Full Length Research Paper

ABSTRACT
Griffonia simplicifolia is a Caesalpiniaceae legume that grows from Liberia to the Democratic Republic of Congo. The leaves are widely used in Togo for their nutritional value. On the other hand it is most known for the richness in 5-Hydroxy-L-tryptophan (L-5-HTP) of its seeds. Farmers collect significant amounts of fodder directly from Togo's forest formations, resulting in a scarcity of this species, which is classed as non-timber forest products. Massive anthropization, in combination with the low germinative potential of G. simplicifolia seeds, poses a threat to the species' survival and availability. It is critical to promote and disseminate G. simplicifolia's culture in order to help save the species, which is presently critically endangered. It is in this perspective that the study of germination and conservation of the germination capacity of G. simplicifolia seeds was conducted. The ecotype being studied was that of Lomé, which was harvested on the campus of the University of Lomé. Light, integuments, time, and storage temperature were all factors considered when studying germination. To optimize the germination speed, it is essential to remove the seed coat and germinate the seeds with light in the greenhouse as in vitro. The cold helps to preserve the germination capacity of seeds, which deteriorates quickly in warm environments.
Key words: Griffonia simplicifolia, L-5-HTP, in vivo and in vitro germination.
INTRODUCTION
Griffonia simplicifolia (Caesalpiniaceae) is a liana native to African which can be found in Ghana, Ivory Coast, and southwest Togo (Hutchinson and Dalziel, 1954). Griffonia is a genus of shrub legumes, and G. simplicifolia is a climbing shrub with a flexible trunk. It has been attracting scientific attention since the 1960s because of its action on mood, sleep and nervous tension (Bell et al ., 1976; Kumar et al., 2010; Zavarise and Dalla, 2017. This observed efficacy is due to one substance: 5-Hydroxy-L-tryptophan (L-5-HTP) synthesized and stored mainly in its seeds. The seed has been reported to contain 6 to 14% in mass of L-5-HTP (Bosch, 2008; Addotey and, Adosraku, 2016). Nowadays, the seed of this climbing shrub is used in the treatment of fibromyalgia, headaches, insomnia, depression and for stomach problems. In fact, G. simplicifolia seed is the richest known plant organ for L-5-HTP and is therefore currently the main source of L-5-HTP used for industrial purposes (Lemaire and Adosraku, 2002; Asiamah et al., 2006). However, it is now possible to synthesize L-5-HTP by genetically transformed Escherichia coli cultures (Mora-Villalobos and Zeng, 2018). Formulations containing G. simplicifolia are available over the counter and are highly appreciated by practitioners of natural and alternative medicine. However, the scientific exploration of the plant within the framework of secondary metabolites, notably lectins (Maddox et al., 1982; Nyarko et al., 2019) and its therapeutic efficacy for psychological and psychiatric pathologies have been widely documented (Methta et al., 2015).
The exploitation of the seeds of G. simplicifolia, non-timber forest products, provides significant income to local communities in the western African sub-region (FAO, 1999). In Togo, G. simplicifolia is specifically exploited and marketed as a fodder plant, whereas neighboring Côte d'Ivoire and Ghana are more likely to be countries engaged in international trade in seed raw materials.
The over exploitation of G. simplicifolia in Togo, for fodder and also incursions for seed collection from neighboring countries, is gradually causing its extinction in the country's forests. This species, which was harvested until the 1980s in the immediate vicinity of the capital, Lomé, can now only be found beyond a radius of 20 to 30 km (Kadévi, 2001). Regrowth of G. simplicifolia is hampered by difficult germination related to poor and too brief retention of seed germinability (Bhatla and Lal, 2018).
Several studies have focused on the use of secondary metabolites extracted from G. simplicifolia seeds and on the therapeutic effects of these extracts (Birdsall, 1998; Esposito et al., 2015; Kakou et al., 2018). However, few scientific studies have been done on the aspect of germination and genetic improvement. Improving various techniques of vegetative multiplication is necessary to enhance the value of the crop and to mitigate the problems of overexploitation of forest resources. We carry out a preliminary study of the generative propagation of the plant.
MATERIALS AND METHODS
Plant
The ecotype studied in this work was the one from the Lomé area. The seeds of G. simplicifolia were harvested every eight days for a period less than twelve months on the campus of the University of Lomé, Togo, during the three months of seed production (January to March). Seeds of 16 to 18 mm, were collected after the mature pods had burst open, the harvested seeds were graded and stored in transparent glass bottles. Seeds harvested over an eight day period were divided into lots.
Germination conditions in the greenhouse
The germination study of G. simplicifolia seeds was carried out in large Petri dishes (r = 14 cm) in the greenhouse, at a temperature of 28 ± 2°C during the day and 25 ± 1°C at night, a relative humidity of 60%, lighting with a photoperiod of 16 h of lght and 08 h of darkness and a light intensity of 120 µE. m-².s-1. The peeled or not peeled seeds were quickly introduced (10 to 15 s) in a solution of pure yellow Betadine® (Povidone iodine: 10 g/100 ml) at 2% (v/v) before undergoing subsequent treatments and being put to germinate. Ten seeds were placed per Petri dish and six replicates were performed per treatment.
To study the influence of the integuments on the germination of G. simplicifolia seeds, the protocol adopted is summarized in Table 1.

To explore light influence, 12 replicates (6 times, 2 replicates) of 10 fully shelled seeds were used. The control seeds were kept in the same conditions as described earlier. The remaining 12 replicates (6 repetitions times 2) of 10 seeds were set in continuous darkness under the same conditions.
For all these treatments different germination indicators as it will be seen later, have been evaluated.
