Full Length Research Paper

ABSTRACT
A superimposed experiment was conducted at Gircha, Chencha district, Southern Ethiopia to determine the effect of four rootstocks on leaf nutrient concentration of five apple cultivars. The experiment was laid out in a randomized complete block design (RCBD) with three replications. Data on leaf nutrient concentrations were collected and analyzed using SAS software. Results revealed that scion-rootstock interaction significantly (p<0.05) affected leaf nutrients (nitrogen - N, phosphorus - P, potassium -K, calcium- Ca and magnesium - Mg) contents. The maximum N (2.32%), P (0.43%), K (2.09%), Ca (1.88%) and Mg (0.45%) were recorded for combination of Local + Saturn, Local + Scarlet, Begidu + Scarlet, TT-Gala + Mlerose and TT-Gala + Scarlet, respectively; while the lowest values of N (1.3%), P (0.12%), K (1.02%), Ca (0.42%) and Mg (0.11%), were recorded for the combination of Gircha + Saturn, Gircha + Saturn, Begidu + Nela, Gircha + Mlerose and Gircha + Mlerose, respectively. Under Gircha soil condition, Mlerose, Saturn, Alkmen and Scarlett O’Hara cultivars grafted on Begidu, TT-Gala and Local rootstocks were confirmed better in acceptable leaf nutrient concentration. Based on leaf nutrient concentrations TT-Gala, Begidu and Local rootstocks were selected as best compatible with most cultivars grafted.
Key words: Apple, scion-rootstock, nutrient and cultivar.
INTRODUCTION
Apple (Malus domestica L. Borkh) is the most omnipresent of temperate climate fruit tree native to Southwestern Asia and it is a woody plant belonging to the family, Rosaceae and sub-family Maloideae or formerly Pomoideae (Brown et al., 1994). Most of apple cultivars are diploid with chromosome number of 2n = 34 (Sabir and Shah, 2004).
According to Hayesso (2008) and Girmay et al. (2014), apple was introduced to Ethiopia in the 1950’s by Chencha Kalehiwot protestant church in the southern region of Ethiopia that has nearly 50 years of experience in growing highland fruits including apple, pears and plums. Recent data indicate that apple fruit production in Chencha is about 15 metric tonnes per year while the overall country production is estimated to be about 50 metric tonnes (Kebede et al., 2015). Since the total production does not meet the local demand, the country imports about 350 metric tonnes of apple fruits mainly from South Africa, Iran, China and Israel (Holleta Agricultural Research Center (HARC), 2010). Apple is one of the most important temperate fruits grown in the highland climates of Ethiopia by virtue of high altitude (Seifu et al., 2014). The apple fruits are popular and have high economic values in most developed countries because of their multipurpose use and extended shelf life (Hempson and Kemp, 2003).
Leaf mineral content of the grafted scion is greatly influenced by the types of apple rootstocks (Sotiropoulos, 2008; Kebede, 2016). Mineral uptake and the leaf nutrient accumulation of apple cultivars influenced by rootstocks and the differences among the scions-rootstock interaction were attributed to capacity of rootstocks due to their changing root structure (Abrisqueta et al., 2011; Joubert et al., 2011; Fazio et al., 2013; Amiri et al., 2014). This indicates that scion-rootstock difference can determine results. Thus, it can reduce the application of nutrients by selecting better type of rootstocks that have high absorption ability (Keller et al., 2001). Several studies have revealed that rootstocks differ in their effect on the level of nutrients in the grafted variety (Fallahi et al., 2001; Kucukyumuk and Ibrahim, 2011). The differences are due to the interspecific variation among rootstocks in terms of nutrient absorption and the translocation of this property to the variety (Tromp et al., 2005). However, site specific studies are important to determine the types of rootstocks used because the performance of rootstocks are site limited and differ from site to site, as well as region to region (Amiri et al., 2014; Kebede, 2016). The effects of most of the rootstocks studied were not consistent from site to site and varied overtime (Yahya et al., 2004). Apple rootstocks regulate tree growth, precocity, water and nutrient uptake and transport (Koepke and Dhigra, 2013; Kviklys et al., 2014). The rootstock has far more effect on the ultimate size of the tree than the scion cultivar (Tromp et al., 2005; Badiu et al., 2014). In agreement with this, Roy and George (1987) revealed that, rootstocks profoundly affect water and nutrients uptake from the soil especially nitrogen which is of prime importance as it affects the photosynthetic rate, production of carbohydrates and proteins, which in turn control plant growth and development. Kebede and Sabura (2016) showed that influence of the rootstocks on nitrogen (N) content in the petioles was significant.
