Full Length Research Paper

ABSTRACT
Groundnut farming has always been an essentially major agricultural venture, both commercially and in subsistence, to the inhabitants of the Northern Regions of Ghana. The current study sought to assess the relationship and genetic diversity, through a phylogenetic analysis, among twenty-five (25) Ghanaian groundnut genotypes, in reference to US groundnut mini-core collection, based on the Kompetitive allele-specific PCR (KASP) protocol (a high throughput genotyping platform for marker-assisted selection, studies [LightCycler 480II]) and axiom single nucleotide polymorphism (SNP) array version 2 (48,000 SNP features). It further sought to classify the groundnuts into sub-species based on botanical types, market types, general uses as well as genetic diversity to late leaf spot resistance. The Qiagen Dneasy DNA extraction Kit was used for DNA extraction. Results of the study indicate that majority of the Ghanaian groundnuts evaluated belonged to the Virginia (hypogaea) (large-seeded), Spanish/Valencia (vulgaris/fastigiata) and Mixed (Virginia x Valencia) types based on characteristics, including growth habit (erect/semi-erect/upright/bunch or trailing/spreading) seed or pod size/shape and disease resistance. Thus, significant genetic diversity in morphological characteristics, botanical and market varieties, growth habit and seed size were found among the groundnuts studied. Information generated from this study can be used to develop new groundnut breeding lines that combine these traits to produce higher yields. Marker-assisted selection procedures could help enhance this process based on the availability of QTLs and genes for the traits and markers developed in that regard.
Key words: Axiom, genetic diversity, genotypes, groundnut, Kompetitive allele-specific PCR (KASP), mini-core collection, phylogenetic analysis, disease resistance, single nucleotide polymorphism (SNP).
INTRODUCTION
Groundnut (Arachis hypogaea L.) is an important crop both in subsistence and commercial agricultural production in Ghana (Tweneboah, 2000; Tsigbey et al., 2004). Taxonomically, cultivated groundnut, an annual herb of indeterminate growth habit, is divided into the following two subspecies: hypogaea, with the following key features; absence of flowers on the central or main axis (stem) and regular alternation of vegetative and reproductive branches on the laterals (alternate ramification), with a long life cycle (Holbrook and Dong, 2005; Nigam, 2014). Subspecies fastigiata has the following key features; presence of flowers on the central or main axis (stem) and no specific order of vegetative and reproductive branches on the laterals (sequential ramification), with a shorter life cycle (Holbrook and Dong, 2005; Nigam, 2014).
Groundnut production in Ghana cuts across all the agro-ecological zones, with more than 85% in the Guinea and Sudan savannah zones alone (Atuahene-Amankwa et al., 1988). Nevertheless, there are lesser amounts of production across almost all parts of the country (Tweneboah, 2000).
Due to groundnut’s ability to fix atmospheric nitrogen essentially for related or successive cereal crops such as maize, millet and sorghum, it has over the years formed a vital part of the cropping system in Northern-Savanna Regions of Ghana (Marfo, 1997).
Notwithstanding the aforementioned, average yields over the years have dwindled with figures below one ton per hectare (1.0 t/ha). According to Asibuo et al. (2008), this figure falls clearly below the average potential yields of 2.0 to 3.0 t/ha. In developed countries, average groundnut pod yields stand at 2,500 kg/ha; a figure which is far above average yields in Ghana (840 kg/ha) (FAO, 2002; Nutsugah et al., 2007) and in some other African countries. Poor yields, end of season droughts due to unreliable rainfall, little technology available to small scale farmers, pests and diseases, poor seed quality, inappropriate agronomic practices, cultivation on marginal lands, early leaf spot (Cercospora arachidicola) and late leaf spot (Phaeoisariopsis personata) diseases (Frimpong et al., 2006a) are among the key challenging factors reducing production recorded for the crop over the years (Atuahene-Amankwa et al., 1988).
Groundnut landraces or local varieties are the major sources of planting resources accessible to most farmers in the Northern parts of Ghana (Atuahene-Amankwa et al., 1990; Adu-Dapaah et al., 2004). Only handful farmers have opportunity to access and use improved seeds (Naab et al., 2005).
Core collection of groundnut is effective at improving the effectiveness at which relevant genes of interest are identified in the whole germplasm collection (Holbrook and Dong, 2005). However, traits of interest that are generally hard and expensive to measure are better studied with a much smaller sub-group of germplasm. A phylogenetic analysis is capable exposing the relationship among groundnuts by identifying and classifying a sub-set of the core collection as per similarities and differences in the physical and genetic characteristics (sub-species). Ghanaian groundnuts cultivars in reference to US groundnut mini-core is aims to evaluate the usefulness of this subset of germplasm to identify genes of interest to aid in a future breeding program.
