Review

ABSTRACT
Potato (Solanum tuberosum L.) is a key alternative to maize crop in Kenya. However, pests and diseases affect the yields. Information on Irish potato virology is continually patchy. Viral disease dynamics require constant updating to track new and novel agents. Efforts to mitigate viruses and crop breeding for tolerance can be determined this way. In Kenya, key potato viruses include: Potato Leaf Roll Virus (PLRV), Potato Virus X (PVX), Potato Virus S (PVS) and Potato Virus Y (PVY). Detection of these viruses has been through symptomatology, serology and nucleic-acid approaches. Molecular biology has revolutionary developments in sequencing technologies influencing diagnosis of plant viruses. Massive parallel sequencing has promoted detection, identification and discovery of novel viruses in plants without use of antibodies or prior virus knowledge. Complete viral genomes can be sequenced from asymptomatic and symptomatic samples. Viral metagenomics, diversity and genome variability can be deduced this way. Next generation sequencing platforms bring robustness, timeliness and affordability to virus detection. However, few studies have attempted to utilize it in unravelling potato virology beyond the routine detectable agents in the country. The current study reviews diagnosis of Irish potato viruses in Kenya against the techniques used, comparing them to next generation sequencing.
Key words: Deep sequencing, next generation sequencing, reverse transcriptase polymerase chain reaction (RT-PCR), serology.
INTRODUCTION
CHARACTERIZATION OF POTATO VIRUSES IN AFRICA
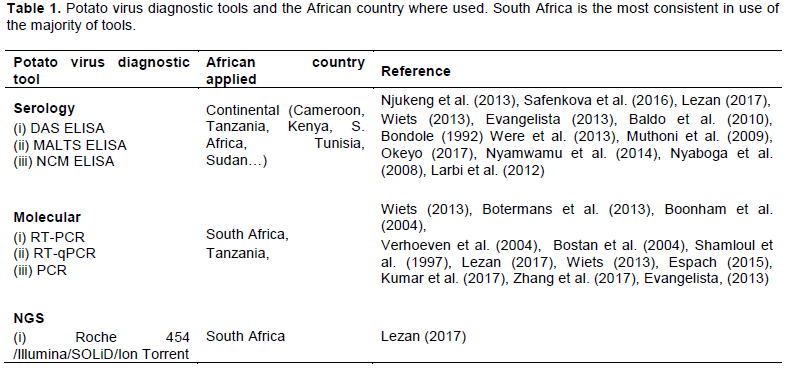
SUCCESS OF NGS IN VIRUS HUNTING AND DISCOVERY
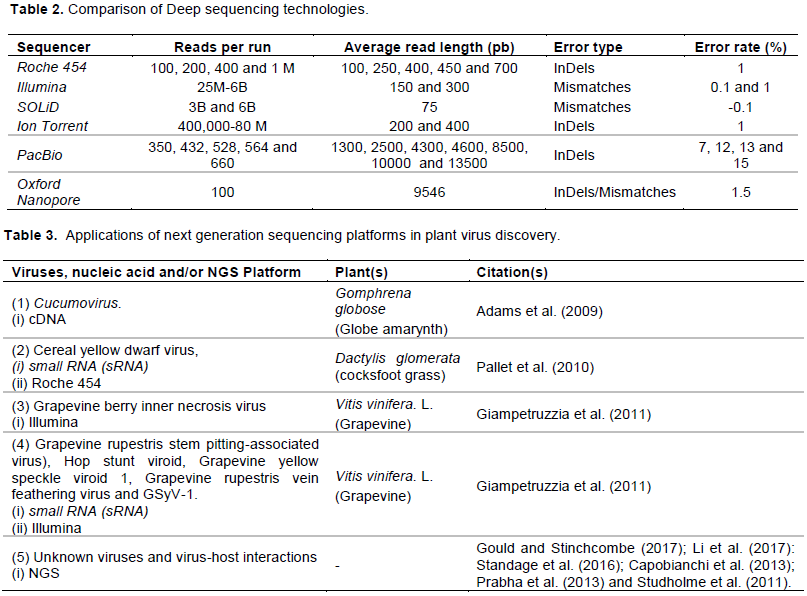
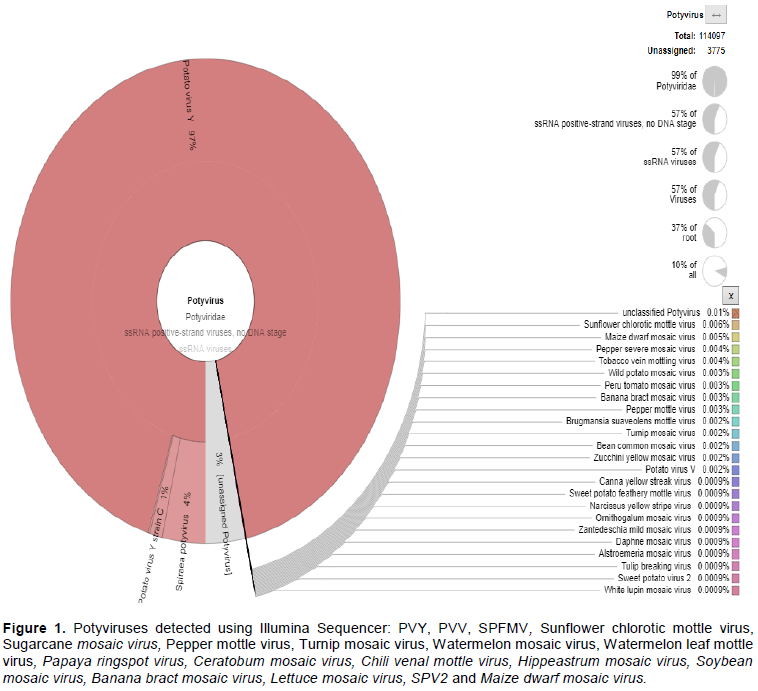
LIMITATIONS OF NEXT GENERATION SEQUENCING
CONCLUSION
RECOMMENDATIONS
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Adams IP, Glover RH, Monger WA, Mumford RA, Jackeviciene E, Navalinskiene M, Samuitiene M, Boonham N (2009). Next-generation sequencing and metagenomic analysis. A universal diagnostic tool in plant virology. Molecular Plant Pathology 10:537-545. |
|
|
Al Rwahnih Maher, Daubert S, Golino D, Islas C, Rowhani A (2015). Comparison of next generation sequsencing versus biological indexing for the optimal detection of viral pathogens in grapevine. Phytopathology 105:758-763. |
|
|
Al Rwahnih Maher, Daubert S, Golino D, Rowhani A (2009). Deep sequencing analysis of RNAs from a grapevine showing Syrah decline symptoms reveals a multiple virus infection that includes a novel virus. Virology Elsevier Inc 387:395-401. |
|
|
Ali M, Hameed S, Tahir M (2014). Luteovirus: Insights into pathogenicity. Archives of Virology 159:2853-2860. |
|
|
Anderson PK, Cunningham AA, Patel NG, Morales FJ, Epstein PR, Daszak P (2004). Emerging infectious diseases of plants: Pathogen pollution, climate change and agrotechnology drivers. Trends in Ecology and Evolution 10s:535-44. |
|
|