In the study area, one limited factor that hinders apple expansion is lack of adequate studies on types of rootstocks used and farmers used these four (Begidu, Local, TT-Gala and Gircha) rootstocks without any evidences on the scions-rootstocks compatibility in terms of nutrient uptake. Apple fruit production and establishment of intensive apple orchards has been limited by lack of adequate information on compatibility of rootstock-scion combination (Derbew and Jeong, 2014; Kebede, 2016). Hence, identifying appropriate scion-rootstock combination is expected to alleviate challenges of production and productivity of apple in the area that farmers have been facing yet. Well matched scion-rootstock combination for the production of temperate fruits especially apple is fundamental for its optimal nutrient concentration (Kebede, 2016). It is unwise to adopt scion-rootstock combination from one part of the region to another without an indepth evaluation locally. A mistake made in selecting the rootstock may adversely affect growth and productivity throughout the lifetime of the orchard. Therefore, this study was aimed to determine the effect of locally named different rootstocks on leaf nutrient concentration of different apple cultivars under Gircha Research Center.
MATERIALS AND METHODS
Description of the study area
The study was conducted in the Highland Fruits and Vegetables Research Center of Arba Minch University at Gircha Chencha, Southern Ethiopia. The study site is situated at 6°19' 0'' N latitude and 37°35'0''E longitude with altitude of 3007 m.a.s.l and about 545 km south of Addis Ababa. The district is categorized under Gamo highlands, Enset, wheat and barley livelihood zone and currently known for its high apple fruit production. The soil of Chencha (Gircha) is clay loam in texture with 77% Aluminium (Al) saturation. It has a weak medium sub angular blocky structure with consistence that is friable when moist and sticky/slightly plastic under wet condition with the pH value of 4.8. The topography of the area is hilly, with slope of 13%. The mean minimum temperatures from December to February and maximum temperatures from July to September of the Gircha area are 12 and 23°C, respectively.
Experimental design and treatments
The experiment was carried out by superimposing on two-year-old (planted in August 2015) apple cultivars (Nela, Alkemne, Melrose, Scarlett O'Hara and Saturn) established/grafted on four rootstocks (Begidu, Local, TT-Gala and Gircha). The experiment was laid out in randomized complete block design (RCBD) and the field experiment was designed consisting 20 treatments (scion rootstock) with three replications of 15-trees/plot. Along the rows in each experimental plot, weeds were controlled manually, and 5 kg organic manure was applied for each tree by ring method considering its canopy per a year during vegetative growth period.
Experimental materials
The four rootstocks such as (Begidu, TT-Gala, Gircha and Local (early introduced) as well as five (Nela, Alkemne, Melrose, Scarlett O'Hara and Saturn) scion cultivars were used.
Data collection methods
Soil data
Five sub-samples, from 0-30 cm depth, were collected from 540 cm2 total experimental site representative spots of the entire experimental field by using diagonal sampling method and the composite sample was obtained by vertical insertion of an auger. The working samples were air-dried and made fine by using mortar and pestle. The fined soil was passed through 2-mm sieve and the following soil physico-chemical characteristics were determined in Arba Minch University Chemistry Laboratory by using standard laboratory procedures (FAO, 2008).
Organic carbon content: This was determined by the volumetric method (Walkley and Black, 1934) as described in (FAO, 2008).
pH value: The pH (1:2.5 solid: liquid ratio) of the soil was measured in water using digital pH meter.
Cation exchange capacity (CEC): This was measured after saturating the soil with 1N ammonium acetate (NH4OAc) and displacing it with 1N NaOAc as described by Chapman (1965).
Soil texture: This was analyzed by Bouyoucos hydrometer method and the textural class was determined using the soil textural triangle (Bouyoucos, 1962).
Nitrogen (N): The total N was analyzed using the Kjeldahl digestion, distillation and titration method as described by Schuman et al. (1973).
Phosphorus: Phosphorus contents of samples have been determined by vanadate-molybdate colorimetric method of Wantabe and Oleson (1963).