Cluster analysis from a phylogenetic tree partitions the groundnut core accessions into groups (sub-species), which are genetically similar in theory (Holbrook and Dong, 2005). Mini-core collection may therefore be useful for improving the effectiveness of identifying desirable traits in the core collection. For instance, a trait such as resistance to late leaf spot (Cercospora sp.), the use of a mini-core collection would have enhanced the effectiveness of identifying late leaf spot resistant cultivars in the entire collection (Holbrook and Dong, 2005).
Due to the growing demand for groundnuts, a corresponding and sustained increase in groundnut productivity is necessary. Unfortunately, conventional approaches to breed these desired characters are insufficient (Pandey et al., 2012). The use of molecular markers in this crop improvement efforts are significant (Semagn et al., 2014).
The main objective of the current study was to perform phylogenetic analysis to study the relationship and genetic diversity among 25 Ghanaian groundnut genotypes in reference to US groundnut mini-core collection, based on KASP protocol and axiom SNP array version 2 (48,000 SNP features). The study further sought to classify the groundnuts into sub-species based on botanical and market types as well as general uses.
MATERIALS AND METHODS
Source of genetic materials
Twenty-five (25) groundnut genotypes (Table 1 and Plate 1) and United States groundnut mini-core (a core of the core collection) made up of one hundred and twelve (112) out of a total of eight-hundred and thirty-one core accessions (831) (Holbrook and Dong, 2005; Holbrook, 1997; Chamberlin et al., 2010) was followed to score and/or identify SNPs in the groundnut genomes, and subsequently draw a phylogenetic tree. This mini-core had been developed to represent the U.S. Arachis hypogaea L. germplasm collection. This core collection had been shown to be effective in improving the efficiency of identifying genes of interest (late and early leaf spot disease resistance, peanut root-knot nematode, tomato spotted wilt virus and aflatoxin resistance) in the entire germplasm collection.
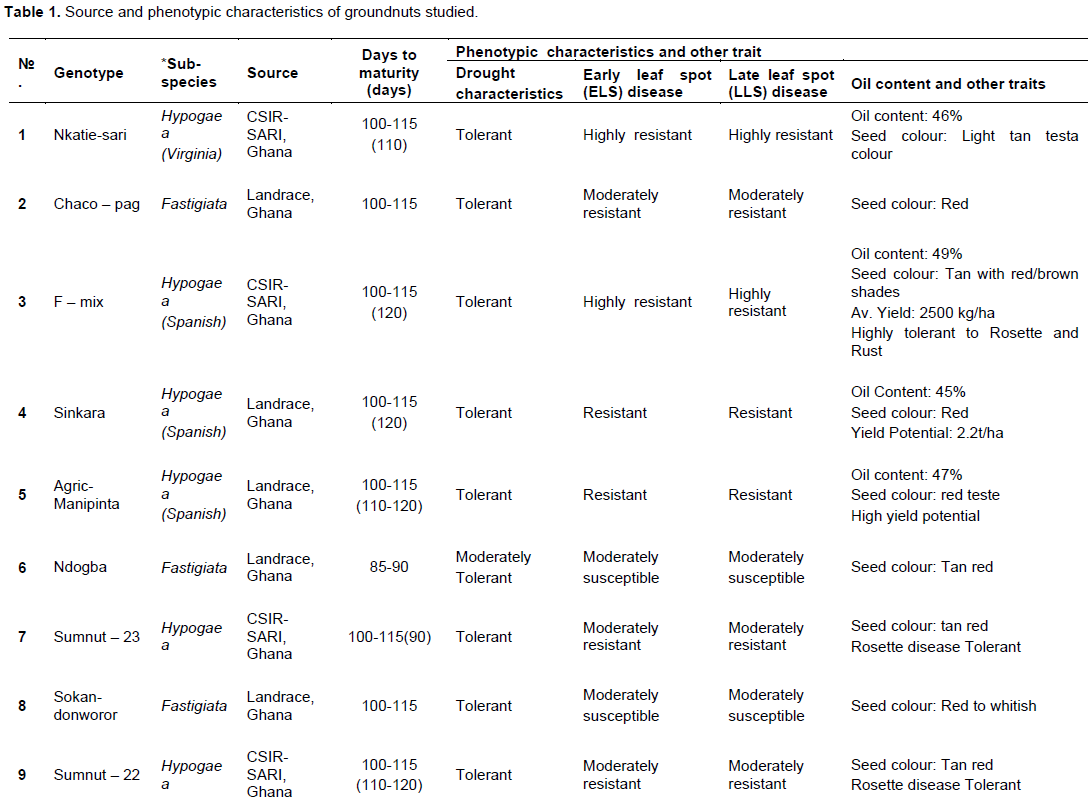
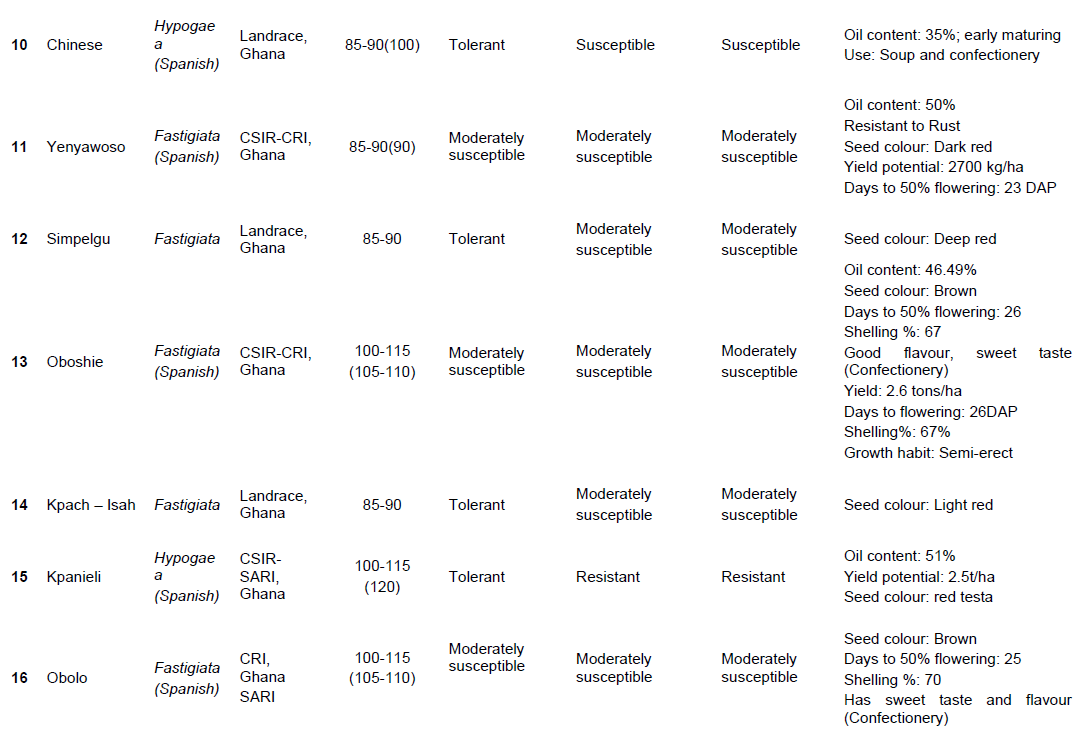
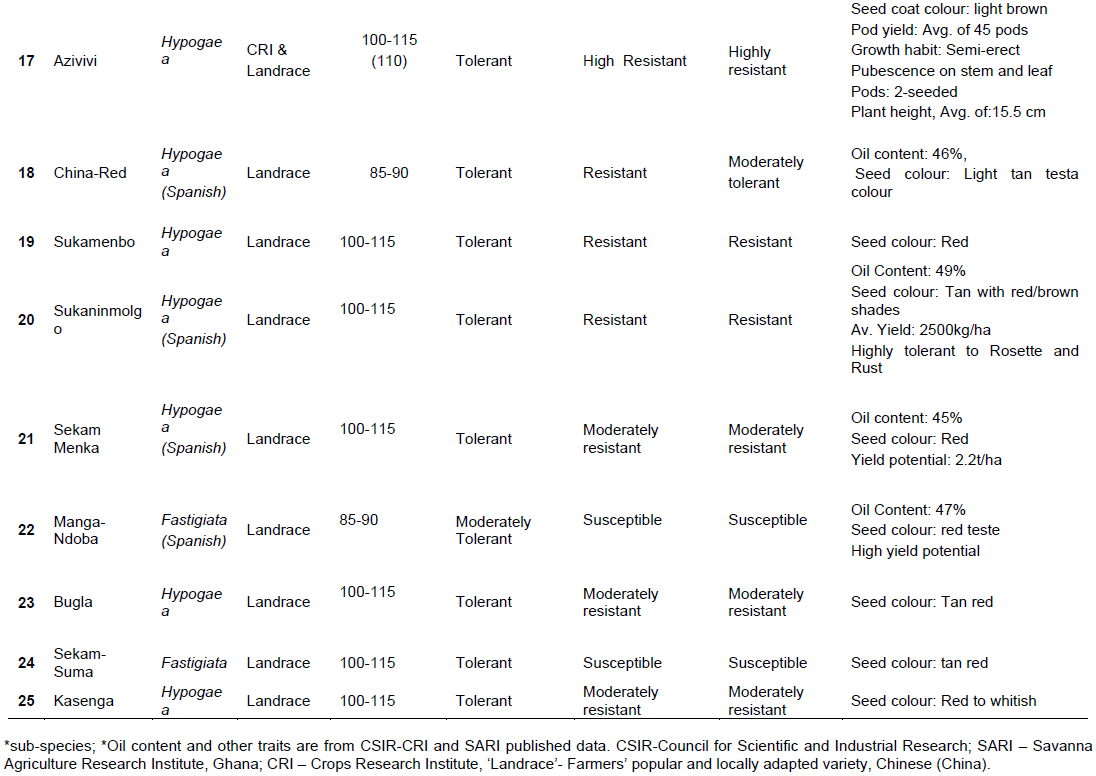

Drought and disease score and assessment: Visual ratings and phenotypic evaluation
Groundnut plants which showed symptoms of disease (early leaf spot, late leaf spot, Rust and Rosette diseases) and drought beginning from 75 DAP were recorded. Severity of disease and drought incidence was scored on a scale of 1 to 5, where 1= no symptoms (0% of crop foliage or drought affected; highly resistant), 2= slight symptoms (25-50% of crop foliage affected; resistant), 3 = moderate symptoms (50-75% of crop foliage affected; moderately resistant), 4 = severe symptoms (>75% of crop foliage affected; susceptible to disease or drought) and 5 = very severe symptoms (about 100% of crop foliage affected; highly susceptible to disease or drought) (Olurunju et al., 1991; Nageswara and Nigam, 2003).
