Full Length Research Paper

ABSTRACT
Axenic cultures of Klebsiella pneumoniae, Escherichia coli and Staphylococcus aureus obtained from the Medical Microbiology Laboratory of the Delta State University Teaching Hospital, Oghara, were inoculated into ten-fold serial dilutions of 10-1 to 10-7 in sterile Nutrient broth and incubated in water bath at 37°C for 18 h initially. The turbid broth cultures of 10-5, 10-6 and 10-7 dilutions were subcultured aseptically on sterile MacConkey agar, Nutrient agar and Blood agar plates and incubated at 37°C for 18 h. Antibiotic sensitivity testing was done using gentamycin, nalidixic acid, nitrofurantoin, ofloxacin, cotrimoxazole, tetracycline, amoxicillin and amoxicillin/clavulanic acid on isolates from the last three dilutions selected and results were recorded as before temperature treatment. Serial dilutions were repeated and the last three dilutions were subcultured on same media as before treatment and all inoculated plates were incubated at 30, 35, 40 and 45°C in the water bath for 18 h. Antibiotic sensitivity testing was then carried out using the same drugs and results were recorded as after treatment. Mean±standard error zones of inhibition (ZI) recorded for 10-5, 10-6 and 10-7 E. coli broth dilutions at 37°C by gentamycin, nalidixic acid, nitrofurantoin and ofloxacin were 13.8±10.0, 16.8±11.0, and 14.0±8.1 mm, respectively. Same broth dilutions of K. pneumoniae recorded 17.6±9.5, 14.4±6.0 and 13.8±9.3 mm ZI, respectively to same drugs. Similarly, same broth culture dilutions of S. aureus recorded 3.4±7.1, 2.8±6.9 and 2.8±5.7 mm ZI, respectively to same antibiotics. There was resistance reduction (RR) of 76.2±11.6 and 57.0±14.3% by 10-6 and 10-5 broth dilutions of E. coli at 30 and 40°C, respectively. In all three broth culture dilutions of K. pneumoniae, there was a less than 30% RR at all tested temperatures. S. aureus broth dilutions of 10-5, 10-6 and 10-7 recorded 173.9±6.4, 150.0±7.3 and 87.0±6.2% RR, respectively to ofloxacin at 30°C. At 35°C, 10-7 S. aureus broth dilution recorded 87.0±4.3% RR. There was more than 50% RR in 10-5 and 10-6 S. aureus broth dilutions at 40°C and more than 50% RR in all three dilutions at 45°C. Vertically, the three pathogens in 10-7 broth dilution and at 40°C recorded a common RR of 58.1±11.2% against gentamicin. Broth culture dilutions of 10-5 and 10-6 of all pathogens produced RR of 74.6±22.2 and 74.6±21.0%, respectively against ofloxacin with no significant RR against the other drugs at the temperatures tested. The implications of these findings are discussed.
Key words: Elevated, temperature, resistance reduction, clinical pathogens.
INTRODUCTION
Numerous classes of antimicrobial agents have become less effective as a result of the emergence of antimicrobial resistance often as a result of the selective pressure of their usage (Oskay et al., 2009). This selective pressure can be attributed to indiscriminate use of antibiotics, complex socio-economic behavioural antecedents and dissemination of drug resistant pathogens in human medicine (Okeke et al., 1999). Moreover, the dis-appointing lack of new antimicrobial agents has led to overuse of existing ones thus leading to the emergence of multi-resistant pathogens (McGowan and Wise, 2001).
Therefore, as the proliferation of multidrug resistant pathogens continues unavoidably within and around us, it is important that their resistance trend be put under check through intensive research and antibiotics surveillance (Akortha and Filgona, 2009). The primary causes of antibiotic resistance in bacteria are mobile elements called plasmids and conjugative transposons. Plasmids are extra chromosomal DNA elements that have the capacity to replicate independently of the chromosome of the bacterial cell (Madigan et al., 2003). Resistance plasmids or R plasmids code for enzymes that can inactivate antibiotics, prevent the uptake of an antibiotic or pump out a particular antibiotic (Neu, 1989).