Awasthi LP, Verma HN (2017). Current status of viral diseases of potato and their ecofriendly management - A critical review. Virology Research and Reviews 1(4):1-16. |
|
|
Balasubramanian S (2015) .Solexa sequencing: Decoding genomes on a population scale. Clinical Chemistry 61:21-24. |
|
|
Baldo NH, Elhassan SM, Elballa MMA (2010). Occurrence of viruses affecting potato crops in Khartoum State-Sudan. Potato Research 53(1):61-67. |
|
|
Barzon L, Lavezzo E, Militello V, Toppo S, Palu' G (2011). Applications of next-generation sequencing technologies to diagnostic virology. International Journal of Molecular Sciences12:7861-84. |
|
|
Bentley DR, Balasubramanian S, Swerdlow HP, Smith GP, Milton J, et al (2008). Accurate whole human genome sequencing using reversible terminator chemistry. Nature 456:53-59. |
|
|
Bondole BM (1992). Potato virus y (PVY) in Irish potatoes (Solanum tuberosum) and tree tomato (Cyphomandra betaceae) and the influence of potato susceptibility to the virus and the aphid virus-vector on the spread of PVY Mosaic in Kenya. Uonbi.erepository found at |
|
|
Boonham N, Glover R, Tomlinson J, Mumford R (2008). Exploiting generic platform technologies for the detection and identification of plant pathogens. European Journal of Plant Pathology 121:355-363. |
|
|
Boonham N, Kreuze JF, Winter S, van der Vlugt R, Bergervoet J, Tomlinson J, Mumford RA (2014). Methods in virus diagnostics: From ELISA to next generation sequencing. Virus Research 186: 20-31. |
|
|
Boonham N, Pe'rez LG, Me'ndez MS, Peralta EL, Blockley A, Walsh K, Barker I, Mumford RA (2004). Development of a real-time RT-PCR assay for the detection of potato spindle tuber viroid. Journal of Virological Methods 116:139146. |
|
|
Bostan H, Nie X, Singh RP (2004). An RT-PCR primer pair for the detection of pospiviroid and its application in surveying ornamental plants for viroids. Journal of Virological Methods 116:189193. |
|
|
Botermans M, van de Vossenberg BT, Verhoeven JThJ, Roenhorst JW, Hooftman M, Dekter R, Meekes ETM (2013). Development and validation of a real-time RT-PCR assay for generic detections of pospiviroids. Journal of Virological Methods 187:4350. |
|
|
CABI/EPPO (Centre for Agriculture and Bioscience International/ European and Mediterranean Plant Protection Organization) (2019). Potato Virus Y (Potato mottle). Distribution Maps of Plant Diseases. |
|
|
CABI/EPPO (Centre for Agriculture and Bioscience International/European and Mediterranean Plant Protection Organisation) (2014). Potato Spindle Tuber Viroid. Distribution Maps of Plant Diseases No. 729. CABI Head Office, Wallingford, UK. |
|
|
Candresse T, Cambra M, Dallot S, Lanneau M, Asensio M, Gorris MT, Revers F, Macquaire G, Olmos A, Boscia D, Quiot JB, Dunez J (1998). Comparison of monoclonal antibodies and PCR assays for the typing of isolates belonging to the D and M serotypes of plum pox virus. Phytopathology 88:198-204. |
|
|
