ABSTRACT
Coexistence between the TOHO-type and Brazilian extended-spectrum (BES)-type of extended-spectrum β-lactamase (ESBL)-produced by bacteria caused public health issue. Several studies have been reported on the coexistence between blaTEM, blaCTX-M and blaSHV genes in ESBL in broad spectrum enterobacteria. The present study involved the prevalence of coexistence of blaTOHO and blaBES genes in enterobacteria identified in hospitalized and out-patients at Saint Camille Hospital in Ouagadougou (Burkina Faso). Firstly, the study was led by microbiological identification of enterobacteria, secondly antibiogram was performed by diffusion method and finally the molecular characterization was done by conventional polymerase chain reaction (PCR) to search for antibiotic resistance genes blaTOHO and blaBES. The ultraviolet (UV) lamp (Gene Flash) for the photography of gels allowed the visualization of specific bands of TOHO and BES genes. Among 250 strains of Gram negative bacilli collected, 60 strains (24.1%) showed resistance profile to antibiotics used. Molecular characterization showed the coexistence between blaTOHO and blaBES genes in 53.3% in bacteria strains carried by the patients. The highest prevalence was observed in Escherichia coli (34.4%) and Klebsiella pneumoniae (21.9%) strains. For the first time in Ouagadougou, Burkina Faso, this study therefore established the coexistence between blaTOHO and blaBES genes in ESBL produced enterobacteria at Saint Camille Hospital.
Key words: Antibiotic, resistance, extended-spectrum β-lactamase (ESBL), genes, TOHO, Brazilian extended-spectrum (BES).
Antimicrobial resistance constitutes a growing danger to human health in the whole world, but hospitals are more
at risk in holding it (Bradford, 2001).
World Health Organization (WHO, 2018)’s first published antimicrobial resistance surveillance data revealed that there are high rates of resistance to many severe bacterial infections in rich and poor nations. Antimicrobial resistance causes around 700 thousand deaths yearly in the globe and affects health cost (Jasovsky et al., 2016). The main mechanism of the antimicrobial resistance is producing extended-spectrum β-lactamases (ESBLs) enzymes by enterobacteria. A lot of research have been done on the main genes producing ESBLs. The commonest ESBLs include temoneira (TEM), sulfhydryl variable (SHV) and cefotaximase-munich (CTX-M) types (Jacoby and, Munoz-Price, 2005).
The TEM-types ESBLs were derived of TEM-1 and TEM-2. The first resistance gene was reported for the first time in 1965 in Escherichia coli taken from a patient in Athens (Greece) called temoneira (hence the designation TEM) (Zubair et al., 2015). With punctual mutations, the SHV-types ESBLs were obtained from the original SHV-1 enzyme. These enzymes originated from Klebsiella pneumoniae chromosomal penicillinase blaSHV gene variant (Brisse and Verhoef 2001; Haeggman et al., 2004). Actually, over different 180 blaSHV-types ESBLs have been defined globally (Liakopoulos et al., 2016).
CTX-M ESBLs were described for the first time in 1986 in Japan, Germany and France in 1989 (CTX-M-1) and have since expanded widely in the world (Thomson and Moland, 2000). CTX-M was the dominant type of ESBL in the world (Anna et al., 2014). These different types of ESBLs could coexist in certain enterobacteria. The presence of more than two different types of β-lactamases in the same isolates has been reported from India, Germany, Malaysia and Iran (Bora et al., 2014; Schmiedel et al., 2014; Seyedjavadi et al., 2016; Yahya Mohsen et al., 2016).
Apart from the main ESBLs, there were minor types ESBLs such as TOHO-type, BES-type, Pseudomonas extended resistance (PER) type, Vietnam extended-spectrum β-lactamase (VEB) type, Guiana extended-spectrum β-lactamase (GES) type, TEM like activity (TLA) type, and Serratia fonticola (SFO) type which were less studied (Cattoir, 2008).