Potassium (K): K contents were measured with flame photometer.
Calcium (Ca) and Magnesium (Mg): Ca and Mg were measured with an atomic absorption spectrophotometer.
Leaf sample collection
For leaf sample data collection, five trees/rootstock, which have been two years old, were randomly selected and six mature leaves per cultivar (a total of 30 leaves near base of current year’s growth) were collected all around the tree from early May to late June (before dormancy) at the active growth stage season for determination of nutrients concentration as recommended by Kenworthy (1984).
Laboratory analysis of leaves
Sample preparation
The collected leaves were washed in succession of tap water, 0.2% Teepol solution, N/10 HCl and double-distilled water. The fresh and free from contamination leaves were shadE dried and put in an oven at temperature of 70±2°C until constant weight. After drying, leaf samples were grinded with the help of a Wiley mill and then sieved through 1-mm mesh sieve. For the determinations of P, K, Ca and Mg concentrations, 0.5 g of samples were ashed in muffle furnace at 550°C for 8 h, the ash was dissolved in 4 ml 3N HCl and filled up with distilled water as described in Kucukyumuk and Erdal (2011).
Nitrogen
Nitrogen concentration in samples was determined according to modified Kjeldahl method in which 0.5 g sample digested in concentrated H2SO4 and distilled with NaOH (40%). The ammonium N was fixed in H3BO3 (2%) and titrated with 0.1N H2SO4 following the procedure described by Kucukyumuk and Erdal (2011). Finally, data on N were expressed in percent on dry weight basis.
Phosphorus
Tissue samples were analyzed by dry ash digestion method by ashing 0.4 g of plant tissue at 500°C in a forced draft muffle furnace for five hours. The ash was then dissolved in 40 ml of 0.3025 M HCl. The suspensions were filtered and analyzed for elements.
Potassium
K concentrations were determined using flame emission spectrophotometers according to procedure described by Kacar and Inal (2008).
Calcium and magnesium
Ca and Mg concentrations were determined using atomic absorption spectrophotometer according to procedure described by Kacar and Inal (2008).
Statistical analysis
The collected data were subjected to analysis of variance (ANOVA) by using SAS software 9.1.3 and means separation was done using Duncan’s multiple range tests at 5% level of significance.
RESULTS AND DISCUSSION
Leaf tissue nutrients concentration
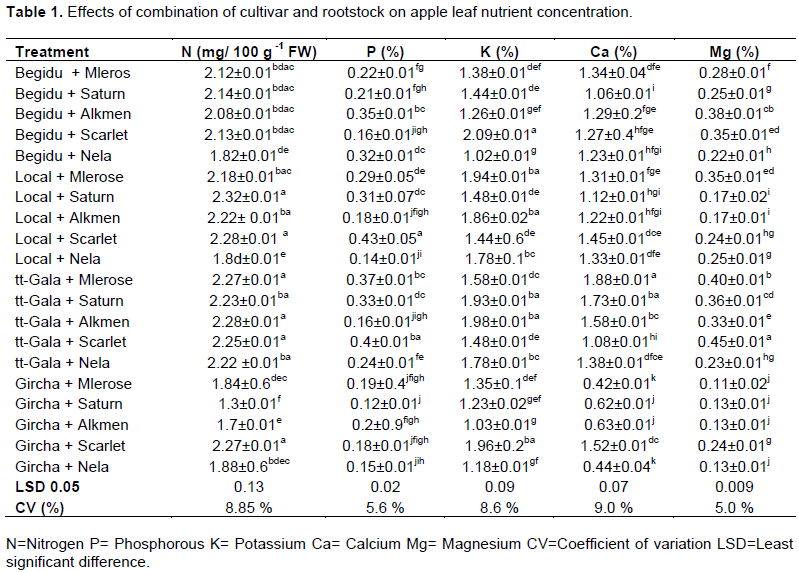
Nitrogen
On basis of results, scion-rootstock interaction had significantly (P<0.05) influenced the leaf N content (Table 2).