Capobianchi MR, Giombini E, Rozera G (2013). Next-generation sequencing technology in clinical virology. Clinical Microbiology and Infection 19:15-22. |
|
|
Coetzee B, Freeborough MJ, Maree HJ, Celton JM, Rees DJ, Burger JT (2010). Deep sequencing analysis of viruses infecting grapevines: virome of a vineyard. Virology 400(2):157-63. |
|
|
Coudray-Meunier C, Fraisse A, Martin-Latil S, Delannoy S, Fach P, Perelle S (2016). A novel high-throughput method for molecular detection of human pathogenic viruses using a nanofluidic real-time PCR system. PLoS ONE 11: 1-17. |
|
|
Cox-Foster DL, Conlan S, Holmes EC, Palacios G, Evans JD, Moran NA, Quan P, Briese T, Hornig M, Geiser DM, Martinson V, van Engelsdorp D, Kalkstein AL, Drysdale A, Hui J, Zhai J, Cui L, Hutchison SK, Simons JF, Egholm M, Pettis JS, Lipkin WI (2007). A metagenomic survey of microbes in honey bee colony collapse disorder'. Science 318: 283-287. |
|
|
Craig G, Webster, Stephen J, Wylie, Michael GK, Jones (2004). Diagnosis of plant viral pathogens. 86(12):10604-1607 Available at |
|
|
Denner FDN, Venter SL, Niederwieser JG (2012). Guide to potato production in South Africa. Pretoria, SA: ARC-Roodeplaat Vegetable and Ornamental Plant Institute. |
|
|
Dodds JA, Morris TJ, Jordan RL (1984). Plant viral double-stranded RNA'. Annual Review of Phytopathology 22:151-168. |
|
|
Dong M, Fisher C, Anez G, Rios M, Nakhasi HL, Hobson JP, Beanan M, Hockman D, Grigorenko E, Duncan R (2016). Standardized methods to generate mock (spiked) clinical specimens by spiking blood or plasma with cultured pathogens. Journal of Applied Microbiology 120:1119-1129. |
|
|
Edwards RA, Rohwer F (2005). Opinion: viral metagenomics. Nature reviews. Microbiology 3: 504. |
|
|
Espach A (2015). Final report. The validation of virus-specific real-time RT-PCR assays for commercial use in the potato industry. |
|
|
Espach Y, Maree HJ, Burger JT (2012). Complete Genome of a Novel Endornavirus Assembled from Next Generation Sequence Data. Journal of Virology 86:13142. |
|
|
Evangelista C (2013). Viruses Occurring in Potatoes (Solanum tuberosum) in Mbeya Region, Tanzania. University of Helsinki. |
|
|
Department of Agricultural Sciences. Plant Production Science/ Plant Pathology January, 2013. |
|
|
FAOSTAT (2011). Available at View |
|
|
FAOSTAT (2019). Available at View |
|
|
Giampetruzzia A, Roumia V, Robertoa R, Malossinib U, Yoshikawac N, Nottea PL, Terlizzi F, Credid R, Saldarelli P (2011). A new grapevine virus discovered by deep sequencing of virus-and viroid-derived small RNAs in Cv Pinot gris. Virus Research 163:262-8. |
|
|
Gildemacher PR (2012). Innovation in seed potato systems in Eastern Africa.PhD Thesis Wageningen University, Netherlands. pp. 12-13. Available at View |
|
|
Gildemacher PR, Paul D, Baker I, Kaguongo W, Woldegiorgis G, Wagoire WW, Wakahiu M, Leeuwis C, Struik PC (2009). A description of seed potato systems in Kenya, Uganda and Ethiopia. Potato Research 86:373-382. |
|
|
