anjah GM, Nguetsop VF, Fonkou T (2017). Floristic diversity of the savannah ecosystems in three altitudinal zones of Bambouto Mountains, West Cameroon. Cameroon Journal of Biological and Biochemical Sciences 25:52-59.
|
|
|
|
|
WWF (2016). Report on the implementation of the Redd project in the Ngoyla-Mintom forest massif through the implementation of integrated sustainable management in the framework of the Dja- Odzala - Minkébé Tri-National Landscape. WWF, Yaoundé, Cameroon.
|
|