Genotyping groundnuts
Laboratory activities and evaluations for the phylogenetic analysis
Axiom SNP array version 2 (48,000 SNP features): DNA samples were extracted from one-week-old germinating tissue (radicle) of twenty-five (25) groundnut genotypes using the Qiagen Dneasy DNA extraction Kit protocol (www.qiagen.com) (with slight modifications), described as follows: A suitable piece of the tissue was isolated and placed in a UV cross-linked 1.5 mL tube.
One hundred and eighty litres (180 L) of Buffer ATL and 20 L Proteinase K were added and vortexed.
The sample was placed in a 55°C incubator for 3 h. The sample was removed from incubator, vortexed, and 200 L buffer AL added and vortexed again. The sample was then placed in a heat block at 70°C for 10 min.
Two hundred litres (200 L) of 100% ethanol was added and the entire volume transferred onto spin column. It was then centrifuged at 8000 rpm for 1 min; flow-through was then discarded. Five hundred litres (500 L) of Buffer AW1 was added and centrifuged at 8000 rpm for 1 min; flow-through was discarded again.
Five hundred litres (500 L) of buffer AW2 was added and centrifuged at 13000 rpm for 3 min; flow through was again discarded. The spin column was placed on UVcross-linked 1.5 mL tube, 200 L buffer AE was added; allowed to settle for 1 min, then centrifuged at 8000 rpm for 1 min. The step was repeated and then flow-throughs combined for a total volume of 400 L.
DNA quality checks, through gel electrophoresis and quantification were performed prior to PCR analysis. The reproducibility of the DNA profile was tested by repeating PCR amplification.
KASP assay: The KASP (PCR) assay protocol (a high throughput genotyping platform for Marker-Assisted Selection, MAS, studies) (Semagn et al., 2014) using the LightCycler 480II, and involving the twenty-five (25) groundnut genotypes and United States groundnut mini-core (a core of the core collection) made up of one hundred and twelve (112) out of a total of eight-hundred and thirty-one core accessions (831) (Holbrook and Dong, 2005; Holbrook et al., 1997; Chamberlin et al., 2010) was followed to score and/or identify SNPs in the groundnut genomes. This mini-core had been developed to represents the U.S. Arachis hypogaea L. germplasm collection. This core collection had been shown to be effective in improving the efficiency of identifying genes of interest (late and early leaf spot disease resistance, Peanut Root-Knot Nematode, tomato spotted wilt virus and Aflatoxin resistance) in the entire germplasm collection.
Analysis of KASP genotyping data using cluster plots
The Kompetitive allele specific PCR genotyping system (KASPTM) was adopted, and the procedure is as follows: The SNP-specific KASP Assay mix and the universal KASP Master Mix were added to the DNA samples (Figure 1); a thermal cycling reaction was then performed, followed by an end-point fluorescent reading. Bi-allelic discrimination was achieved through the competitive binding of two allele-specific forward primers, each with a unique tail sequence that corresponds with two universal FRET (Fluorescence Resonant Energy Transfer) cassettes; one was labelled with FAM™ dye and the other with HEX™ dye (Figure 1) (www.lgcgenomics.com; Semagn et al., 2014).
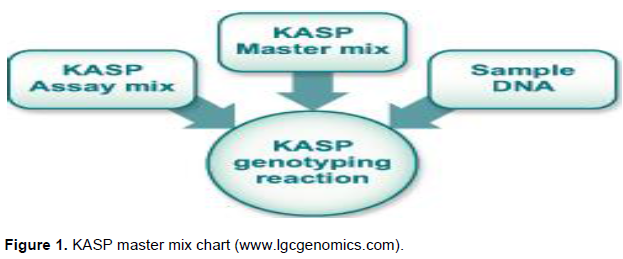
Once the KASP reactions were complete, and the resulting fluorescence had been measured, the raw data was interpreted to enable genotypes to be assigned to the groundnut DNA samples. This was typically achieved by viewing the fluorescence values for each sample on a Cartesian plot (cluster plot) (www.lgcgenomics.com; Semagn et al., 2014).