TOHO-type ESBL is a variant of CTX-M2c (Andres et al., 2005). The blaTOHO gene has been described for the first time at Toho University School of Medicine (Japan) in the urine of a one-year-old girl in E. coli TUH12191 (Ishii et al., 1995). This gene has been notified for the first time in Argentina in Shigella flexneri in the stool of a 33-year -old woman (Andres et al., 2005). TOHO-2 ESBL has also been described as produced by E. coli TUH1083. It is classified as a β-lactamase like enzyme of TOHO-1 group rather than mutants of TEM or SHV enzymes (Ling et al., 1998). BES-type ESBL is less distributed and highly resist Ceftazidime, Aztreonam and not Cefotaxime (Bradford, 2001; Arlet and Philippon, 2003). Bacterial strains that carried these genes caused a variety of nosocomial infections and have become a serious problem in clinical practice (Ma et al., 2002).
The prevalence of coexistence between blaTOHO and blaBES genes in ESBL-producing enterobacteria has not been reported in the literature yet. Researches on β-lactamases at Burkina Faso scale were relatively recent and have already identified the presence of TEM, SHV and CTX-M genes, which are responsible for bacterial resistance in enterobacteria (Zongo et al., 2015). The blaTOHO and blaBES genes were not concerned yet. The objective of this work was to detect cases of coexistence between blaTOHO and blaBES genes in enterobacteria that produce ESBLs at Saint Camille Hospital, Ouagadougou, Burkina Faso.
Type of study
This work is a cross-sectional one done at Saint Camille Hospital, Ouagadougou, Burkina Faso, from September to November 2018. Stool, urine and vaginal swab samples consisted the samples taken from out-patients or hospitalized patients. 250 samples were obtained from 250 patients. Uri Select medium, Hektoen medium and Salmonella-Shigella (SS) media were used to inoculate the samples for the growth of Enterobacteria and then incubated at 37°C for 24 h. Later, for antimicrobial analysis, enterobacteria grown on past media were subcultured on a Mueller-Hinton (MH) medium and incubated at 37°C for 24 h (Gaillot et al., 1999).
Antimicrobial assays
Analytical Profile Index (API 20 E) Identification method was used to identify the bacterial strains. Based on the recommendations of the Committee of antibiogram of the French Society of Microbiology (CASFM/EUCAST 2018), antibiotic susceptibility and resistance test werte done on Mueller-Hinton (MH) medium with pure colonies of enterobacteria. Amoxicillin + Clavulanic acid (Augmentin), Cefotaxime, Ceftazidime, Ceftriaxone and Aztreonam were the antibiotic discs utilized. All Augmentin resistant enterobacteria and one third generation cephalosporin were seen in this study as the producers of ESBL (Pereckaite et al., 2018).
Molecular characterization of ESBLs
Extraction of bacterial DNAs
To extract DNAs from bacteria boiling method was utilized (Ribeiro Junior et al., 2016). The strains were reactivated by culturing on the MH medium for 18 ton at 24 h. A colony isolated was removed from the Petri dish and suspended in 200 µl distilled water aliquoted previously in Eppendorf tubes. It was immersed in a water bath to release the genetic material at 100°C for 15 min. Later, the suspension was centrifuged at 12,000 rpm for 10 min and the supernatant having the released DNA was taken to another Eppendorf tube. Nanodrop-type spectrophotometer (BioDrop UV/Vis DUO, Holliston, USA) was used to determine the DNA concentration.
Molecular analysis
25 μl of the Master Mix, DNA and primers (TOHO and BES) constituted the reaction medium for PCR. PCR program consisted of an initial denaturation at 95°C for 5 min followed by 30 cycles (Denaturation 95°C/59 s, Annealing 50°C/59 s, Elongation 68°C/59 s) and a final extension at 68°C for 5 min. We used Gene Amp Thermocycler PCR System 9700.