Among treatments, cultivar Saturn grafted on Local rootstock recorded the highest N concentration (2.32%), followed by Scarlet O’Hara grafted on Begidu rootstock (2.28%), while the least N concentration (1.3%) was registered for cultivar Saturn grown on Gircha rootstock (Table 2). Among the combinations, (Mleros, Saturn, Alkmen and Scarlett O’Hara + Begidu), (Mlerose, Saturn, Alkmen and Scarlett O’Hara + Local), (Mlerose, Saturn, Alkmen, Scarlett O’Hara and Nela + TT-Gala) and (Scarlett O’Hara + Gircha) recorded optimum nitrogen concentrations within the range of 2.00-2.25% as stated by Heckman (2004) whereas combinations Nela + Begidu, Local and Gircha, Mlerose, Saturn and Alkmen + Gircha registered under deficient range (below 0.2%).
Regarding individual cultivar grafted on Begidu, Local and TT-Gala rootstocks, the value of N content recorded was within the standard or optimum range as stated by Heckman (2004). Whereas cultivars grafted on Gircha rootstock had less value and under deficient range with combination of Saturn and Alkemin on Gircha. This CEC, characteristics of root exudates, due to the interspecific difference among rootstocks with respect to nutrient absorption and the transference of this traits to the scion variety and phenotype of the scion cultivars (Kucukyumuk and Erdal, 2011). In comparing leaf mineral composition of the same cultivar with four different rootstocks and the same rootstock with five different cultivars, there was a lack of consistency between rootstocks and their combination; this indicates that there was a strong influence of rootstocks and scion cultivars in uptake of mineral nutrients. The results agree with Kucukyumuk and Erdal (2011), Joubert et al. (2011) and Fazio et al. (2013). Amiri et al. (2014) reported that, rootstocks had a significant effect on scion leaf mineral concentration in apples. This study confirmed that, the uptake and transport of nutrients to the scion was mainly determined by the type of rootstocks on which the cultivars were grown. These findings are in line with Webster (2004), Abdolhossein et al. (2012) and Kebede (2016) who reported that rootstocks directly influenced the ability of grafted cultivars for absorption of nutrients from the soil.
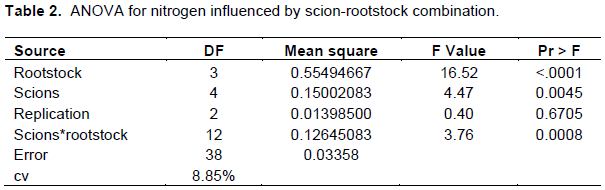
Phosphorous
There was significant (p<0.05) interaction between scion-rootstock combination in viewpoint of total phosphorous concentration (Table 3).
Maximum P concentration (0.43%) was recorded for cultivars Scarlett O’Hara grafted on local rootstock followed by cultivar Scarlett O’Hara grown on TT-Gala rootstock (0.4%) and least was registered for cultivar Saturn grafted on Gircha rootstock (0.12%) (Table 1). Among the treatments, the combinations of (Mleros, Saturn, Alkmen and Nela + Begidu), (Mlerose, Saturn and Scarlett O’Hara + Local), (Mlerose, Saturn, Scarlett OHara, and Nela + TT-Gala) and (Alkmen + Gircha) were recorded within the optimum levels of phosphorus as stated by Heckman (2004) whereas the remaining combinations such as (Scarlett O’Hara + Begidu), (Alkmen, Nela + Local), (Alkmen + TT-G) and (Mlerose, Saturn, Scarlett O’Hara and Nela + Gircha) recorded values within the deficient levels and below 0.2%.
The above mentioned combinations which ranged within optimum (0.2-0.3%) levels for phosphorus concentration may be due to well-matched scion-rootstock combination that favored better up take and translocation of P to the leaves of the cultivars. On the contrary, the deficient levels might be due to less available Phosphorous existence within the soil and compatibility between scion-rootstock interactions and which result in poor phosphorus absorption. These results agree with Kucukyumuk and Erdal (2011), Joubert et al. (2011), Fazio et al. (2013) and Amiri et al. (2014), who reported that, rootstocks had a significant effect on scion leaf mineral concentration in apples.
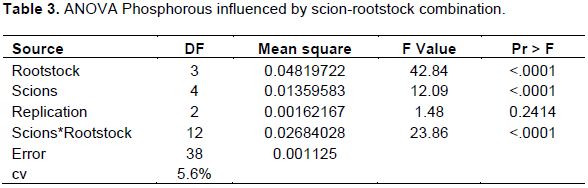
Potassium
Leaf potassium concentration was significantly (p<0.05) affected by different combinations of scion-rootstock interaction (Table 4).