Goodwin S, McPherson JD, Richard McCombie W (2016). Coming of age: Ten years of next-generation sequencing technologies. Nature Reviews Genetics 17:333-351. |
|
|
Gould BA, Stinchcombe JR (2017). Population genomic scans suggest novel genes underlie convergent flowering time evolution in the introduced range of Arabidopsis thaliana. Molecular Ecology 26:92-106. |
|
|
Grothaus DG, Bandla M, Currier T, Giroux R, Jenkins RG, Lipp M, Shan G, Stave JW, Pantella V (2006). Immunoassay as an analytical tool in agricultural biotechnology. Journal of AOAC International 89:913-928. |
|
|
Harris TD, Buzby PR, Babcock H, Beer E, Bowers J, Braslavsky I, Causey M, Colonell J, DiMeo J, Efcavitch JW, Giladi E (2008). Single molecule DNA sequencing of a viral genome. Science 320:106-9. |
|
|
Hockman D, Dong M, Zheng H, Kumar S, Huff MD, Grigorenko E, Beanan M, Duncan R (2017). Comparison of multiplex PCR hybridization-based and singleplex real-time PCR-based assays for detection of low prevalence pathogens in spiked samples. Journal of Microbiological Methods. Elsevier B.V 132:76-82. |
|
|
Hossain B, Nasir IA, Tabassum B, Husnain T (2013). Molecular characterization, cloning and sequencing of coat protein gene of a Pakistani Potato leaf roll virus isolate and its phylogenetic analysis. African Journal of Biotechnology 12:1196-1202. |
|
|
Hühnlein A, Schubert J, Zahn V, Thieme T (2016). Examination of an isolate of Potato leafroll virus that does not induce visible symptoms in the greenhouse. European Journal of Plant Pathology 145: 829-845. |
|
|
Huse SM, Huber JA, Morrison HG, Sogin ML, Welch DM (2007). Accuracy and quality of massively parallel DNA pyrosequencing. Genome Biology 8:R143. |
|
|
Hwang YT, Kalischuk M, Fusaro AF, Waterhouse PM, Kawchuk L (2013). Small RNA sequencing of Potato leafroll virus-infected plants reveals an additional subgenomic RNA encoding a sequence-specific RNA-binding protein. Virology 438:61-69. |
|
|
Ibaba JD, Gubba A (2011). Diversity of potato virus Y isolates infecting solanaceous vegetables in the province of KwaZulu. Natal in the Republic of South Africa. Crop Protection 30:1404-1408. |
|
|
IPBO (International Plant Biotechnology Outreach) (2019). Potato in Africa. RE, Silvia Travella, (IPBO/VIB) D/2019/12.267/3. Technologie park 19 9052 Ghent Belgium Tel. + 32 9 292 80 44 |
|
|
John O, Kiarie N, Solomon S, Muthoni J, Otieno SP (2013). Potato virus Y (PVY) and potato virus X (PVX) resistance breeding in Kenya: Applicability of conventional approaches. Agriculture and Biology Journal of North America 4:398-405. |
|
|
Kabira JN, Wakahihu M, Wagoire W, Gildemacher P, Lemaga B (2006). Guidelines for production of healthy seed potatoes in East and Central Africa. Edited by Lusike Wasilwa, Kenya Agricultural Research Institute, Nairobi, Kenya. pp. 1-28. Available at |
|
|
Kreuze Jan F, Perez A, Untiveros M, Quispe D, Fuentes S (2009). Complete viral genome sequence and discovery of novel viruses by deep sequencing of small RNAs: A generic method for diagnosis, discovery and sequencing of viruses. Virology 388:1-7. |