The sequences of the primers used in this study provided by Applied Biosystems were TOHO-1A 5'-ATGTGCAGT ACCAGTAA-3' and TOHO-1B 5'-TAGGTCACCAGAACCAG-3' for blaTOHO with 876 bp as molecular weight (Laurent et al., 2000). For blaBES, we used BES-F 5 'TAATAACCCTGACCAAGCCTA 3' and BES-R 5 'CCCTTTCAAAAGTCATAAATC 3' with 879 bp as molecular weight (Bonnet et al., 2000).
Electrophoresis in agarose gel
Agarose gel (1%) used for electrophoresis was made using 1X TBE buffer by adding 8 µl Ethidium bromide (BET) 0.5 µg/ml. This made the bands to be visualized in the UV light. A molecular weight marker (1 kb) was used to perform an electrophoretic migration (110 mv) for 30 min on the PCR products. UV light (Gene Flash) was to visualize the fragments and the images were recorded (Lee et al., 2012).
Data processing
The hospital data were entered in Excel 2013 and Standard Statistical Package for Social Sciences (SPSS) version 17.0 for Windows and the EPI Info version 6.0 software were used to analyze the data. All tests were statistically significant at P-value < 0.05 (WHO, 2019).
A total of 250 samples constituted by 169 stools, 79 urines and 2 vaginal swab samples were collected. Out of 250 samples collected, 60 (24.09%) had shown an antibiotic resistance profile by ESBL production. Among these 60 patients 26 were male and 34 were female, with a sex ratio of 0.75. The ages ranged from 16 months to 95 years with an average age of 36 years. There were 28 hospitalized patients and 32 out-patients.
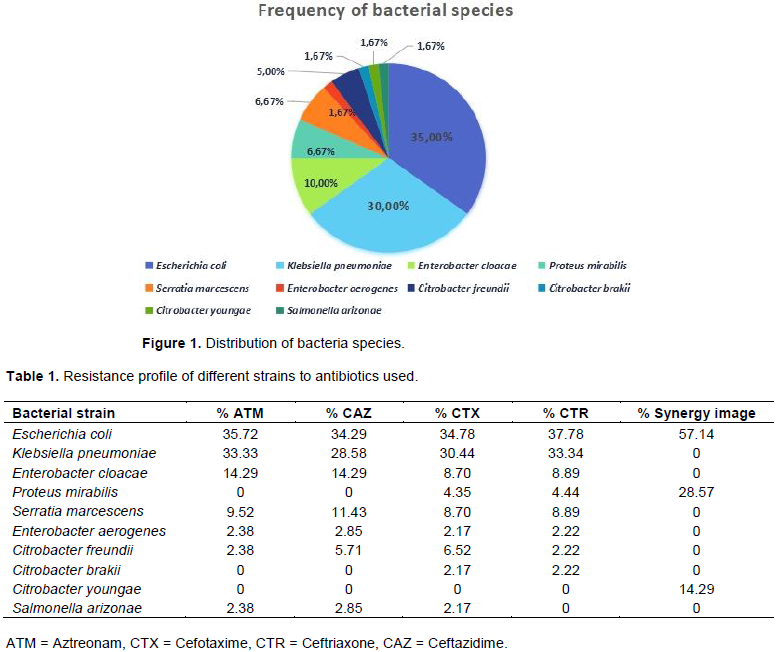
Figure 1 shows the distribution of strains taking part in bacterial resistance by ESBL production.
Results for the sensitivity/resistance of the 60 enterobacteria isolates to the antibiotics tested revealed that 46 strains (76.6%) resist Cefotaxime, 45(75%) resist Ceftriaxone, 42(70%) resist Aztreonam and 35 (58.3%) resist Ceftazidime. Table 1 shows the resistance profiles by bacterial strains. All the strains resist Amoxicillin + Clavulanic acid (Augmentin®).