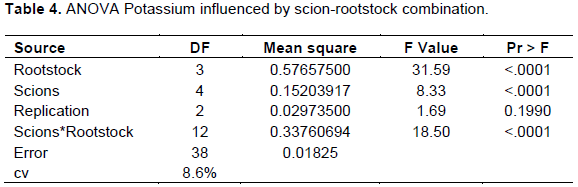
The maximum K concentration (2.09%) was recorded for cultivars Scarlett O’Hara grafted on local rootstock followed by cultivar Alkmen grown on TT-Gala rootstock (1.98%) and least was registered for cultivar Alkmen grafted on Gircha rootstock (1.03%) (Table 1). Among the treatments, combinations of (Mleros, Saturn, Alkmen, Scarlett O’Hara + Begidu), (all cultivar with Local) (all cultivar with TT-Gala), and (Mlerose, Saturn and Scarlett O’Hara + Gircha) were within optimum range (1.25-1.75%) of potassium stated by Heckman (2004), while combination of (Nela, Saturn and Alkmen + Gircha) were recorded under deficient range (below 1.25%); this indicates that the combinations within the optimum range were more efficient in uptake and transport of K+ nutrient whereas combinations in deficient levels implies poor compatibility and inefficient potassium uptake. The other alternative reason was probably due to the variation of CEC. These differences in K+ accumulation in leaf tissues of scion cultivars may be due to variation in absorption capacity of rootstock or differences in the incorporation of K+ ions into xylem and their translocation from root to shoot.
Previous researchers have reported contrasting results. Georgiou (2002) and Tsakelidou et al. (2002) reported that, rootstocks had no effect on K+ concentration. However, the present study confirmed significant effects of rootstocks in absorption of K+, even causing deficiency for certain scion-rootstock combinations. These findings are in line with Abdolhossein et al. (2012) and Kebede (2016) who reported that rootstocks directly influence the ability of grafted cultivars for absorption of nutrients from the soil.
Calcium
The analysis of variance showed that, scion-rootstock interaction was significantly (p<0.05) affected by leaf Calcium concentration (Table 5).
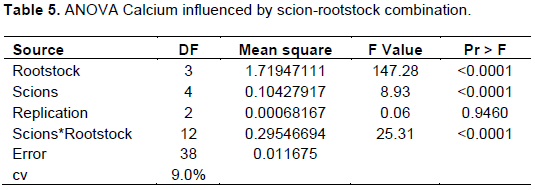
The maximum Ca concentration (1.88%) was recorded for cultivars Mlerose grafted on TT-Gala rootstock followed by cultivar Saturn grown on TT-Gala rootstock (1.73%) and least was registered for cultivar Mlerose grafted on Gircha rootstock (0.42%) (Table 1). Among the twenty treatments, the following combinations (Mlerose, Alkmen, Scarlett O’Hara and Nela + Begidu), (Mlerose, Alkmen, Scarlet and Nela + Local), (Mlerose, Saturn, Alkmen and Nela + TT-Gala) and (Scarlett O’Hara + Gircha) were recorded in the optimum range (1.2-1.6%) of calcium as stated by Heckman (2004) whereas the rest combinations of (Saturn + Begidu), (Saturn + Local), (Scarlett O’Hara + TT-Gala), (Mlerose, Saturn, Alkmen and Nela + Gircha) were recorded within the deficient levels (below 1.2%).
This might be due to the interspecific difference among rootstocks with respect to nutrient absorption and the transference of these traits to the scion variety and phenotype of the scion cultivars (Kucukyumuk and Erdal, 2011). These results are in agreement with earlier findings on apple (Jitender et al., 2005; Wertheim, 1998) who reported that, apple rootstocks’ performance differed due to scion-rootstock interaction. And also these results are in line with study of Kebede (2016) who reported that rootstock effects occur because each rootstock has a different root growth pattern which influences nutrient concentrations.
Magnesium
The interaction effects of scion-rootstock had significantly (p<0.05) influenced leaf Mg concentration (Table 6).