|
|
Kreuze JF (2014). siRNA deep sequencing and assembly: Piecing Together Viral Infections in Detection and diagnostics of plant pathogens. Springer Netherlands, pp. 21-38. |
|
|
Kumar R, Jeevalatha A, Baswaraj R, Kumar R, Sharma S, Nagesh M (2017). A multiplex RT-PCR assay for simultaneous detection of five viruses in potato. Journal of Plant Pathology 99:37-45. |
|
|
Kunta M, da Graca JV, Malik NSA, Louzada ES, Setamou M (2014). Quantitative distribution of "Candidatus Liberibacter asiaticus" in the aerial parts of the Huanglongbing-infected citrus trees in Texas. HortScience 49:65-68. |
|
|
Lacomme C, Glais L, Bellstedt DU, Dupuis B, Karasev A, Jacquot E (2017). Potato virus Y: biodiversity, pathogenicity, epidemiology and management. Springer International Publishing. |
|
|
Lacroix C, Renner K, Cole E, Seabloom EW, Borer ET, Malmstrom CM (2016). Methodological guidelines for accurate detection of viruses in wild plant species. Applied and Environmental Microbiology 82:1966-1975. |
|
|
Laehnemann D, Borkhardt A, McHardy AC (2015). Denoising DNA deep sequencing data-high-throughput sequencing errors and their correction. Brief Bioinformatics 17:154-79. |
|
|
Larbi I, Djilani-Khouadja F, Khamassy N, Fakhfakh H (2012). Potato virus surveys and wide spread of recombinant PVYNTN variant in Central Tunisia. African Journal of Microbiology Research 6(9):2109-2115. |
|
|
Laver T, Harrisona J, O'Neill PA, Moorea K, Farbosa A, Paszkiewicz K, Studholme DJ (2015). Studholme. Assessing the performance of the oxford nanopore technologies MinION. Biomolecular Detection and Quantification 3:1-8. |
|
|
Lezan W (2017). An investigation into the Potato leafroll virus problem in the Sandveld region, South Africa. |
|
|
Li Z, Guo B, Yang J, Herczeg G, Gonda A, Balazs G, Meril€a J (2017). Deciphering the genomic architecture of the stickleback brain with a novel multilocus gene-mapping approach. Molecular Ecology 26:1557-1575. |
|
|
Lotos L, Olmos A, Orfanidou C, Efthimiou K, Avgelis A, Katis N, Maliogka VI (2017). Insights into the etiology of Polerovirus-induced pepper yellows disease. Phytopathology 107. |
|
|
Machangi JM, Olubayo FM, Njeru RW, Nderitu JH, El-Bedewy R, Yobera DM, Aura JA (2004). Occurrence of four major potato viruses in three main potato growing areas in Kenya. 6th Trienieal Conference, April 2004. African Potato Association (APA), pp. 273-281. |
|
|
Mardis ER (2008). Next-generation DNA sequencing methods. Annual Review of Genomics and Human Genetics 9:387-402. |
|
|
Maree HJ, Pirie MD, Oosthuizen K, Bester R, Jasper D, Rees G, Burger JT (2015). Phylogenomic analysis reveals deep divergence and recombination in an economically important grapevine virus. PLoS ONE 10: 1-19. |
|
|
Margulies M, Egholm M, Altman WE, Attiya S, Bader JS, Berka J, Braverman MS, Chen YJ, Chen Z, Dewell SB (2005). Genome sequencing in microfabricated high-density picolitre reactors. Nature 437:376-380. |