The cultures were observed daily and the germinated seeds were counted every 24 h for eight days. Seeds were considered to have germinated upon radical emergence ≥ 0.5 mm. The following parameters were recorded for each protocol (Joshi et al., 2016):
(1) Final germination percentage (FGP %) (Gashi et al., 2012) (Equation 1):
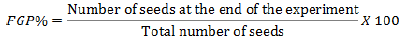
(2) Mean germination time (MGT) in day (Equation 2):
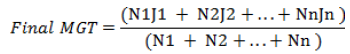
where N1 = number of seeds germinated the first day, N2 = the cumulative number of seeds germinated the second day, Nn = the cumulative number of seeds germinated the last day, j1 = first day = 1, j2 = second day = 2, and jn = nth day = n.
(3) Germination rate index (GRI %) (Esechie, 1994) (Equation 3):
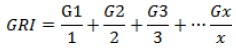
where G1, G2, G3, …, Gx were germination percentage x100 at days 1, 2, 3, …x days after sowing, respectively.
(4) Corrected germination rate index (CGRI) (Gashi et al., 2012) (Equation 4):
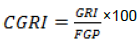
(5) Germination rate (GR) (Rajabi and Poustini, 2005) (Equation 5):
GR = Number of seeds germinated per day
(6) Percentage germination rate (Shah et al., 2015) (Equation 6):
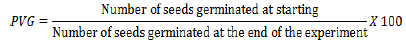
(7) Mean daily germination rate (MDG) (Kafi and Goldani, 2001) (Equation 7):
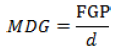
where d = number of days to the end of germination
(8) Germination value1 (GV1) (Djavanshir and Pourbeik, 1976) (Equation 8):
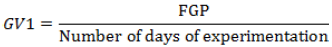
(9) Germination value (GV2) (Czabator, 1962) (Equation 9):
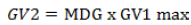
(10) Coefficient of germination velocity (CVG %) (Elyasi et al., 2014) (Equation 10):
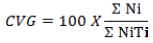
where Ni = number of seeds germinated per day, Ti = Number of days since the beginning of the experiment.
In vitro germination conditions
In vitro germination studies were carried out on Murashige and Skoog medium (Murashige and Skoog, 1962) supplemented with 30 g/L of sucrose, solidified with 8 g/L of agar, or cast liquid on washed and disinfected silica sand. Seeds aged between 8 and 16 days, fully dehulled and distributed at a rate of 1 seed per tube and sixty (60) replicates, were used with a photoperiod of 16 h of light intensity 120 µE. m-².s-1, at a temperature of 24 ± 2°C during the day and 24 ± 3° C at night. Before being sown, the completely shelled seeds were disinfected outside the hood by soaking in a solution of pure yellow Betadine® (Povidone iodine: 10 g/100 ml) at 2% (v/v) for 30 min, followed by a rinse with sterile water. Under laminar flow hood, the seeds undergo two disinfection protocols called in vitro 1 (for the 2 min soaking time) and in vitro 2 (for the 5 min soaking time). They were then soaked with shaking successively in water with 0.2% (v/v) Tween 80 added for 5 min, and in a solution of pure yellow Betadine® (Povidone iodine: 10 g/100 ml) at 10% (v/v) for 2 to 5 min and finally in a solution of sodium hypochlorite at 50% (v/v) during 2 or 5 min followed by six rinsing with sterile water.
Conditions for preserving the viability and germinative capacity of seeds
Niewenhuis and Nieuwelink (2005) have shown a correlation between cotyledon color and seed viability conservation. Seeds of G. simplicifolia that are likely to germinate are those that have retained their yellowish color after dehulling. The yellowish color of the seeds is thus chosen as a criterion for seed viability and germination is carried out on a regular basis.
For the viability study, the batches of seed representing eight days of harvest and stored in transparent glass bottles, were divided into four distinct batches without any pre-treatment and stored at four different temperatures: 30 ± 4°C to 60% relative humidity; 24 ± 1°C to 60%; 7 ± 2°C to 44% relative humidity; and -11 ± 3°C to 22% relative humidity, respectively the ambient, laboratory, refrigerator and freezer compartment temperatures. Every week for three to four months, a batch of six times ten seeds (6 × 10 seeds) per storage temperature were dehulled, examined for seed coloration and then germinated under the predefined conditions. Two germination capacities were then defined: the real germination capacity and the estimated one.
Statistical processing
For the germination data, a bivariate analysis method and FGP variables were used with a Chi-square test to determine whether the pre-treatment method used for the seeds influenced the FGP value. To determine an initial classification of the methods, a pairwise comparison test of the germination percentage was carried out. Then a multivariate analysis using Principal Component Analysis (PCA) was performed on the data set.
For the germination retention data, the method of least squares was used, approximation polynomials of the data were determined and the comparison of the averages was done by the Welch (1947) test.
RESULTS
Germination study of G. simplicifolia
Influence of teguments
The germination of G. simplicifolia seeds has the peculiarity of causing the opening and greening of the cotyledons prior to the exit of the radicle. The observed greening will persist throughout the germination period and the cotyledons persist. Under the experimental conditions applied here, 10% of the control seeds start germinating on day 3, germination stops after 6 days, with a maximum capacity of 70% and an average germination time of 5.13 days (Figure 1). Germination kinetics was discontinuous, with 1 seed on day 3, 2 on day 4, 1 on day 5 and 3 on day 6, so the average germination rate was just 1.16 seeds per day with a peak on day 3.