Visualizing KASP genotyping results using a cluster plot
The fluorescent signal from each individual groundnut DNA sample was represented as an independent data point on a cluster plot. One axis was used to plot the FAM fluorescence value (typically the X-axis) and the second axis was used to plot the HEX fluorescence value (typically the Y-axis) for each sample (Figure 2).
A sample that was homozygous for the allele reported by FAM only generated FAM fluorescence during the KASP reaction. This data point was plotted close to the X-axis, representing high FAM signal and no HEX signal (blue data points), as shown in Figures 2. A sample that was homozygous for the allele reported by HEX only generated HEX fluorescence during the KASP reaction. This data point was plotted close to the Y-axis, representing high HEX signal and no FAM signal (red data points), as in Figures 2. A sample that was heterozygous contained both the allele reported by FAM and the allele reported by HEX. This sample generated half as much FAM fluorescence and half as much HEX fluorescence as the samples that were homozygous for these alleles. This data point was plotted in the centre of the plot, representing half FAM signal and half HEX signal (green data points), as in Figures 2. To ensure the reliability of the results, a KASP reaction without any template DNA was included as a negative control. This is typically referred to as a no template control or NTC. The NTC will not generate any fluorescence and the data point was therefore plotted at the origin (black data points), as in Figures 2.
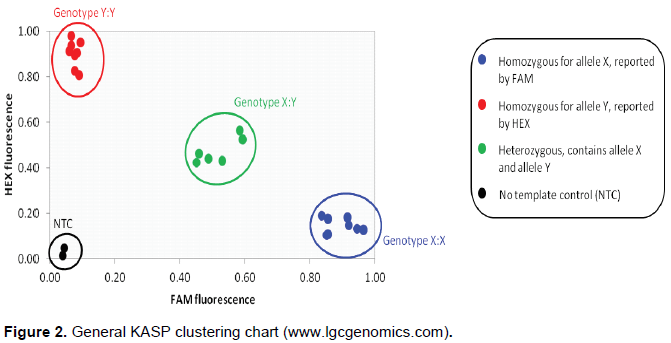
All samples that had the same genotype generated similar levels of fluorescence and therefore all appeared close together on the plot. The genotype of all the data points were determined based on the relative position of the clusters generated (Semagn et al., 2014).
After DNA sample extraction (using the Qiagen Dneasy DNA extraction Kit), gel electrophoresis and quantification of the 25 Ghanaian groundnut genotypes, the KASP PCR assay procedure (KASP PCR protocol, LightCycler 480II), was adopted and the axiom SNP array (version 2 [48,000 SNP features] (Section 6.2.4) protocol used to generate a phylogenetic tree (cluster diagram) in reference to about one hundred and twelve (112) U.S. peanut mini core collection by using the Affymetrix Axiom vs. 2 SNP array software for groundnuts (Figure 3).

RESULTS
Phylogenetic analysis to study the relationship among Ghanaian groundnuts in reference to US groundnut mini-core collection
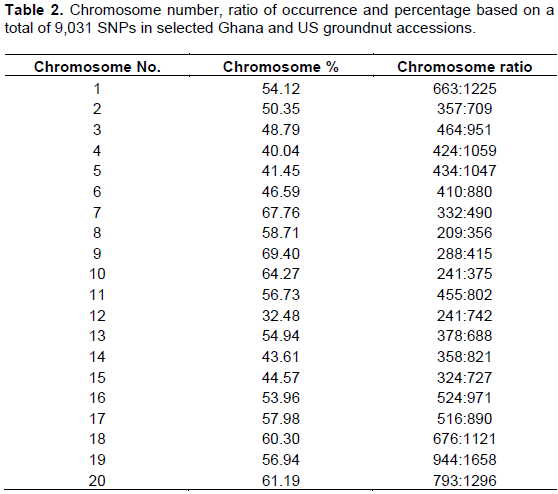
Results from the genotyping as shown in Table 2 indicate that, chromosomes 1, 2, 7, 8, 9, 10, 11, 13, 16, 17, 18, 19 and 20 scored more than fifty percent (>50%) chromosome percentage of the groundnut genotypes. Chromosome 9 recorded the highest percentage of 69.4% followed by chromosome number 7 with 67.76%. The lowest chromosome percentage of the groundnut accessions was recorded by chromosome 12 with 32.48%
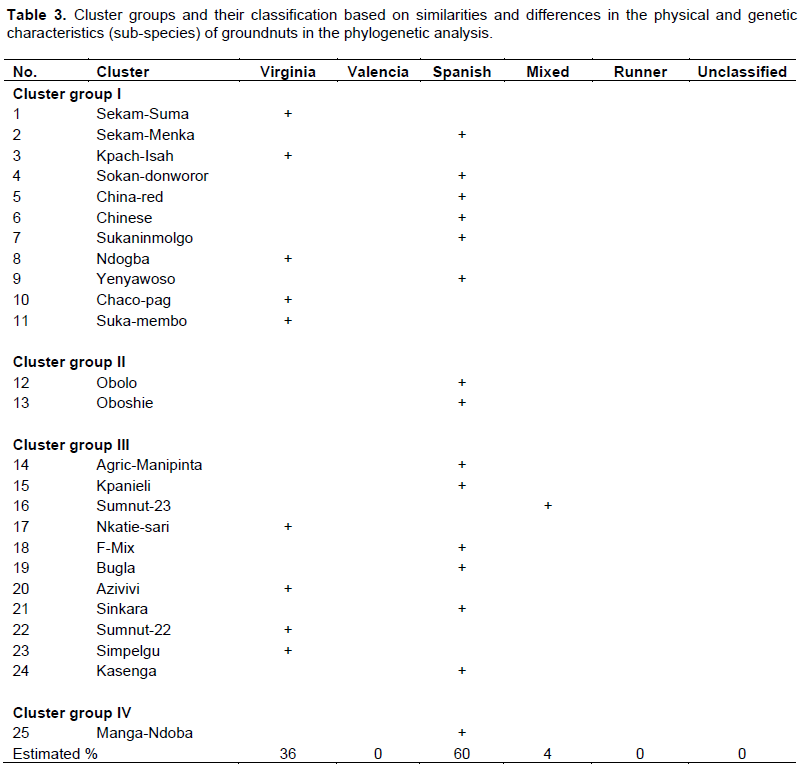