|
|
Marina B, Henryk C, Ahmed H (2014). Historical Perspective, Development and Applications of Next-Generation Sequencing in Plant Virology. Viruses 6:106-136. |
|
|
Matthews REF (1991). Plant Virology (Third Edition), Academic Press, 1991. Page xix, ISBN 9780124805538. |
|
|
Martin IC (2012). Molecular Diagnostics in Plant Disease Diagnostic Clinics. What's the Status? Fungal Genomics and Biology 2:1. |
|
|
Marz M, Beerenwinkel N, Drosten C, Fricke M, Frishman D, Hofacker IL, Hoffmann D, Middendorf M, Rattei T, Stadler PF, Töpfer A (2014). Challenges in RNA virus bioinformatics. Bioinformatics 30:1793-1799. |
|
|
McCoy RC, Taylor RW, Blauwkamp TA, Kelley JL, Kertesz M, Dmitry Pushkarev, Dmitri AP, Anna-Sophie F-L (2014). Illumina TruSeq synthetic long-reads empower de novo assembly and resolve complex, highly-repetitive transposable elements. PLoS ONE 9:e106689. |
|
|
Mikheyev AS, Tin MMY (2014). A first look at the oxford nanopore MinION sequencer. Molecular Ecology Resources 14:1097-1102. |
|
|
Mokili JL, Rohwer F, Dutilh BE (2012). Metagenomics and future perspectives in virus discovery. Current Opinion in Virology 2:63-77. |
|
|
Muthomi JW, Kinyungu TN, Nderitu JH, Olubayo FM (2011). Incidence of aphid- Transmitted viruses in farmer-produced seed potato tubers in Kenya. African Journal of Horticultural Science 5:18-25. |
|
|
Muthomi JW, Nyaga JN, Olubayo FN, Nderitu JH, Kabira JN, Kiretai SM, Aura JA, Wakahiu M (2009). Incidence of aphid transmitted viruses in farmer based seed potato production in Kenya. Asian Journal of Plant Sciences 8:166-171. |
|
|
Myllykangas S, Buenrostro J, Ji HP (2012). Overview of sequencing technology platforms, bioinformatics for high throughput sequencing. |
|
|
Ng TFF, Duffy S, Polston JE, Bixby E, Vallad GE (2011). Exploring the diversity of plant DNA viruses and their satellites using vector enabled metagenomics on whiteflies. PLoS ONE 6(4):e19050. |
|
|
Njukeng PA, Chewachong GM, Sakwe P, Chofong G, Nkeabeng LW, Demo P, Njualem KD (2013). Prevalence of Six Viruses in Potato Seed Tubers Produced in Informal Seed System in the North West Region of Cameroon. Cameroon Journal of Experimental Biology 09(01):44-49. |
|
|
Notomi T, Okayama H, Masubuchi H, Yonekawa T, Watanabe K, Nobuyuki A, Tetsu H (2000). Loop-mediated isothermal amplification of DNA. Nucleic Acids Res 28:E63. |
|
|
Nyamwamu PA, Mukoy B, Osogo AK, Omuse CN, Ajanga S, Were HK (2014). Distribution and Biological Characterization of Potato Virus Y in Kenya. Journal of Agri-Food and Applied Sciences 2(9):258-264. |
|
|
Nyaboga EN, Ateka EM, Bulimo WD (2008). Serological detection of virus diseases of sweet potato in Kenya. Journal of Applied Biosciences 7:222-229. |
|
|
Okeyo OG (2017). Response of Potato Genotypes to Virus infections and effectiveness of Positive selection in management of seed borne potato viruses. UoNbi.erepository. |
|
|