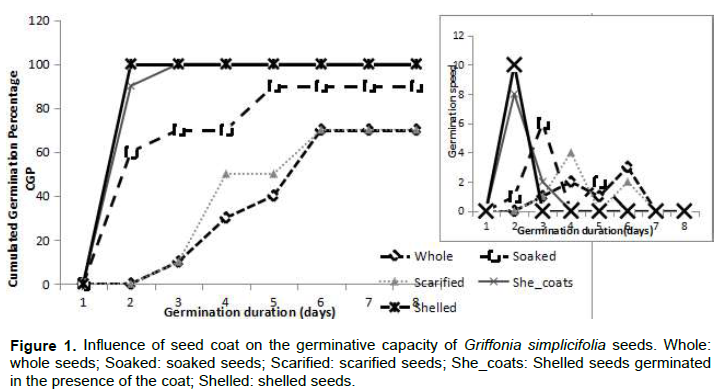
Scarification of the seeds improves the progress of germination with 1 seed on day 3, 4 on day 4 and 2 on day 6 without increasing the average speed which remains at 1.16 seeds per day and the final cumulative percentage of germination which remains at 70%. However, the average germination time drops significantly by one day to 4 days.
The preliminary soaking of the whole seeds, not shelled, allows them to start germinating at 60% from day 2. This germination was spread over five days. At the end of the experiment, a maximum rate of 90% and an average germination time of 3.65 days were achieved. The remaining 10% of the seeds were all necrotic. Pre-soaking not only improved the germination rate, but also shortened the average germination time (Table 2).
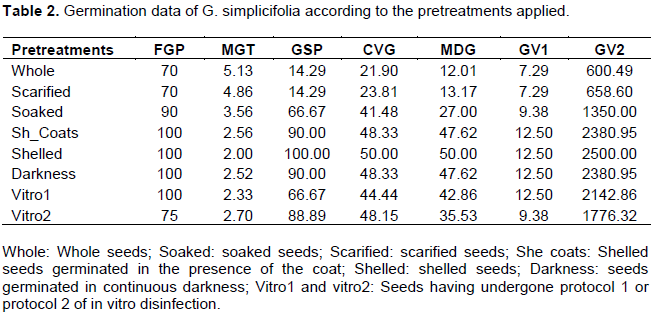
When the seeds were peeled and germinated in the presence of the teguments, germination started as in the case of pre-soaked whole seeds from day 2, at a capacity of 80% and from day 3, the final germination percentage reached the maximum of 100% (Figure 1). The average germination time was 2.56 days. Peeled seeds germinated in the absence of the seed coat all germinate from day 2, the average germination time is 2 days. When the seeds were shelled, the germination speed showed a single maximum peak from the second day with 100% of the seeds germinating in the absence of the seed coat and 80% of the seeds germinating in the absence of the seed coat (Table 2). In general, and for all experiments, a fungal contamination of the seed coat has been observed as early as three days after the start of germination, which continues to develop. The influence of the seed coat on the germination of G. simplicifolia seeds was mainly due to contamination, the germination percentages recorded were essentially dependent on the germination speed of the seeds. When the average germination time exceeds 3 days, the seeds were contaminated and therefore unable to germinate.
Influence of light
The completely shelled seeds of G. simplicifolia germinated both in complete darkness and in day/night alternation with a maximum germinative capacity of 100%. However, even though the average germination time was 2.52 days in the dark, the significant result was that the cotyledons do not turn green in the dark. They remained yellowish and there was a beginning of necrosis around the seeds. On the other hand, in light, two days were enough to reach the maximum germination capacity. All seeds germinated with green, slightly open cotyledons and no necrotic parts were observed.
Germination kinetics were almost identical whether the seeds were in the light or in the dark, the maximum speeds were also the same, with 9 seeds per day in the dark and 10 seeds per day in the light. The average times were also not significantly different. In conclusion, the remarkable difference when comparing the germination of G. simplicifolia seeds in darkness and in day/night alternation is the beginning of necrosis of the pale yellow cotyledons of the shelled seeds germinated in complete darkness. Light therefore has no direct influence on the germination of G. simplicifolia seeds.
Germination of G. simplicifolia in vitro
The protocols used for seed disinfection yielded good results and maximum germination for the former, although 8% of seeds were still found to be contaminated, mainly by fungal contamination. As for the second protocol, complete disinfection was achieved; however, the germination percentage was 25% lower than the percentage obtained with protocol 1, even though the seeds germinated. For seeds that underwent the protocol germination takes place as quickly in vitro as in greenhouse conditions, since the same average germination time was observed. A browning of the culture medium was observed when the cotyledons were affected by disinfectants. The same browning was observed around the roots and intensified with root growth on the MS agar medium. Similarly, when agar-agar was replaced by silica sand, germination took place normally as on agar and browning was no longer observed. The average germination time was the same in vitro as in the greenhouse.
Global analysis of the results of germination of G. simplicifolia
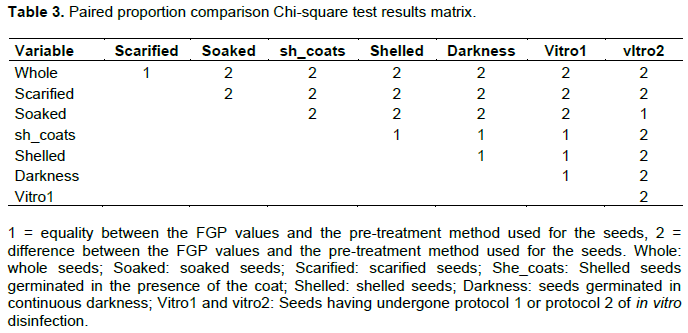
The Chi-square test statistic was 216 with high significance (p-value < 2.2.10-16). It can therefore be concluded that seed pre-treatments have a highly significant influence on the final percentage of germination (FGP). The results of the paired proportion comparison test also allow us to affirm that the observed differences are really significant to the point of clearly identifying three distinct classes: the weakest class 1, consisting of whole and scarified seeds, the average class 2, consisting of soaked seeds and those subjected to in vitro disinfection protocol 2 and finally the strongest class 3, consisting of shelled with coats, shelled, light seeds and those subjected to the in vitro disinfection protocol 1 (Table 3).