Results from the cluster analysis showed four (4) cluster groups of groundnut sub-species specific to Virginia, Valencia, Spanish, mixed types, runner and unclassified accessions. Clusters I and III (Table 3) each contains the largest of eleven (11) Ghanaian groundnut genotypes whereas cluster IV contained only one (1) genotype. In reference to the US groundnut mini-core collection, majority (60%) of the Ghanaian groundnuts belonged to the Spanish sub-species followed by 36% being in the Virginia sub-species. Only 4% of the groundnuts were ‘mixed’ types (Table 3).
DISCUSSION
Mini-core collection and phylogenetic analysis
Mini-core collection
A peanut core collection of about 831 accessions (112 core of the core collection) had been developed to represent the U.S. Arachis hypogaea L. germplasm collection (Holbrook and Dong, 2005). This core collection has proved to be effective in improving the efficiency at which genes of interest could be identified in the entire germplasm collection (Holbrook and Dong, 2005). Nonetheless, an even smaller sub-set of germplasm was necessary for traits that are difficult and may be expensive to measure (Holbrook et al., 1997). An aspect of the phylogenetic study was effective at identifying and classifying a core of the core collection based on similarities and differences in the physical and genetic characteristics (sub-species) of the Ghanaian groundnuts in reference to the US mini-core so as to evaluate the usefulness of this subset of germplasm to identify genes of interest in groundnuts to aid in a future breeding programme (Holbrook and Dong, 2005).
The cluster analysis from the phylogenetic tree partitioned the core accessions into groups (sub-species), which are genetically similar (Holbrook and Dong, 2005). The mini-core collection, as in the groundnuts, can be used to form the basis for improving the efficiency at which preferred characters (such as disease and drought tolerance) and other unique morphological traits in a population can be identified in the core collection of groundnuts for further improvement. For instance, for tolerance to late leaf spot (LLS) disease caused by Phaeoisariopsis sp., the adoption or use of a mini-core collection would have enhanced the effectiveness and speed for identifying late leaf spot resistant groundnut varieties among the total plant collection (Holbrook and Dong, 2005). According to the cluster groups generated by the phylogenetic tree, majority of the Ghanaian groundnut genotypes were classified as Virginia, Spanish, Mixed and Valencia types (belonging to the hypogaea, fastigiata, hirsuta or vulgaris groups based on their growth characteristics, thus, either erect/semi-erect/upright/bunch or trailing or spreading), small, large or medium seeded or pod size, tan red, light or deep red, mix-coloured/black and white, or off-white. Classification into cluster groups was based more or less on the groundnuts’ genetic and physical (morphological) characteristics in reference to the US groundnut mini-core collection (Krapovikas and Gregory, 1994).
Many groundnut cultivars are grown around the world (Holbrook and Dong, 2005; Nigam, 2014). The major market classes of groundnut grown are Spanish, Runner, Virginia, and Valencia or mixed types (Holbrook and Dong, 2005; Nigam, 2014).
The terms, Virginia, Runner, Valencia and Spanish are used both in botanical sense (as defined above), and as market types (described below) (Holbrook and Dong, 2005; Nigam, 2014).