Olubayo F, Kibaru A, Nderitu J, Njeru R, Kasina M (2010). Management of aphids and their vectored diseases on seed potatoes in Kenya using synthetic insecticides, mineral oil and plant extract. Journal of Innovation and Development Strategy 4(2):1-5. |
|
|
Pallett DW, Ho T, Cooper I, Wang H (2010). Detection of cereal yellow dwarf virus using small interfering RNAs and enhanced infection rate with cocksfoot streak virus in wild cocksfoot grass (Dactylis glomerata). Journal of Virological Methods 168:223-7. |
|
|
Pereira AM, Lister RM (1989). Variations in virus content among individual leaves of cereal plants infected with Barley yellow dwarf virus. Phytopathology 79:1348-1353. |
|
|
Quan P, Briese T, Palacios G, Lipkin WI (2008). Rapid sequence-based diagnosis of viral infection'. Antiviral Research 79:1-5. |
|
|
Prabha K, Baranwal VK, RK Jain (2013). Applications of Next Generation High Throughput Sequencing Technologies in Characterization, Discovery and Molecular Interaction of Plant Vi-ruses. Indian Journal Virology. |
|
|
Rashed A, Workneh F, Paetzold L, Gray J, Rush CM (2014). Zebra chip disease development in relation to plant age and time of "Candidatus Liberibacter solanacearum" infection. Plant Disease 98:24-31. |
|
|
Rhoads A, Au KF (2015). PacBio sequencing and its applications. Genomics, Proteomics and Bioinformatics 13:178-289. |
|
|
Roossinck MJ, Saha P, Wiley GB, Quan J, White JD, Lai H, Chavarrıoa F, Shen G, Roe BA (2010). Ecogenomics: Using massively parallel pyrosequencing to understand virus ecology. Molecular Ecology 19(Suppl. 1):81-88. |
|
|
Rotheberg JM, Hinz W, Rearrick TM, Schultz J, Mileski W, Davey M, Leamon JH, Johnson K, Milgrew MJ, Edwards M, Hoon J (2011). An integrated semiconductor device enabling non-optical genome sequencing. Nature 475: 348-52. |
|
|
Safenkova IV, Pankratova GK, Zaitsev IA, Varitsev YA, Vengerov YY, Zherdev AV, Dzantiev BB (2016). Multiarray on a test strip (MATS): rapid multiplex immunodetection of priority potato pathogens. Analytical and Bioanalytical Chemistry 408:6009-6017. |
|
|
Sanchez-Navarro JA, Cañizares MC, Cano EA, Pallás V (2007). Plant tissue distribution and chemical inactivation of six carnation viruses. Crop Protection 26:1049-1054. |
|
|
Schadt EE, Turner S, Kasarskis A (2010). A window into third-generation sequencing. Human Molecular Genetics19:R227-240. |
|
|
Schulte-Geldermann E, Gildemacher PR, Struik PC (2012). Improving seed health and seed performance by positive selection in three Kenyan potato varieties. American Journal of Potato Research 89(6):429-437. |
|
|
Seguin J, Rajeswaran R, Malpica-López N, Martin RR, Kasschau K, Dolja VV, Otten P, Farinelli L, Pooggin MM (2014). De novo reconstruction of consensus master genomes of plant RNA and DNA viruses from siRNAs. PLoS ONE 9:1-8. |
|
|
Shamloul AM, Hadidi AF, Zhu SF, Singh RP, Sagredo B (1997). Sensitive detection of potato spindle tuber viroid using RT-PCR and identification of a viroid variant naturally infecting pepino plants. Canadian Journal of Plant Pathology 19:8996. |