The results of the principal components analysis (PCA) with highly correlated variables, confirm the classification made by the comparison test. The main axis (axis 1) distinguishes seed types with high FGP, GV1, GV2, MDG, CVG, GR values and low MGT values compared to the others (Figure 2). Figure 2 shows the three classes: low, medium and high performance. The high-performance class therefore combines high FGP and speed values with a reduced average germination time of only 2 days on average.
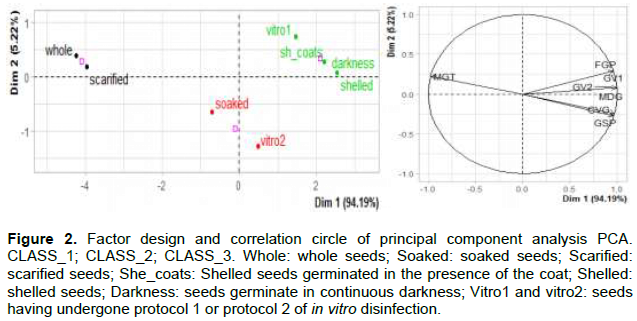
Conservation study of seeds germinative capacity
Conservation analysis of G. simplicifolia seeds germination capacity according to conservation temperature
Seeds stored at room temperature, that is, 30 ± 4°C with a relative humidity of 60%, begin to be darkened as early as the fourth week after harvest. 50% of the seeds necrotized for the tests at 4 weeks and after 2 months of storage under the same conditions, the seeds were no longer viable. After germination, 67% of the seeds were necrotic on day 2 and all seeds were necrotic on day 4 (Figure 3D).
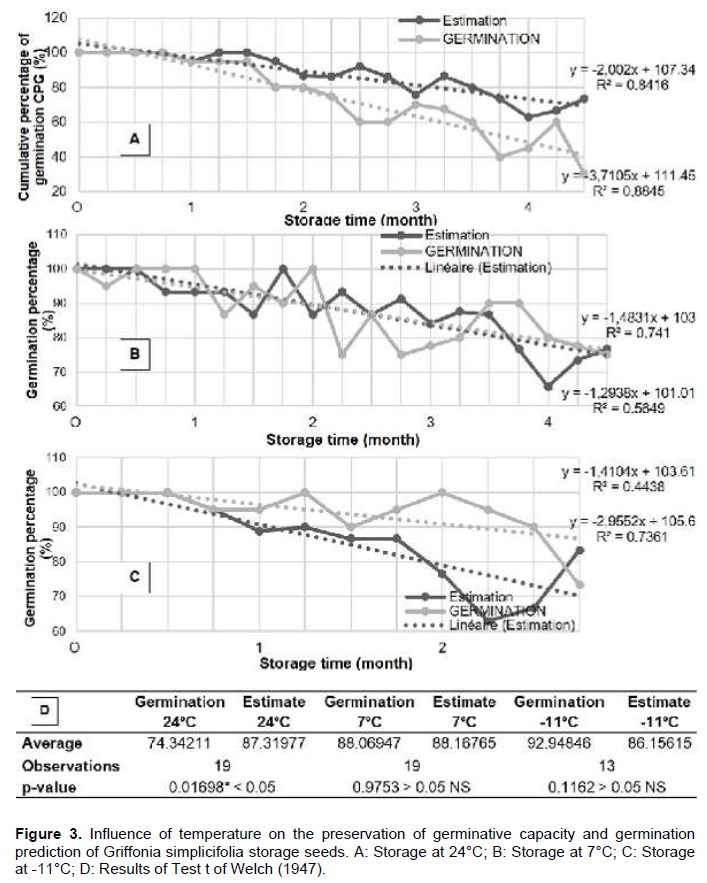
In general, it can be seen that for the three storage temperatures chosen for the study: 24 ± 2°C, 7 ± 2°C and -11 ± 3°C, the curves obtained for experimental germination and theoretical germination estimation were decreasing. They also have, in a very particular way, a sinusoidal shape; the amplitude of the signal also seems to widen with decrease in temperature. It should be remembered that the protocol adopted considers batches of seeds stored at different temperatures with samples of germinated grains regularly every week for 3 to 4 months. Comparing the three curves in Figure 3, we can see a decrease in the germination capacity of seeds stored at different temperatures as the storage time increases. However, this decrease is drastically reduced when the storage temperature is lowered.
Experimental results show that when G. simplicifolia seeds are stored at 24 ± 2°C, 3.71% or almost 4% of the seeds lose their germinative power each month. When stored at 7 ± 2°C, the germinative capacity of the seeds decreases with time. However, during the first month, this percentage remains close to 100%. However, the decreasing slope indicates that at this temperature, only 1.43% of the seeds lose their germinative capacity each month. For seeds stored at -11 ± 3°C, during the first month, the germination percentage remains close to 100%, except in the third and fourth weeks when it decreases to 95%. A decay coefficient of 1.41% is found for seeds stored at -11 ± 3°C.
The conservation of the germinative capacity of G. simplicifolia seeds depends indeed on storage temperature, the lower it is, the more it conserves. After 4 months of storage, germination rates are estimated at 30% for storage at 24 ± 2°C, 75 and 73%, respectively for storage at 7 ± 2°C and -11 ± 3°C (Figure 3A, B, and C). However, viability will also depend on shelf life, the sinusoidal appearance of the curves obtained, clearly shows that physiological processes are attenuated but not stopped and the extrapolations calculated using linear approximations make it possible to quantify the conservation of germinative power. At over 30°C, they will not survive for more than 8 weeks. Extrapolating to 12 months of conservation at 24 ± 2°C, nearly 45% of the seeds will have lost their germinative power. In the refrigerator, after 12 months of storage, 82.84% of the seeds would remain viable, whereas in the freezer part of the refrigerator, only 16.92% of the seeds would be lost in 12 months.