Virginia (Large-seeded) type/group: Virginia group groundnuts are either bunch or running in growth habit. The bunch type is upright to spreading. It attains a height of 45 to 55 cm (18 to 22 in), and a spread of 70 to 80 cm (28 to 31 in), with 80-to-90 cm (31-to-35 in) rows that seldom cover the ground. The pods are borne within 5 to 10 cm of the base of the plant.
Valencia (fastigiata) type/group: They are comparatively tall, having a height of 125 cm (49 In) and a spread of 75 cm (30 In). Valencia group of groundnuts are usually coarse, and they have heavy reddish stems and large foliage. Groundnut pods are borne on pegs arising from the main stem and the side branches. Most of the pods are clustered around the base of the plant, and only a few are found several inches away. Valencia types normally have 3 to 4 seeded smooth pods with no constriction of the shell between the seeds. The seeds are oval and tightly crowded into the pods. Typical seed weight is 0.4 to 0.5 g.
Spanish (vulgaris) type/group: These have smaller kernels (Holbrook and Dong, 2005; Nigam, 2014). The small Spanish types are commonly grown in South Africa and in the southwestern and southeastern US (Nigam, 2014).
Certain cultivar groups are preferred for particular characteristics, such as differences in flavour, oil content, pod or seed size, shape, and disease tolerance (Holbrook and Dong, 2005; Nigam, 2014). Most groundnuts marketed in the shell are of the Virginia type, along with some Valencias selected for large size and the attractive appearance of the shell (Holbrook and Dong, 2005; Nigam, 2014). Spanish groundnuts are used mostly for peanut candy, salted nuts, and peanut butter (or groundnut paste) (Holbrook and Dong, 2005; Nigam, 2014). Each year, new cultivars of groundnuts are bred and introduced, creating changes in the planting rate, among other things (Holbrook and Dong, 2005; Nigam, 2014).
Virginia (Large-seeded) type/group: Their largest kernels are attractive for direct consumption; their roasted in-shell kernels are consumed as salted groundnuts. The large-seeded Virginia group groundnuts are increasing in popularity due to demand for large groundnuts for processing, particularly for salting, confections, and roasting in the shells, common in Ghana (Holbrook and Dong, 2005; Nigam, 2014).
Runner (small-seeded) type/group: Mainly used in peanut butter (groundnut paste) due to their good flavour, better roasting characteristics and higher yields when compared to Spanish types, leading to food manufacturers' preference for the use in groundnut paste (peanut butter) and salted nuts, particularly common in Ghana.
Valencia (fastigiata) type/group: Valencia types are sweet in taste. Freshly harvested pods are used as boiled groundnuts; dried pods are usually roasted in-shell. This type is used heavily for sale; roasted and salted in-shell groundnuts and peanut butter (Holbrook and Dong, 2005; Nigam, 2014). Varieties include 'Valencia A' and 'Valencia C' (Holbrook and Dong, 2005; Nigam, 2014).
Spanish (vulgaris) type/group: Spanish groundnuts have higher oil content than other types of groundnuts. These have smaller kernels and are used primarily in candy making, as salted groundnuts and in peanut butter (groundnut paste) (Holbrook and Dong, 2005; Nigam, 2014). In the years past (1940s), about 90% of the groundnuts grown in Georgia, USA, were Spanish types (smaller kernels), but the trend since then has been larger-seeded (Virginia group), higher-yielding and more disease-resistant cultivars (Holbrook and Dong, 2005; Nigam, 2014).
Phylogenetic study
Phylogenetic study of the relationship among the Ghanaian groundnut genotypes in reference to US groundnut mini-core collection generally classified majority of the Ghanaian groundnut genotypes into Virginia, Spanish, Mixed and Valencia types, in which case they belonged to the hypogaea, fastigiata, hirsuta or vulgaris groups based on their unique characteristics, thus, erect/semi-erect/upright/bunch or trailing or spreading, small, large or medium seeded or pod size, tan red, light or deep red, mix-coloured/black and white, or off-white. The classification of a variety also included the crop’s tolerance to diseases such as foliar (early, late leaf spots). This classification was based more or less on the groundnuts’ characteristics in reference to the US groundnut mini-core collection, which revealed similar traits among the genotypes (Holbrook and Dong, 2005). Results obtained in the current study were similar to those obtained in an earlier study by Asibuo et al. (2009) in a genetic diversity study of groundnut botanical varieties using SSR markers, and found results that are generally similar to those found in the current study.
Varieties belonging to the hypogaea group are also known as Virginia type when it is large-seeded. They are regarded as Runner types when they are small-seeded. This assertion is confirmed by Nigam (2014), in a similar groundnut study. Variety hirsuta is referred to as Peruvian runner. Variety fastigiata is designated a Valencia type and variety vulgaris is a Spanish type (Nigam, 2014).
Spanish types of groundnut may have originated from a hybridization between Virginia x Valencia types (Nigam, 2014). These could therefore be referred to as ‘Mixed’ types.