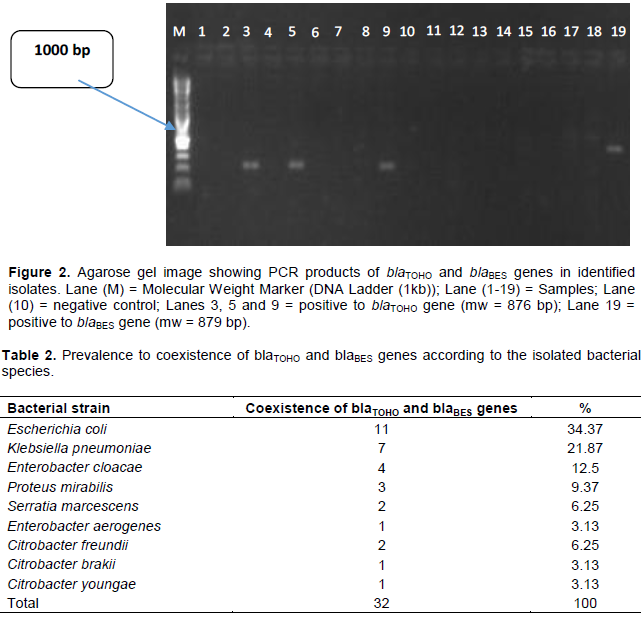
Molecular characterization of the ESBLs by PCR showed that 32 (53.3%) strains were carried blaTOHO and blaBES genes in the same time as displayed by the electrophoresis bands (Figure 2). The distribution to coexistence of blaTOHO and blaBES genes according to the bacterial species is shown in Table 2.
The findings are similar regarding the prevalence of some strains that cause resistance to antibiotic as seen in other works, especially those at Laghouat Hospital, Algeria; 43% E. coli, 30% K. pneumoniae, 20% Enterobacter cloacae and 7% Citrobacter freundii (Lagha, 2015). Other studies that were done at the Charles De Gaulle Paediatric Teaching Hospital (CHUP/CDG) of Ouagadougou, Burkina Faso revealed 47.22% of Escherichia coli; 15.55% K. pneumoniae and 3.33% Klebsiella oxytoca (Mètuor-Dabiré, 2014). The types of ESBLs seen in these works were CTX-M, SHV and TEM. The prevalence of ESBLs produced by Escherichia coli and K. pneumoniae was studied in South America (45.4 to 51.9%) (Villegas et al., 2008) and Saudi Arabia (55%) (Al-Agamy et al., 2009).
These findings show that the total occurrence of ESBLs produced by enterobacteria varies greatly based on geographical zones, nations and various clinics. However, E. coli and K. pneumoniae with high level of ESBL production were the bacterial strains mainly concerned with antibiotic resistance (Lagha, 2015; Villegas et al., 2008).
The antibiotic susceptibility profile of the 60 strains tested revealed resistance to most β-lactams antibiotics: 46 strains (76.6%) resist Cefotaxime, 45 (75%) resist Ceftriaxone, 42(70%) resist Aztreonam and 37 (58.3%) resist Ceftazidime. These could be attributed to improper use of antibiotics. Currently, it is shown using antibiotics, plus third-generation cephalosporin for treatment is very risky in the advancement of bacterial resistance (Mètuor-Dabiré, 2014). Other types of resistance mechanisms can cause these levels of antibiotic resistance such as modifying the permeability of the membrane, modifying antibiotic target, changing metabolic pathway or the efflux phenomena (Munita and Arias, 2016).
The molecular characterization of the 60 bacteria strains by classical PCR revealed the coexistence of TOHO and BES genes in 32 (53.3%) of enterobacteria strains, which makes antibiotic resistance more serious. The antibiotic resistance profile of these different bacteria strains with TOHO and BES genes coexistence was not significantly different from the antibiotic resistance profile of the bacteria strains producing the TOHO and BES enzymes separately in the present study.