Analysis of the ability to predict the germination of G. simplicifolia as a function of temperature based on the coloring of shelled seeds
For storage at 24 ± 2°C, the study carried out over 4 months, shows that both curves decrease over time. However, there is good agreement between the theoretical estimate and the experimental results during the first month. Between the first and the fourth months, a discrepancy between the measured quantities can be observed. During the first month, the germination percentage remains practically at 100%, except in the last week when it is 95%. From the first to the fourth month, it can be seen that the estimated germinative power of the seeds is higher than the experimentally determined percentages. The germination percentage varies from 100 to 63% during this period and the estimated germination percentage varies from 95 to 30%. The linear approximation of the curves obtained by straight lines allows comparison of their slopes (steering coefficients) in order to better appreciate differences. In Figure 1, comparing the theoretical estimate with the experimental results, the steering coefficient goes from -2.00 to -3.71, with the R² coefficients in both cases higher than 0.8. The Welch mean comparison test allows us to conclude that the experimental germination percentages are higher than the estimated percentages, confirming the observed graphical results.
At 7 ± 2°C, the germination percentage of the seeds is decreasing with time. However, during the first month, this percentage remains close to 100%. The slopes of the two representations are almost the same (germination as well as its estimation according to storage temperature). The linear approximation of the two curves shows very similar slopes (steering coefficients). Differences are noted especially in the first month and towards the end of the fourth month. However, they remain below 3%. The slope of the linear approximation of the germination percentage is -1.29 with an R² coefficient of 0.56 and that of the estimated germination percentage is -1.43 with a correlation coefficient of 0.74. Experimental results show that when Griffonia seeds are stored at -7 ± 2°C, each month 1.29% of the seeds lose their germinative power. The theoretical estimate is close to the experimental values. The results of the Welch test confirm that the experimental germination results are equivalent to those resulting from germination estimates based on seed color.
For seeds stored at -11±2°C, both curves are decreasing over time. During the first month, the germination percentage remains practically at 100%, except in the third and fourth weeks when it decreases to 95%. The estimate of the germination percentage is also very good during the first month, compared to the experimental values.
From the first to the fourth month, it can be seen that the estimated germinative power of the seeds is in this case lower than the experimental percentages determined. The correlation coefficients are -2.95 for the estimate and -1.41 for the experimental germination percentage. However, there is no significant difference between the experimental results and the germination estimate.
The steering coefficients of the approximation lines for the curves are -3.71 at 24 ± 2°C, -1.2 at and -1.4 at 11 ± 3°C. Decay has been greatly reduced between 7 and -11°C. If at 24°C, a significant difference between the experimental (stronger) and estimated results was observed, when the conservation temperatures were lower, respectively 7 and -11°C, thus more efficient to conserve; the results are equivalent.
DISCUSSION
The control of the germination process of a species depends on a set of factors and environmental conditions that interact with the genetic characteristics of the plant material. Germination is a physiological process that allows the embryo contained in the seed to develop into a young seedling, thus ensuring the survival and sustainability of the species. It is well established that the process of germination begins with the imbibition of the seed and finishes with the piercing of the integument by the radicle that emerges first. The results obtained in this study with G. simplicifolia, highlight the crucial role of seed coat and light on this process, as well as the impact of the conditions in which the seeds are stored on their viability.
The preliminary decortication and anti-fungal treatment allow the improvement of the germination rate and can help maximize the germination of G. simplicifolia seeds. Unlike most legumes, which are subject to strong inhibition (Danthu et al., 2003), the seed coat of G. simplicifolia seeds is relatively permeable to water and therefore is not a barrier to water absorption by the embryo. However, the presence of a seed coat influences the germination kinetics of G. simplicifolia, their presence increases the average germination time and reduces germinative capacity. They are mechanical inhibitors of germination but they are also a source of fungal contamination which causes seed necrosis despite prior disinfection. Fungal contamination is relatively important since it reduces the germination capacity of G. simplicifolia seeds to 70% for whole seeds, whereas it is maximum for seeds germinated in the presence or absence of seed coat. Pre-soaking or soaking in water for 30 min before germination gives a germinative capacity of 90%. These results show that seed coats do not exert chemical inhibition on germination; however, they contain external microorganisms, which, even with prior disinfection, cause 30 and 10%, respectively of whole and peeled seeds contamination and germination in the presence of the seed coats to be necrotic. Contamination breaks the cotyledons, destroys the apical meristem and degrades the reserves of the cotyledons through necrosis, depriving the embryo of the nutrients essential for its growth.
Decortication and prior disinfection are essential as they reduce and even completely eliminate micro-organisms and achieve a maximum germination rate (Quashie and Tchezoum, 2009; Quashie and Ahama, 2018). Decortication is the most effective treatment method to achieve both a high germination rate and growth of young seedlings in the absence of bio-aggressors.
The germination of G. simplicifolia seeds is hypogeous, unlike that of many other legumes such as groundnut (Arachis hypogea) (Shorter and Patanothai, 1989), soybean (Glycine max) (Schiffers et al., 1982). Considering the peculiarity of the hypogeous germination of G. simplicifolia and the observed greening of the cotyledons, it is important that germination of this species takes place without burying the seeds. Indeed, in the absence of the observed greening of cotyledons when germinating in the dark, the seeds are vulnerable to external fungal contamination, which is the cause of the observed seed necrosis during germination; this explains the poor germination results of G. simplicifolia obtained so far (Bosch, 2008; Kumar et al., 2010).
After germination, the true cotyledons turn green before the first leaves appear. The cotyledons play a nutritive role in the seedling of G. simplicifolia, and later a partial rotting of the cotyledons results in a reduction of the reserves available to the seedling and less vigorous growth. In the case of G. simplicifolia a relatively long persistence of the green cotyledons after germination of the seeds of G. simplicifolia has been noted, these cotyledons could therefore be considered as the true first leaves, which remain even after the development of the seedling.