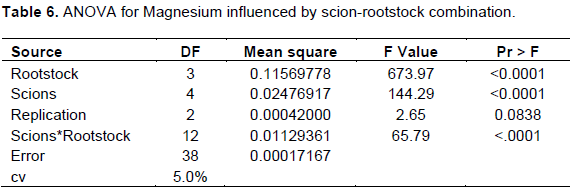
The maximum Mg concentration (0.45%) was recorded for cultivars Scarlett OHara grafted on TT-Gala rootstock followed by cultivar Mlerose grown on TT-Gala rootstock (0.40%) and least was registered for cultivar Mlerose grafted on Gircha rootstock (0.11%) (Table 1). Out of the treatments, the combinations of (Mlerose, Saturn, Alkmen and Scarlet + Begidu), (Mlerose, Scarlett O’Hara & Nela + Local), (Mlerose, Saturn, Alkmen and Scarlett O’Hara + TT-Gala) were recorded in the optimum range (0.25-0.40%) of Magnesium as stated by Heckman (2004) whereas the rest combinations of (Nela + Begidu), (Saturn and Alkmen + Local), (Nela + TT-Gala), and (all cultivar with Gircha) were registered under deficient range (below 0.25%). The combinations ranges below optimum levels might be due to excess K which prevents Mg uptake due antagonistic effect of variations in CEC, characteristics of root exudates, and therefore the correct quantities must be given to reduce Ca related disorders (Shear and Faust, 1980). These findings are in line with Webster (2004), Zarrouk et al. (2005), and Kebede (2016) who reported that, rootstocks directly influence the ability of grafted cultivars for absorption of nutrients from the soil.
CONCLUSION
The results revealed that scion-rootstock combination significantly (P<0.05) influenced leaf nutrient (N, P, K, Ca and Mg) contents. In general, based on the results of the present study, it can be concluded that nutrient concentrations were influenced by different scion-rootstock combination. The nutrient concentration of apple tree at study area can therefore be improved by selecting best compatible scion-rootstock combination. Under the study, soil condition, Mlerose, Saturn, Alkmen and Scarlett O’Hara cultivars grafted on Begidu, TT-Gala and Local rootstocks demonstrated better and adequate leaf nutrient concentration. Based on the present study, it can also be reflected that almost all cultivars grafted on TT-Gala rootstock were well-matched (compatible) for better accumulation of N, P, K, Ca and Mg for most of the apple cultivars studied.
Among the combinations, all scions grafted on Gircha rootstock showed least performances whereas cultivar Nela is inferior in leaf nutrient concentration from all the rootstocks to its leaves. The scion-rootstock combination is fundamental for optimal growth and leaf nutrient concentration. Each rootstock has its own root growth pattern, so that this difference influences leaf nutrient concentrations. Therefore, it should be necessary to accept scion-rootstock combination with an in-depth evaluation locally. In general, the study confirmed that leaf nutrient concentration of the apple scion was determined by the type of rootstocks as well as scion cultivars.
RECOMMENDATION
Based on results it can be determined that rootstock influenced leaf nutrient concentrations of apple cultivars. However, it is difficult to make overall generalized recommendations for suitability of rootstock for particular apple cultivars because some cultivars and rootstocks combinations induced positive effect on nutrient concentrations but did not find it so significant for other important characters. However, based on the combinations of Mlerose, Saturn, Alkmen and Scarlett O’Hara cultivars grafted on Begidu, tt-Gala and Local rootstocks were identified as best compatible for leaf nutrient concentrations. However, the nutrient uptake and translocation of the rootstocks is much determined by the final yield and quality of apple fruit tree. Thus, the research ideas that emerged from this study and the recommendations for further action are as follows:
i) The growth and leaf nutrient concentrations of cultivars under study soil condition were determined by rootstock types used. Thus, rootstocks that are compatible with the cultivars should be identified before grafting.
ii) Miss-combination of scion-rootstock may leaf nutrient concentrations of apple cultivars. Therefore, before grafting, the right rootstock type should be selected by choosing correct combination.