TOHO-1 enzymes have been described for the first time in Japan (Tetsuya et al., 1997) and were structurally very close to CTX-M and were therefore classified among this group (Bonnet et al., 2004). This type of ESBL (CTX-M) was frequently encountered in hospitals (Paterson and Bonomo, 2005). This could explain the presence of TOHO enzymes in the present study. The first detection of TOHO-1 outside Japan was reported by Andres (2005) in S. flexneri in the stool of a 33-year-old woman in Argentina (Andres et al., 2005). This bacterial strain expressed an enzyme belonging to CTXM2c whose DNA sequencing gave TOHO-1. There were two types of TOHO enzymes (TOHO-1 and TOHO-2) and their precise prevalence has never been reported in an epidemiological study and the truth of the sequence has been questioned because it was so closely related to CTX-M-2 (Hawkey, 2008).
BES-1 was distantly related to either the CTX-M or GES-1 enzyme that has been isolated from South American isolates. The origin of BES-1 thus remainsunknown (Bonnet et al., 2000).
BES-1 was first described in Serratia marcescens in Brazil in 1996. This strain (S. marcescens) had a very high level of resistance to Aztreonam; distinctly high to Cefotaxime than to Ceftazidime (Bonnet et al., 2000).
In the present study, 21 strains, more than 55% of the strains carrying the BES gene were resistant to Aztreonam. Philippon found that BES-1 ESBLs, characterized by a high level of resistance to Ceftazidime and sometimes to Aztreonam rather than Cefotaxime, have a less wide distribution than the CTX-M group (Philippon and Arlet, 2006). The presence of the BES gene in our study could be explained by the variation of ESBLs frequencies between different geographical areas (Lagha, 2015).
In the presence of coexistence between blaTOHO and blaBES genes in the present study, about antibiotic-resistant bacteria strains could be explained by the spread of these genes in Africa and at Saint Camille Hospital in Ouagadougou, HOSCO (Burkina Faso) or through improper use of antibiotic in animal and human health. The genetic support of TOHO and BES genes carried by enterobacteria in the present study if there were plasmids or integrons could be easily transmitted between several bacteria. Most studies found in the literature have described the isolated presence of the TOHO and BES genes in ESBL-producing enterobacteria (Tatsuro et al., 2002; Liakopoulos et al., 2016).
The present study did not show a significant association between TOHO and BES genes with a particular bacterial species. The distribution of these coexistences was homogeneous in all the bacterial strains of our study (Table 2). E. coli and K. pneumoniae were the bacterial species which showed in the study the most prevalence of coexistence genes (TOHO and BES).
The study of Ibrahimagic et al. (2017) showed the coexistence of more than two types of β-lactamases detected in 77.3% of in-patients and 45.2% in out-patients. Among the isolates of the in-patients, Klebsiella species and E. coli were the most common to produce more than two types of genes, respectively in 65 and 12% of cases. These results showed that the coexistence of β-lactam resistance genes is frequent but their distribution between bacteria like K. pneumonia and E. coli is not homogeneous depending on the geographical area (Canton et al., 2012). Others studies have found 11 multidrug-resistant ESBL-producing strains harboring both CTX-M and SHV as well as TEM and SHV (Mètuor-Dabiré et al., 2018). Our study is the first to have highlighted the coexistence between two types of ESBL, TOHO and BES.
This study made it possible to know the distribution of the coexistence of certain antibiotic resistance genes (TOHO and BES) in Enterobacteriaceae encountered at Saint Camille Hospital in Ouagadougou (Burkina Faso). It also revealed that alongside the CTX-M, TEM and SHV genes, there were other rare types such as TOHO and BES which coexist in Burkina Faso.
For an effective fight against the emergence and diffusion of these bacterial strains multi-resistant to antibiotics, we recommend the population to use antibiotics in a reasonable way and the proper use of hygiene measures at hospitals and doctors should monitor treatment with antibiotics. Our future investigations will focus on the sequencing of the coexistence genes of BES and TOHO.