Light, by stimulating the greening of the cotyledons during germination, helps to improve germination speed. However, light has no influence on the germination of G. simplicifolia, which belongs to the restricted group of seeds with indifferent photosensitivity (Heller et al., 2000; Danthu et al., 2003). However, germination in darkness has a negative effect on the subsequent growth of the seedling. It does not favor the greening of the cotyledons. The greening of the cotyledons can be considered as the heterotrophic growth of the seedling that initiates the colonization of its environment. When chlorophyll appears, often very early, in the cotyledons, new carbonaceous materials are introduced to serve as carbonaceous nutrition; a new function develops; the phenomena of vegetative life are added to those of germinative life. The cotyledons are thus transformed into cotyledon leaves. According to the work of Tchang (1984), greening would be linked to the formation of glyoxysomes closely related to the metabolism of lipids and the regulation of peroxysomal activities by light. The seeds of G. simplicifolia contain lipids, the main one of which is linoleic acid which represents 53 to 60% of total lipids (Petkov and Ramazanov, 2003). The delayed greening of the cotyledons is a risk factor for the seedling, because the cotyledons become sensitive to the microorganisms that cause their necrosis. Thus, the longer the cotyledons remain in the dark, the greater the risk of necrosis. The increase in germination time and the increase in heterotrophic growth when conditions are unfavorable increase the risk of attack by different microorganisms or pests (Brunel, 2008). In G. implicifolia, the frequency of necrosis of the cotyledons and the seedlings from germination in the dark and the persistence of the cotyledons, which remain green for a very long time in seedlings from germination in light, show that the rapid greening of the cotyledons plays an important role during the early stages of seedling development.
The seed of G. simplicifolia is mainly oleo-proteaginous. Seeds with a high total lipid content have a slower germination than seeds with starchy reserves in the face of a low O2 content in the medium (Raymond et al., 1985). However, G. simplicifolia, which is an oil-protein seed, has an average germination time of 2 days under ideal conditions.
The reserves of G. simplicifolia seeds are easily altered and induce a short life span, thus confirming that this seed is indeed an oleo-proteinaceous one. The climatic conditions (temperature, light, humidity) can influence the viability of these seeds; under ambient conditions of humidity and temperature, seeds of G.simplicifolia are very hygrophilic, do not store well and rot quickly in situ. This study shows that cold storage (7 ± 2°C or -11 ± 3°C) improves the germination capacity of G. simplicifolia. These temperatures allow the preservation of their viability (75% of the seeds germinate after 4 months of storage). Similar results were obtained by Danthu et al. (2003) with Acacia raddiana seeds (96% at -18°C and 80% at room temperature). These low temperatures slow down the browning of the cotyledons, a sign of lengthening of the viability of the seeds. The low temperatures slow down the oxidation and peroxidation of the seed reserves; this explains the lengthening of the viability of G. simplicifolia seeds stored in the refrigerator (freezer section) and in the freezer, therefore at temperatures below 7°C. The peroxidation and oxidation of the unsaturated fatty acids in the lipids produce highly reactive free radicals, hydro peroxides, and secondary products that accelerate the ageing of the seeds through damage to the membranes, enzymes and chromatin of the cells (Le Buanec and Lechappe, 2006). As G. simplicifolia seeds are rich in lipids, particularly unsaturated fatty acids (linoleic acid and oleic acid), followed by saturated fatty acids (stearic acid and palmitic acid), it is therefore the progressive oxidation of the reserves of G. simplicifolia seeds that leads to the loss of viability (Vigliante et al., 2019). A correlation between the change in color of the cotyledons (from yellowish to brownish during storage) and the preservation of viability of G. simplicifolia seeds makes it possible to sort the seeds. Seeds that lose their yellowish color lose their germination capacity. This result is in agreement with that obtained on soybeans; a relationship between seed color and viability maintenance has been established (Nieuwenhuis and Nieuwelink, 2005). Biological rhythms, which are generated endogenously, are sensitive to environmental information which allows a stable phase relationship to be maintained with the cycles of the environment, thus making it possible to adjust the physiological cycle of the variables necessary for survival. When this relationship is established, the organism is considered to be synchronized with the environment (Pontes et al., 2010). Factors detrimental to good seed conservation are high temperature and humidity, as well as a seed moisture content of more than 30%. Fluctuations in storage conditions are also detrimental to adequate seed preservation (Gaudreault, 2005).
The biotechnological improvement of any plant involves controlling its culture in vitro, in particular its germination in vitro (Bosch, 2008; Kumar et al., 2010). In vitro introduction requires disinfection. Disinfection is not a hindrance to germination, but rather a gain that prevents the seedling from necrosis caused by contaminating agents. The disinfection protocol does not affect the germination capacity of the seeds in vitro. However, when the disinfection solutions are too concentrated, they cause a burning of the cotyledons, which start to release compounds in the culture medium that cause browning. These compounds, probably polyphenols (Margara, 1984; Nyarko et al., 2019), delay seed germination when produced in large quantities. The presence of brown polyphenols that may, depending on the species, be released into the growing medium was reported by Yokotani-Tomita et al. (in 1998).; Kato and Arima (2006) showed that seeds release during germination a range of specific compounds with trophic or signaling roles, which are important in the early stages of plant life. As the seedlings grow on the growing medium, root exudation causes the same brownish coloration of the growing medium. The roots therefore release the same compounds as the cotyledons of the germinating seeds. However, in quantitative terms, exudation is dominated by low molecular weight soluble compounds such as sugars, organic acids and amino acids, which are present in high concentrations in the cytoplasm (Farrar et al., 2003). L-5-HTP would probably be among the compounds released into the culture media for two reasons, first it is an amino acid derivative and second the brownish color caused by exudation, refers to the natural color of the L-5-HTP extract.