Curing is the process of removing plasmids from a bacterial cell (Trevors, 1986). The resulting bacteria then become sensitive to the selective agent. It was initially thought that the phenomenon of curing would proffer solution in controlling the development of antibiotic resistance in formerly antibiotic susceptible bacteria. Sadly, many plasmids are retained in the absence of the selective agent and this could be due to the presence of DNA cassettes that encode for stability (Zielenkiewicz and Ceglowski, 2001).
Novobiocin, ethidium bromide, acriflavine, acridine orange, ascorbic acid and elevated temperatures have been used as curing agents (Ramesh et al., 2000). Physical treatments, chemical compounds and growth conditions may increase the frequency of elimination of drug resistant R-plasmids resulting in sensitive cells that were previously resistant to antibiotics (Lakshmi et al., 1989). DNA intercalating dyes (ethidium bromide), sodium dodecyl sulphate (SDS), antibiotics, thymine starvation and elevated temperatures have been used as curing agents (Chakrabarrty et al., 1984; Gupta et al., 1980; Obaseki-Ebor, 1984; Reddy et al., 1986). The various researched curing agents may have several different modes of action. They may affect membrane potential, membrane permeability, protein synthesis and the processing of DNA (Viljanen and Boratynski, 1991). Some curing compounds block plasmid transfer (Zhao et al., 2001).
Temperature is a basic requirement for bacterial growth and most clinically significant microorganisms are mesophiles that grow optimally at temperatures between 43 and 44°C .Temperature is among the environmental factors microorganisms require for their survival (Goller and Romeo, 2008; Rachid et al., 2000). High temperature will often bring about increased growth rate due to catabolic processes (such as respiration) being surpassed by anabolic processes such as protein synthesis (Degerman et al., 2013).
Growth of Staphylococcus aureus at 45°C increased the rate of loss of penicillinase production (Asheshov, 1966a or b). Fairbrother et al. (1954) reported that growth of some strains of Staphylococci at elevated temperature (44°C) increased the rate of loss of penicillin resistance. May et al. (1964) reported loss of both penicillin and tetracycline resistance by S. aureus when grown at elevated temperature of 43 to 44°C. Bondi et al. (1958) showed that growth of S. aureus at 45°C increased the rate of loss of penicillinase production.
Stanier et al. (1984) reported that whereas elevated temperature of 40°C alone was ineffective in curing Pseudomonas putida, the plasmid or resistance marker of this organism was cured up to 2.63% when elevated temperature (40°C) and acridine orange combined were used together. Curing by elevated temperature is the most efficient method among others and this may be due to the fact that the enzymes which mediate DNA replication processes are more affected by high temperature (Kheder, 2002). Enzyme inactivation may be due to a change in the folding of polypeptide at high temperature (40 to 46°C), that is, the enzymes are sensitive to elevated temperature (Kheder, 2002).
The antibacterial activity of antimicrobial agents against some strains of S. aureus was significantly enhanced by increasing the incubation temperature (Hajdu et al., 2009). Incubation of the strains with antibiotics at 40 or 45°C resulted in significant reductions in bacterial growth (Hajdu et al., 2009). Elevated temperature (46°C) treatment of three strains of P. aeruginosa efficiently cured all the plasmids being harboured within the genome of these organisms (Radi and Rahman, 2010). The genes located on the chromosomal DNA were however not affected by high temperature treatment compared with that encoded by plasmid DNA (Radi and Rahman, 2010).
Treatments that increase the frequency of elimination of plasmids will enhance microbial susceptibility to antibiotics. To our knowledge, there was no published current work on elevated temperature treatment of multidrug resistant bacterial pathogens, such as Escherichia coli, Klebsiella pneumonia, and Staphylococcus aureus which are pathogens implicated in a great number of human diseases. The focus of this work, therefore, is antibiotic resistance marker reduction of pure broth cultures of selected clinical pathogens after elevated temperature treatments with the following objectives:
1. Determine the antibiotic susceptibility profile of selected clinical isolates (E. coli, K. pneumoniae and S. aureus) after incubation at 37°C for 24 h before elevated temperature treatment.