|
|
Shendure J, Ji H (2008). Next-generation DNA sequencing. Nature Biotechnology 26:135-1145. |
|
|
Snyder M, Du J, Gerstein M (2010). Personal genome sequencing: Current approaches and challenges. Genes and Development 24:423-431. |
|
|
Standage DS, Berens AJ, Glastad KM, Severin AJ, Brendel VP, Toth AL (2016). Genome, transcriptome and methylome sequencing of a primitively eusocial wasp reveal a greatly reduced DNA methylation system in a social insect. Molecular Ecology 25:1769-1784. |
|
|
Studholme DJ, Glover RH, Boonham N (2011). Application of high throughput DNA sequencing in phytopathology. Annual Review of Phytopathology 49:87-105. |
|
|
Thomas T, Gilbert J, Meyer F (2012). Metagenomics - A guide from sampling to data analysis'. Microbial Informatics and Experimentation 2:1-12. |
|
|
Verhoeven JThJ, Jansen CCC, Willemen TM, Kox LFF, Owens RA, Roenhorst JW 2004). Natural infections of tomato by citrus exocortis viroid, columnea latent viroid, potato spindle tuber viroid and tomato chlorotic dwarf viroid. European Journal of Plant Pathology 110:823831. |
|
|
Wangai A, Lelgut D (2013). Status of potato viruses in Africa. Kenya Agricultural Research Institute, NPBRC, P. O. Box Njoro, Kenya (KALRO). |
|
|
Were HK, Kabira JN, Kinyua ZM, Olubayo FM, Karinga JK, Aura J, Lees AK, Cowan GH, Torrance L (2014). Occurrence and Distribution of Potato Pests and Diseases in Kenya. Potato Research. |
|
|
Were HK, Kabira JN, Kinyua ZM, Olubayo FM, Karinga JK, Aura J, Torrance L (2013). Occurrence and Distribution of Potato Pests and Diseases in Kenya. Potato Research 56(4):325-342. |
|
|
Wiets GR (2013). An investigation of prevalence and the detection and race identification of South African potato viruses. Stellenbosch University View |
|
|
Wu Q, Ding S, Zhang Y, Zhu S (2015). Identification of viruses and viroids by next generation sequencing and homology-dependent and homology-independent algorithms. Annual Review of Phytopathology 53:425-444. |
|
|
Wylie SJ, Jones MGK (2011). The complete genome sequence of a Passion fruit woodiness virus isolate from Australia determined using deep sequencing, and its relationship to other potyviruses. Archives of Virology 156:479-482. |
|
|
Yanagisawa H, Tomita R, Katsu K, Uehara T, Atsumi G, Tateda C, Kobayashi K, Sekine K (2016). Combined DECS analysis and next-generation sequencing enable efficient detection of novel plant RNA viruses. Viruses 70:1-11. |
|
|
Zhang W, Zhang Z, Fan G, Gao Y, Wen J, Bai Y, Qiu C, Zhang S, Shen Y, Meng X (2017). Development and application of a universal and simplified multiplex RT-PCR assay to detect five potato viruses. Journal of General Plant Pathology 83:1-13. |
|
|
Zheng Y, Gao S, Padmanabhan C, Li R, Galvez M, Gutierrez D, Fuentes S, Ling K, Kreuze JF, Fei Z (2017). VirusDetect. An automated pipeline for efficient virus discovery using deep sequencing of small RNAs. Virology. Elsevier 500:130-138. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