CONCLUSION
This investigation has examined the necessary pre-treatments of the seed and the conditions of G. simplicifolia germination. It highlights the fact that prior decortication and disinfection are essential to improve germination. Whether it takes place in alternating day/night or in complete darkness, seeds with indifferent photosensitivity germinate with the same celerity. Many aspects concerning the ecology and physiology of the germination of this species are now well known and the control of germination in vitro opens the way for the introduction and in vitro multiplication of this species with a view to its domestication and conservation. The conservation of the germination capacity of G. simplicifolia seeds is influenced by the conditions of conservation. Temperature and humidity are therefore the two main factors that should be controlled to maintain seed viability. It is now clear from this investigation that cold storage can extend seed viability. However, the state of maturity of the harvested seeds and possibly an antifungal treatment would prolong their viability.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
The authors thank the Mathematics Department of the Faculty of Science, University of Lomé, particularly colleague Edarh-Bossou and the late colleague Boussari, for their involvement in the understanding of the mathematical models used in the manuscript.
REFERENCES
|
Addotey JNA, Adosraku RK (2016). Pilot production of 5-HTP from the seeds of Griffonia simplicifolia. World Journal of Pharmacy and Pharmaceutical Sciences 5(6):204-221. |
|
|
Asiamah I, Adosraku RK, Owusu-Ansah E, Abassah-Oppong S (2006). Post-Harvest Stability of 5-Hydroxytryptophan in Griffonia Simplicifolia Seeds. International Journal of Pure and Applied Chemistry 1(1):129-131. |
|
|
Bell EA, Fellows LE, Qureshi MY (1976). 5-Hydroxy-L-tryptophan; taxonomic character and chemical defense in Griffonia. Phytochemistry 15(5):823. |
|
|
Bhatla SC, Lal AM (2018). Plant Physiology, Development and Metabolism. Springer Singapore, Gateway East, Singapore 1251p. |
|
|
Birdsall TC (1998). 5-Hydroxytryptophan: a clinically-effective serotonin precursor. Alternative Medicine Review: A Journal of Clinical Therapeutic 3(4):271-280. |
|
|
Bosch CH (2008). Griffonia simplicifolia (Vahl ex DC.) Baill. In: Schmelzer GH & Gurib-Fakim A. PROTA, Wageningen, Netherlands (Editors). Prota 11(1). Medicinal plants/Plantes Médicinales 1 868p. |
|
|
Brunel S (2008). Caractérisation écophysiologique de différents génotypes de Medicago truncatula au cours des phases de germination et de croissance hétérotrophe. Thèse de doctorat, Université d'Angers, France. 279p. |
|
|
Czabator FJ (1962). Germination value: an index combining speed and completeness of pine seed germination. Forest Science 8(4):386-396. |
|
|
Danthu P, Roussel J, Neffati M (2003). La graine et la germination d'Acacia raddiana. In: Grouzis M. & Le Floc'h E., eds. Un arbre au désert Acacia raddiana. Paris: IRD 265-283. |
|
|
Djavanshir K, Pourbeik H (1976). Germination value-a new formula. Silvae Genetica 25(2):79-83. |
|
|
Elyasi S, Mollasadeghi V, Mirzamasoumzadeh B (2014). Study of Genetic Diversity of Rice of Some Germination Components and Growth Indices. Agriculture Science Developments 3(1):143-146. |
|
|
Esechie HA (1994). Interaction of salinity and temperature on the germination of sorghum. Journal of Agronomy and Crop Science 172(3):194-199. |
|
|
Esposito M, Precenzano F, Sorrentino M, Avolio D, Carotenuto M (2015). A medical food formulation of Griffonia simplicifolia/magnesium for childhood periodic syndrome therapy: An open-label study on motion sickness. Journal of Medicinal Food 18(8):916-920. |
|
|
Food and Agriculture Organisation (FAO) (1999). ''State of the world's forests.'' Italy, Rome: Food & Agriculture Organisation (Eds). |
|
|
Farrar J, Hawes M, Jones D, Lindow S (2003). How roots control the flux of carbon to the rhizosphere. Ecology 84(4):827-837. |
|
|
Gashi B, Abdullai K, Mata V, Kongjika E (2012). Effect of gibberellic acid and potassium nitrate on seed germination of the resurrection plants Ramonda serbica and Ramonda nathaliae. African Journal of Biotechnology 11(20):4537-4542. |
|
|
Gaudreault M (2005). Amorçage et séparation des graines d'épinette noire (Picea mariana (Mill.) BSP): amélioration de la germination des lots de semences forestières. Mémoire de maîtrise, Université du Québec à Chicoutimi 98 p. |
|
|
Heller R, Esnault R, Lance C (2000). Tome 2, Développement, 6ème édition de l'Abrégé. Dunod, Paris; New York; Barcelone; Mexico: Masson. 366p. |
|
|
Hutchinson J, Dalziel JM (1954). Flora of West Tropical Africa. Vol. I, Part 1, 2nd Edition. Crown Agents for Overseas Governments Administrations. London, United Kingdom 1954p. |
|
|
Joshi C, Kumar R, Nailwal TK (2016). Effect of gibberellic acid, potassium nitrate and chilling on seed germination response of apple (Pyrus malus L. Cv. Red delicious). International Journal of Advanced Research 4(6):1141-1155. |
|
|