2. Show or evaluate ≤50.0% resistance marker (RM) reduction of clinical pathogens at different temperatures (30, 35, 40 and 45°C) of incubation for 18h.
3. Show a summary of ≤50.0% RM reduction of clinical pathogens at all tested elevated temperatures.
MATERIALS AND METHODS
Sampling
Axenic (pure) isolates of K. pneumoniae, E. coli and S. aureus isolated from midstream urine and infected wound samples were obtained from the Medical Microbiology Department of Delta State University Teaching Hospital, Oghara, Nigeria. Pathogens which were appropriately labelled and stocked on sterile Nutrient agar slants were then transported to the Microbiology and Biotechnology Laboratory of the Western Delta University, Oghara for immediate processing.
Processing
Stocked cultures of named pathogens were re-confirmed by subculturing them on selective and sterile agar media and incubated appropriately at 37°C (Cowan and Steel, 1996). Stocked cultures of K. pneumoniae and E. coli were subcultured on sterile Nutrient agar, MacConkey agar and Cystine Lactose Electrolyte Deficient agar plates. S. aureus stock culture was subcultured on sterile MacConkey agar and Mannitol Salt agar plates. All inoculated plates were incubated at 37°C for 24 h.
The resulting colonies were identified by standard methods of Cowan and Steel (1996) and Cullimoore (2000). Isolates were identified culturally, morphologically, biochemically and by sugar fermentation. Gram negative, raised, entire, circular, mucoid, motile, lactose, glucose fermenting, indole positive, methyl red positive, voges praskauer negative, citrate negative and urease negative bacilli in singles were identified as K. pneumoniae. Red to pink smooth and convex colonies (on MacConkey agar), mucoid or non-mucoid yellowish smooth and convex colonies (on CLED agar), gram negative bacilli in singles, indole positive, methyl red positive, voges proskauer negative, citrate negative, urease negative, gelatine liquefaction negative colonies with gas from glucose and lactose fermentation were confirmatory of E. coli. Bacterial organism that showed gram positive cocci in clusters, catalase positive, DNAase negative, mannitol fermenting, raised, round and smooth colonies were identified as S. aureus. All three re-confirmed isolates were subjected to serial dilutions in sterile Nutrient broth.
Serial dilution (ten-fold dilution) of isolates
Each isolate was first inoculated into 10 ml sterile Nutrient broth and incubated at 37°C for 18 h. The resulting turbid (cloudy) broth cultures (well labelled) were used for the serial dilutions. For each isolate, seven sterile test tubes each containing 9 ml of sterile Nutrient broth were set on a test tube rack and labelled 10-1, 10-2 to 10-7. Into the first tube, 1.0 ml of the nutrient broth culture was pipetted and mixed. From this tube, 1 ml was pipetted (using another sterile pipette) into tube 2, mixed and from this, 1 ml was pipetted into the next till the 7th tube. From the 7th tube, 1 ml was pipetted and discarded. All inoculated tubes with their dilution concentrations were incubated in a water bath at 37°C for 18 h. At the end of incubation, the last three turbid tubes (having dilutions 10-5, 10-6 and 10-7) were subcultured on sterile MacConkey and Blood agar plates and incubated at 37°C for 24 h. The resulting colonies were then used for antibiotic sensitivity testing.