The authors have not declared any conflict of interests.
The authors wish to thank the CERBA/LABIOGENE, University Joseph Ki-Zerbo. They also express their deep gratitude to the Italian Episcopal Conference (IEC).
REFERENCES
|
Al-Agamy MH, Shibl AM, Tawfik AF (2009). Prevalence and molecular characterization of extended-spectrum beta-lactamase-producing Klebsiella pneumoniae in Riyadh, Saudi Arabia. Annals of Saudi Medecine 29(4):253-257.
Crossref
|
|
|
|
Andres P, Petroni A, Faccone D (2005). Extended-spectrum β-lactamases in Shigella flexneri from Argentina : first report of TOHO-1 outside Japan. International Journal of Antimicrobial Agents 25:501-507.
Crossref
|
|
|
|
|
Anna O, Soderquist B, Persson K, Molling P (2014). Characterization of CTXâ€Mâ€producing Escherichia coli by repetitive sequenceâ€based PCR and realâ€time PCRâ€based replicon typing of CTXâ€Mâ€15 plasmids. Apmis 122(11):1136-114.
|
|
|
|
|
Arlet G, Philippon A (2003). Les nouvelles beta-lactamases à l'aube du troisième millénaire. Revue française des laboratoires 352(2003):41-55.
Crossref
|
|
|
|
|
Bonnet R, Sampaio JL, Chanal C, Sirot D, De Champs C, Viallard J, Labia R, Sirot J (2000). A novel class A extended-spectrum β-lactamase (BES-1) in Serratia marcescens isolated in Brazil. Antimicrobial. Agents Chemotherapy 44.11(2000):3061-3068.
Crossref
|
|
|
|
|
Bonnet R (2004). Growing group of extended-spectrum-beta-lactamases: the CTX-M enzymes. Antimicrobial Agents Chemotherapy 48:1-14.
Crossref
|
|
|
|
|
Bora A, Hazarika NK, Shukla SK, Prasad KN, Sarma JB, Ahmed G (2014). Prevalence of blaTEM, blaSHV and blaCTX-M genes in clinical isolates of Escherichia coli and Klebsiella pneumoniae from northeast India. Indian Journal of Pathology and Microbiology 57(2):249-254.
Crossref
|
|
|
|
|
Bradford PA (2001). Extended-spectrum β-lactamases in the 21st century: characterization, epidemiology and detection of this important resistance threat. Clinical Microbiology Reviews 14(4):933-951.
Crossref
|
|
|
|
|
Brisse S, Verhoef J (2001). Phylogenetic diversity of Klebsiella pneumoniae and Klebsiella oxytoca clinical isolates revealed by randomly amplified polymorphic DNA, gyrA and parC genes sequencing and automated ribotyping. International Journal of Systematic and Evolutionary Microbiology 51(3):915-924.
Crossref
|
|
|
|
|
Canton R, Gonzalez-Alba JM, Galan JC (2012). CTX-M enzymes: origin and diffusion. Frontiers in Microbiology.
Crossref
|
|
|
|
|
CASFM/EUCAST (2018). Antibiotic Committee of the French Microbiology Society. Recommendation 2018.
|
|
|
|
|
Cattoir V (2008). Les nouvelles β-lactamases à spectre étendu (BLSE).
View.
|
|
|
|
|
Gaillot O, Camillo PD, Berche P, Courcol R, Savage C (1999). Comparison of CHROMagar Salmonella Medium and Hektoen Enteric Agar for Isolation of Salmonellae from Stool Samples. Journal of Clinical Microbiology 37(3):762-765.
|
|
|
|
|
Haeggman S, Lofdahl S, Paauw A, Verhoef J, Brisse S (2004). Diversity and evolution of the class A chromosomal beta-lactamase gene in Klebsiella pneumoniae. Antimicrobial Agents Chemotherapy 48.7 (2004):2400-2408.