ACKNOWLEDGMENTS
The authors would like to thank Arba Minch University college of Agricultural sciences department of Horticulture for financial support and Research Directorate Office of Gircha High land Fruit and Vegetable Research center (GHFVRC) for allowing the farm land and also their assistants for their invaluable contributions to this research.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Abdolhossein AJ, Hamed H,Mehdi HF (2012). Effect of rootstock type and scion cultivar on citrus leaf total nitrogen.World Applied Sciences Journal 19 (1):140-143. |
|
|
Abrisqueta J, Mounzer O, Álvarez S, Conejero W, García-Orellana Y, Tapia LM, Vera J, Abrisqueta I, Ruiz-Sánchez M (2011).Root dynamics of peach trees submitted to partial rootzone drying and continuous deficit irrigation. Agricultural Water Management 95(8):959-967. |
|
|
Amiri ME, Fallahi E, Safi-Songhorabad M (2014). Influence of rootstock on mineral uptake and scion growth of'golden delicious' and 'royal gala' apples. Journal of Plant Nutrition 37(1): 16-29. |
|
|
Badiu D, Mitre I, Tripon A, Mitre L, Mitre V (2014). Studies on the influence of cultivar, rootstock and culture system on growth and yield in apple. Bulletin UASVM Horticulture 72(1):34-38. |
|
|
Bouyoucos GJ (1962). Hydrometer method improved for making particle size Analysis of soils. Agronomy Journal 54(5):464-465. |
|
|
Brown PH, Zhang Q, Ferguson L (1994). Influence of rootstock on nutrient acquisition by pistachio. Journal of Plant Nutrition 17(7): 1137-1148. |
|
|
Chapman HD (1965). Cation exchange capacity by ammonium saturation. In: Methods of Soil Analysis Chemical and Microbiological Properties. American Society of Agronomy Inc, Publisher, Madison, Wisconsin, USA. |
|
|
Derbew Belew, Jeong CS (2014). Fruit Production in Ethiopia. Kangwon National University Press. P 72. |
|
|
Fallahi E, Chun IJ, Neilsen GH, Michael WC (2001). Effects of three rootstocks on photosynthesis, leaf mineral nutrition, and vegetative growth of Bc-2 Fuji apple trees. Journal of Plant Nutrition 24(6):827-834. |
|
|
Fazio G, Kviklys D, Grusak MA, Robinson TL (2013). Phenotypic diversity and QTL mapping of absorption and translocation of nutrients by apple rootstocks. Aspects of Applied Biology 119:37-50. |
|
|
Food and Agriculture Organization of the United Nations (FAO) (2008). FAO fertilizer and plant nutrition bulletin: Guide to laboratory establishment for plant nutrient analysis, FAO, Rome, Italy P 203. |
|
|
Georgiou A (2002). Evaluation of rootstocks for 'Clementine' mandarin in Cyprus. Scientia Horticulturae 93(1):29-38. |
|
|
Girmay G, Menza M, Mada M, Abebe T (2014). Empirical Study on Apple Production, Marketing and its Contribution to Household Income in Chencha District of Southern Ethiopia. Scholarly Journal of Agricultural Science 4(3):166-175. |
|
|
Hayesso T (2008). Hope in Apples. SNV Netherlands development cooperation case studies. |
|
|
Heckman J (2004). Leaf analysis for fruit trees. In: Malinowski K (Ed.). Rutgers cooperative research and extension, NJAES, Rutgers, The state University of New Jersey, New Brunswick, USA pp. 1-2. |
|
|
Hempson CR, Kemp H (2003). Characteristics of commercial apple cultivars. In: Apples, (Ferre, D.C. and Warrington, I.J. ed.). Cambridge, CABI pp. 62- 70. |
|
|
Holeta Agricultural Research Center (HARC) (2010). Temperate fruit research division. Review paper. |
|
|
Jitender S, Cox B, Clara I (2005). The relationship between anxiety disorder and physical disorders in the U.S. National comorbidity survey Depression and Anxicty 21(4):193-202. |
|
|
Joubert J, Stassen P, Wooldridge J (2011). Effect of rootstock on leaf and fruit macro-element composition in 'Reinders Golden Delicious' apple. Acta Horticulturae 903:391-396. |
|
|
Kacar B, Inal A (2008). Plant Analysis, prognosis and correction of iron chlorosis in peach trees: Influence on fruit quality. Journal of Plant Nutrition 20(11): 1567-1572. |
|
|