The varieties of groundnut so classified as Virginia, Runner, Valencia and Spanish maintain two major usefulness, thus, they have both botanical and market uses. This is confirmed in a study by Holbrook and Dong (2005) and Nigam (2014).
A cross between groundnut of Virginia or Runner or Valencia or Spanish type in a deliberate groundnut breeding programme may result in groundnut genotypes that are desirable both in the botanical sense and/or with high demand or marketability. For example a cross between Virginia type (with large seeds) and a Runner (small-seeded) could result in mixed type of groundnut with medium seed size, which may also combine higher yield and increased oil content.
CONCLUSION
The study revealed that, generally, there were both large-seeded (hypogaea, also referred to as Virginia type) or small-seeded (Runner) groundnut types among the Ghanaian groundnut genotypes. These were either used and/or referred to in botanical sense or as market types. Peruvian runner types of groundnut belonged to the hirsuta group whereas fastigiata types were labeled as Valencia type; Spanish types of groundnut belonging to the vulgaris group. ‘Mixed’ type of groundnut genotypes was also identified. In general, the Virginia types of groundnut were identified as late maturing genotypes whereas the Spanish (and Valencia) types were early maturing. The ‘mixed’ types could possibly pass as intermediate in term of days to maturity.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Holbrook CC, Dong W (2005). Development and Evaluation of a Mini Core Collection for the U.S. Peanut Germplasm Collection. Crop Science 45:1540-1544. |
|
|
Holbrook CC (1997). The U.S. Germplasm Collection of Arachis hypogaea: How Much Diversity Do We Have?. Proceedings of the American Peanut Research Education Society 29:64(Abstr). |
|
|
Krapovikas A, Gregory WC (1994). Taxonomia del genero Arachis (Leguminosae). Bonplandia VIII: 1-187. (In Spanish). (English translation by Williams DE, Simpson CE 2007). Taxonomy of the genus Arachis (Leguminosae). Bonplandia 16(suppl):1-205]. |
|
|
Marfo KO (1997). The performance and association among some important groundnut yield traits in Northern Ghana. Pages 133-140 In: Improvement of cropping system in the savanna zone: the challenges ahead, Mercer-Quarshie, H., Marfo K.O., Langyintuo A.S. and Owusu R. K. (eds.), Proceedings of the third conference on improving farming systems in the savanna zone of Ghana. 11-14 March 1993, Nyankpala Agricultural Experiment Station. Nyankpala, Tamale, Ghana. The Crop Research Institute (CRI)/ Nyankpala Agricultural Experiment Station (NAES) and Deutsche Gesellschaft fur Technische Zusammenarbeit (GTZ) joint project. |
|
|
Naab JB, Tsigbey FK, Prasad PVV, Boote KJ, Bailey BRL (2005). Sowing Date and Fungicide Application on Yield of Early and Late Maturing Peanut Cultivars Grown Under Rainfed Conditions in Ghana. Crop Protection 24:325-332. |
|
|
Nageswara RRC, Nigam SN (2003). Genetic Options for Drought Management in Groundnut. In: (Wright, G.C and Reddy, A. G. S., Eds) Management of Agricultural Drought - Agronomic and Genetic Options. Science Publishers, Inc, 123-141. |
|
|
Nigam SN (2014). Groundnut at a Glance. ICRISAT, Patancheru, P 121. |
|
|
Nutsugah SK, Abdulai M, Oti-Boateng C, Brandenburg RL, Jordan DL (2007). Management of Leaf Spot Diseases of Peanut with Fungicides and Local Detergents in Ghana. Plant Pathology Journal 6(3):248-253. |
|
|
Olurunju PE, Kuhn CW, Demski JW, Misari SM, Ansa AO (1991). Disease Reactions and Yield Performance of Peanut Genotypes Grown Under Groundnut Rosette and Rosette Free Environments. Plant Dis. 75:1269 - 1273. |
|
|
Pandey MK, Monyo E, Ozias-Akins P, Liang X, Guimarães P, Nigam SN, Upadhyaya HD, Janila P, Zhang X, Guo B, Cook DR, Bertioli DJ, Michelmore R, Varshney RK (2012). Advances in Arachis Genomics for Peanut Improvement. Biotechnology Advances 30(3):639-651. |
|
|
Semagn K, Badu RH, Olsen M (2014). Single Nucleotide Polymorphism Genotyping Using Kompetitive Allele Specific PCR (KASP): Overview of the Technology and its Application in Crop Improvement. Molecular Breeding 33(1):1-14. |
|
|
Tsigbey FK, Brandenburg RL, Clottey VA (2004). Peanut production methods in Northern Ghana and some disease perspectives. World Geography of the Peanut pp. 1-10 |
|
|
Tweneboah CK (2000). Modern Agriculture in the Tropics. Co-Wood publishers P 405. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