Crossref
|
|
|
|
|
Hawkey PM (2008). Prevalence and clonality of extended-spectrum β-lactamases in Asia. Clinical Microbiology and Infection 14(2008):159-165.
Crossref
|
|
|
|
|
Ibrahimagic A, Usonovic S, Bedenic B (2017). Prevalence of coexistence genes and clonal spread of ESBL-producing isolates causing hospital-and community-acquired infections in Zenica-Doboj Canton. Bosnia and Herzegovina. Journal of Health Sciences 7(2):80-90.
Crossref
|
|
|
|
|
Ishii Y, Akira O, Hayao T (1995). Cloning and Sequencing of the Gene Encoding a Cefotaxime-Hydrolysing Class A β-lactamase Isolated from Escherichia coli. American Society for Microbiology. 39(10):2269-2275.
Crossref
|
|
|
|
|
Jacoby GA, Munoz-Price LS (2005). The new β-lactamase. The new England Journal of Medicine 352(2005):380-391.
Crossref
|
|
|
|
|
Jasovsky D, Littmann J, Zorzet A, Cars O (2016). Antimicrobial resistance a threat to the world's sustainable development. Upsala Journal of Medical Sciences 121(3):159-164.
Crossref
|
|
|
|
|
Lagha N (2015). Antibiotic resistance study of extended spectrum β-lactamase-producing Enterobacteriaceae (ESBL) isolated from Laghouat Hospital. Alger (2015).
|
|
|
|
|
Laurent P, Isabelle LT, Thierry N, Amal K, Patrice N (2000). Biochemical Sequence Analyses of GES-1, a Novel Class A Extended-Spectrum β-Lactamase, and the Class 1Integron In52 from Klebsiella pneumonia. Antimicrobial agents and Chemotherapy 44(3):622-632.
Crossref
|
|
|
|
|
Lee PY, Costumbrado J, Hsu CY, Kim YH (2012). Agarose Gel Electrophoresis for the Separation of DNA Fragments. Journal of Visualized Experiments (62):e3923.
Crossref
|
|
|
|
|
Liakopoulos A, Mevius D, Ceccarelli D (2016). A Review of SHV Extended-Spectrum β-Lactamases: Neglected Yet Ubiquitous. Frontiers in Microbiology 7:1374.
Crossref
|
|
|
|
|
Ling M, Yoshikazu I, Masaji I (1998). Cloning and Sequencing of gene Encoding Toho-2, a Class A β-lactamase Preferentially Inhibited by Tazobactam. American Society for Microbiology 42(5):1181-1186.
Crossref
|
|
|
|
|
Ma L, Matsuo H, Ishii Y, Yamaguchi A (2002). Characterization of cefotaxime-resistant Escherichia coli isolates from a nosocomial outbreak at three geriatric hospitals. Journal of Infection and Chemotherapy 8(2):155-162.
Crossref
|
|
|
|
|
Mètuor-Dabiré A (2014). Molecular and kinetic characterization of broad-spectrum β-lactamase (ESBL) types of bacterial strains collected at Charles De Gaulle Paediatric Teaching Hospital (CHUP/CDG). Ouagadougou (2014).
|
|
|
|
|
Mètuor Dabiré A, Zongo J, Kaboré B, Zèba B, Baucher M, El Jaziri M, Simporé J (2018). Resistance to β-Lactamines by Gram Negative Bacteria, Producing Several Types of Enzymes , Isolated from Urines in Pediatric Center of Ouagadougou in Burkina Faso. International Journal of Microbiology and Biotechnology 3(4):95-98.
|
|
|
|
|
Munita JM, Arias CA (2016). Mechanisms of Antibiotic Resistance. Microbiology Spectrum 4(2).
Crossref
|
|
|
|
|
Paterson DL, Bonomo RA (2005). Extended-spectrum β-lactamases: a clinical update. Clinical Microbiology Reviews 18(4):657-686.