Kebede J (2016). Influence of Rootstocks on Nitrogen, Phosphorus and Potassium Content in Leaves of Four Apple Cultivars. Agriculture and Biology Journal of North America 7(4):196-199. |
|
|
Kebede J. Sabura S (2016). Effect of rootstock type and scion cultivar on apple leaf total nitrogen. Agriculture and Biology Journal of North America 8(4):27-31. |
|
|
Kebede J, Masresha F Sheleme B (2015). Effect of Rootstock Type and Low Soil pH on Growth and Yield of 'Anna' and 'Jonagored' cultivars. Scholarly Journal of Agricultural Science 5(3): 67-75. |
|
|
Keller M, Kummer M, Carmo Vasconcelos (2001). Soil nitrogen utilization for growth and gas exchange by grapevines in response to nitrogen supply and rootstock. Australian Journal of Grape and Wine Research 7(1): 2-11. |
|
|
Kenworthy AL (1984). Fruit, nut and plantation crops, deciduous and evergreen, guide for collecting foliar samples for nutrient element analysis. Horticulture Department of Michigan State University, Mimco pp. 1-39. |
|
|
Koepke T, Dhingra A (2013). Rootstock scion somato-genetic interactions in perennial composite plants Plant. Plant Cell Report 32(9):1321-1337. |
|
|
Kucukyumuk Z, Erdal I (2011). Rootstock and cultivar effect on mineral nutrition, seasonal nutrient variation and correlations among leaf, flower and fruit nutrient concentrations in apple trees.Bulgarian Journal of Agricultural Science 17(5):633-641. |
|
|
Kviklys D, Liaudanskas M, Janulis V, Viskelis P, Rubinskiene M, Lanauskas J, Uselis N (2014). Rootstock genotype determines phenol content in apple fruits. Plant, Soil and Environment 60(5):234-240. |
|
|
Roy C, George R (1987). Cultivar effect on adventitious root development of apple rootstock. Horticultural Science 22(1): 57-58. |
|
|
Sabir S, Shah A (2004). Effect of chemical treatment, wax coating, oil dipping and storage behavior of apple. Pakistan Journal of Nutrition 3(2): 122-127. |
|
|
Schuman G, Stanley A, Knudsen D (1973). Automated total nitrogen analysis of soil and plant samples. Proceedings Soil Science Society of America, 37 (3): 480-481. |
|
|
Seifu F, Sabura S, Agena A, Guchie G, Fantahun W, Belete Y (2014). Survey on apple production and variety identification. Journal of Agriculture and Food Technology 4(5):7-15. |
|
|
Shear CB, Faust M (1980). Nutritional ranges in deciduous tree fruits and nuts. Horticultural Reviews 2:142-163. |
|
|
Sotiropoulos Thomas (2008). Performance of the apple (Malus domestica Borkh) cultivar Imperial Double Red Delicious grafted on five rootstocks. Horticultural Science 35(1):7-11. |
|
|
Tromp J, Webster AD, Wertheim SJ (2005). Fundamentals of Temperate Zone Tree Fruit Production. Backhuys Publishers, Leiden, the Netheder lands. |
|
|
Tsakelidou K, Papanikolaou X, Protopapadakis E (2002). Rootstock effects on the yields, tree and fruit characteristics of the mandarin cultivar 'Clementine' on the Island of Rhodes. Experimental Agriculture 38(3):351-358. |
|
|
Walkley A, Black IA(1934). An examination of the Degtjareff Method for Determining Soil Organic Matter, and a proposed Modification of the Chromic Acid Titration Method. Soil Science 37(1):29-38. |
|
|
Wantabe FS, Olsen SR (1963). Test of vanadate-molybdate colorimetric method for determining P in Wantabe and Oleson. Soil Soil Science Society of America, Proceedings 29(6):677-678. |
|
|
Webster A (2004). Vigour mechanisms in dwarfing rootstocks for temperate fruit trees. Acta Horticulturae 658:29-41. |
|
|
Wertheim SJ (1998). Useful differences in growth vigor between sub-clones of the apple rootstock M.9. Acta Horticulture 451:121-128. |
|
|
Yahya, R, Al-Hinai Teryl R (2004). Rootstock effects on growth and quality of gala apples. Horticultural Science 39(6):1231-123. |
|
|
Zarrouk,Zarrouk O, Gogorcena Y, Gómez-Aparisi J, Betrán J, Moreno, M (2005). Influence of almond peach hybrids rootstocks on flower and leaf mineral concentration, yield and vigour of two peach cultivars. Scientia Horticulture 106(4):502-514. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