Kadévi K (2001). Statistiques sur les produits forestiers non ligneux (PFNL) dans la république togolaise (Ministère de l'Environnement et des Ressources Forestières). EC/FAO ACP collecte et analyse de données-rapport technique: AFDCA/TN/20 40 p. |
|
|
Kafi FM, Goldani M (2001). Effects of Water Potential and Type of Gsmoticum on Seed Germination of Three Crop Species of Wheat, Sugarbeet, and Chikpea. Agricultural Sciences and Technology 15(1):121-133. |
|
|
Kakou BA, Kabran GRM, N'Guessan AH, Bohoussu VK, Benie A, Mamyrbékova-Békro JA, Békro YA (2018). Investigations phytochimique et antioxydante de cinq plantes médicinales ivoirienne utilisées contre la carie dentaire. International Journal of Innovation and Applied Studies 24(4):1794-1804. |
|
|
Kato K, Arima Y (2006). Potential of seed and root exudates of the common bean Phaseolus vulgaris L. for immediate induction of rhizobial chemotaxis and nod genes. Soil Science and Plant Nutrition 52(4):432-437. |
|
|
Kumar PS, Praveen T, Jain N, Jitendra B (2010). A review on Griffonia simplicifollia-an ideal herbal anti-depressant. Phytomedicine 18(10):848-851. |
|
|
Le Buanec B, Lechappe J (2006). Les semences évolution au cours des 50 dernières années. Comptes Rendus de l'Académie d'agriculture de France 92(3):21-41. |
|
|
Lemaire PA, Adosraku RK (2002). An HPLC method for the direct assay of the serotonin precursor, 5?hydroxytrophan, in seeds of Griffonia simplicifolia. Phytochemical Analysis: An International Journal of Plant Chemical and Biochemical Techniques 13(6):333-337. |
|
|
Maddox DE, Shibata S, Goldstein IJ (1982). Stimulated macrophages express a new glycoprotein receptor reactive with Griffonia simplicifolia I-B4 isolectin. Proceedings of the National Academy of Sciences 79(1):166-170. |
|
|
Margara J (1984). Bases de La Multiplication Végétative?: les mécanismes de l'organogenèse. Institut national de la Recherche agronomique, Paris 262 p. |
|
|
Mehta H, Mangrulkar S, Chourasia A (2015). A review on Griffonia simplicifollia-A natural anti-depressant. International Journal of Phytopharmacology 6(2):76-79. |
|
|
Mora-Villalobos JA, Zeng AP (2018). Synthetic pathways and processes for effective production of 5-hydroxytryptophan and serotonin from glucose in Escherichia coli. Journal of Biological Engineering 12(1):1-12. |
|
|
Murashige T, Skoog F (1962). A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiologia Plantarum 15(3):473-497. |
|
|
Nieuwenhuis R, Nieuwelink J (2005). La culture du soja et d'autres legumineuses. Agromisa, CTA. Wageningen; Pays-Bas 76 p. |
|
|
Nyarko RA, Larbie C, Anning AK, Baidoo PK (2019). Phytochemical constituents, antioxidant activity and toxicity assessment of hydroethanolic leaf extract of Griffonia simplicifolia. International Journal of Phytopharmacology 10(1):6-18. |
|
|
Petkov G, Ramazanov Z (2003). Fatty acids and sterols of Griffonia seeds oil. Grasas y Aceites 54(1):30-31. |
|
|
Pontes ALBD, Engelberth RCGJ, Nascimento Jr EDS, Cavalcante JC, Costa MSMDO, Pinato L, de Toledo CAB, Cavalcante JDS (2010). Serotonin and circadian rhythms. Psychology and Neuroscience 3(2):217-228. |
|
|
Quashie MLA, Tchezoum YA (2016). Étude de la germination de Moringa oleifera LAM. Afrique Science 5(3):169-180. |
|
|
Quashie-Mensah Attoh A, Ahama KYS (2018). Multiplication générative de Crataeva adansonii DC. Journal de La Recherche Scientifique de l'Université de Lomé 20(3):103-116. |
|
|
Rajabi R, Poustini K (2005). Effects of NaCl salinity on seed germination of 30 wheat (Triticam aestivum L.) cultivars. Scientific Journal of Agriculture 28(1):29-44. |
|
|
Schiffers B, Cornet D, Fraselle J, Mboka-Unda B (1982). Etude de l'association du Rhizobium et de l'insecticide carbofuran dans le pralinage des semences de soja (Glycine max (L.) MERRIL). Parasitica, 38(2):55-63. |
|
|
Shah S, Gong ZH, Arisha MH, Khan A, Tian SL (2015). Effect of ethyl methyl sulfonate concentration and different treatment conditions on germination and seedling growth of the cucumber cultivar Chinese long (9930). Genetics and Molecular Research 14(1):2440-2449. |
|
|
Shorter R, Patanothai A (1989). Arachis hypogaea L. In: van der Maesen, L.J.G. & Somaatmadja, S. (Editors). Plant Resources of South-East Asia No 1. Pulses. Pudoc, Wageningen, Netherlands, pp 35-39. |
|
|
Tchang F (1984). Biogenèse des glyoxysomes et des peroxysomes au cours du développement des plantules de tournesol (Helianthus annuus L.) ou de Pharbitis nil (Chois.). Thèse d'Etat. |
|
|
Vigliante I, Mannino G, Maffei ME (2019). Chemical Characterization and DNA Fingerprinting of Griffonia simplicifolia Baill. Molecules 24(6):1032. |
|
|
Welch BL (1947). The generalization of student's' problem when several different population variances are involved. Biometrika 34(1-2):28-35. |
|
|
Yokotani-Tomita K, Goto N, Kosemura S, Yamamura S, Hasegawa K (1998). Growth-promoting allelopathic substance exuded from germinating Arabidopsis thaliana seeds. Phytochemistry 47(1):1-2. |
|
|
Zavarise P, Dalla VG (2017). A Combination of Tanacetum parthenium, Griffonia simplicifolia and Magnesium (Aurastop) as Symptomatic Acute Treatment for Migraine Aura: A Retrospective Cohort Study. Open Access Library Journal 4(6):1-9. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