Crossref05
|
|
|
|
|
Pereckaite L, Tatarunas V, Giedraitiene A (2018). Current antimicrobial susceptibility testing for beta-lactamase-producing Enterobacteriaceae in clinical settings. Journal of Microbiological Methods 152:154-164.
Crossref
|
|
|
|
|
Philippon A, Arlet G (2006). β-Lactamases of Gram negative bacteria: Never-ending clockwork. Annals of Clinical Biology 64(1):37-51.
|
|
|
|
|
Ribeiro Junior JC, Tamanini R, Soares BF, Marangon de Oliveira A, Silva F (2016). Efficiency of boiling and four other methods for genomic DNA extraction of deteriorating spore-forming bacteria from milk. Agricultural Sciences 37(5):3069-3078.
Crossref
|
|
|
|
|
Schmiedel J, Falgenhauer L, Domann E, Bauerfeind R, Prenger-Berninghoff E, Imirzalioglu C, Chakraborty T (2014). Multiresistant extended-spectrum β-lactamase-producing Enterobacteriaceae from humans, companion animals and horses in central Hesse, Germany. BioMed Central Microbiology 14(1):187.
Crossref
|
|
|
|
|
Seyedjavadi SS, Goudarzi M, Sabzehali F (2016). Relation between blaTEM, blaSHV and blaCTX-M genes and acute urinary tract infections. Journal of Acute Disease 5(1):71-76.
Crossref
|
|
|
|
|
Tatsuro S, Akiko I, Shinya F, Takayoshi W, Masaji I, Ishii Y, Hiroshi M (2002). Acyl-intermediate Structures of the Extended-spectrum Class A β-lactamase, Toho-1, in Complex with Cefotaxim, Cephalothin, and Benzylpenicillin. The Journal of Biological Chemistry 277(48):46601-46608.
Crossref
|
|
|
|
|
Tetsuya Y, Hiroshi K, Kazuyoshi S (1997). Nosocomial spread of cephem-resistant Escherichia coli strains carrying multiple Toho-1-like beta-lactamase genes. Antimicrobial Agents and Chemotherapy 41(12):2606-2611.
Crossref
|
|
|
|
|
Thomson KS, Moland ES (2000). Version 2000: The new β-lactamases of Gram negative bacteria at the dawn of the new millennium. Microbes Infection 2(10):1225-1235.
Crossref
|
|
|
|
|
Villegas MV, Kattan JN, Quinteros MG, Casellas JM (2008). Prevalence of extended-spectrum β-lactamases in South America. Clinical Microbiology and Infection 14:154-158.
Crossref
|
|
|
|
|
World Health Organization (WHO) (2018). High levels of antibiotic resistance found worldwide, new data shows.
|
|
|
|
|
World Health Organization (WHO) (2019). Instructions for data-entry and data-analysis using Epi Info™.
|
|
|
|
|
Yahya Mohsen SM, Yahya Mohsen SM, Hamzah HA, Al-Deen MMI, Baharudin R (2016). Antimicrobial susceptibility of Klebsiella pneumoniae and Escherichia coli with extended spectrum β-lactamase associated genes in Hospital Tengku Ampuan Afzan, Kuantan, Pahang. Malaysian Journal of Medical Sciences 23(2):14-20.
|
|
|
|
|
Zongo KJ, Dabire AM, Compaore LG, Sanou I, Sangare L, Simpore J, Zeba B (2015). First detection of bla TEM, SHV and CTX-M among Gram negative bacilli exhibiting extended spectrum β-lactamase phenotype isolated at University Hospital Center, Yalgado Ouedraogo, Ouagadougou, Burkina Faso. African Journal of Biotechnology 14(14):1174-1180.
Crossref
|
|
|
|
|
Zubair M, Malik A, Ahmad J (2015). Microbiology of Diabetic Foot Ulcer with Special Reference to ESBL Infections. American Journal of Clinical and Experimental Medicine 3(1):6-23.
Crossref
|
|