Antibiotic sensitivity testing
Antibiotic sensitivity testing was done on all three bacterial isolates and done according to the disc diffusion method by Bauer et al. (1996). A loopful of a colony was picked aseptically using a flamed and cooled wire loop and placed in the centre of three sterile Mueller Hinton agar plates (each of the three plates was used for each dilution earlier subcultured). That is, three sterile Mueller Hinton agar plates were used for each isolate. The loopful of a colony in the centre of each plate was then spread all over the plate applying the caution of not touching the edges of the plate. The seeded plates were allowed to stand for about 2 min to allow the agar surface to dry. The appropriate multi- discs containing minimum inhibitory concentrations (MIC) of gentamicin (10 µg), nalidixic acid (30 µg), nitrofurantoin (300 µg), ofloxacin (10 µg), cotrimoxazole (25 µg), amoxicillin (25 µg), tetracycline (25 µg) and augmentin (30 µg), chloramphenicol (30 µg), erythromycin (5 µg) and cefuroxime (30 µg) were then aseptically placed (impregnated) firmly onto the surface of the dried plates using a pair of sterile forceps. Discs were placed at least 22 mm from each other and 14 mm from the edge of the plates (Ochei and Kolhatkar, 2008). All plates were allowed to stand on laboratory bench for 10 min before incubation (Mbata, 2007).
Antibiotic discs used were selected on the basis of their clinical importance and efficacies on E. coli, K. pneumoniae and S. aureus. The plates were left at room temperature for about 30 min to allow for diffusion of the different antibiotics from the discs into the medium. The plates were then incubated at 37°C for 18 to 24 h. At the end of incubation, the diameters of the zones of inhibition from one edge to the opposite edge were measured to the nearest millimetre using a transparent ruler. Zones of inhibition greater than 10 mm were considered sensitive, 5-10 mm as moderate sensitive and no zone of inhibition as resistant (NCCLS, 2000).
Temperature treatments of clinical pathogens
Serial dilutions (ten-fold dilution) were again carried out using the original overnight Nutrient broth culture of each isolate into a set of seven sterile test tubes each containing 9 ml of sterile nutrient broth. One millilitre (1.0 ml) of broth culture was pipetted into the first tube and mixed and then 1.0 ml was pipetted from first tube into the second and so on till the seventh tube to give 10-1, 10-2 to 10-7 as earlier described. All tubes were then incubated at 30°C in a water bath. The dilutions were repeated using new sets of sterile tubes and incubated in water bath at 35, 40 and 45°C. This was separately done for each isolate. At the end of incubation, all turbid tubes were recovered and only 10-5, 10-6 and 10-7 broth cultures were subcultured on MacConkey agar and Cystine Lactose Electrolyte Deficient agar plates and incubated at 30, 35, 40 and 45°C for 18 to 24 h. Antibiotic sensitivity testing was then carried out on the resulting colonies of each isolate after the various temperature treatments as previously described.
RESULTS
The values of zones of inhibition (in millimetres) by gentamicin, nalidixic cid, nitrofurantoin, ofloxacin,cotrimoxazole, amoxicillin, tetracycline, and amoxicillin/clavulanic acid against 105 to 107 dilutions of overnight broth pure cultures of E. coli (isolated from urinary system), K. pneumoniae (isolated from urinary system) and S. aureus (isolated from an infected wound) after incubation at 37°C for 24 h are shown in Table 1.
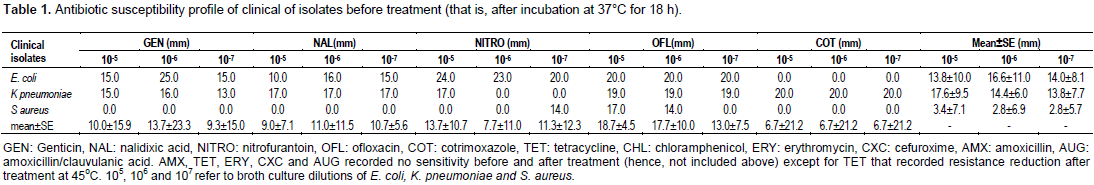
The zones of inhibition recorded for 105 E. coli broth dilution by gentamicin, nalidixic acid, nitrofurantoin and ofloxacin ranged from 10.0 to 25.0 mm with a mean±standard error (SE) of 13.8±10.0 mm zone of inhibition. The same antibiotics produced 16.0 to 25.0 mm range of inhibition zone for 106 E. coli dilution with a mean of 16.8±11.0 mm. The dilution 107 of E. coli broth recorded a mean of 14.0±8.1 mm zone of inhibition and a range of 15.0 to 20.0 mm to the same antibiotics.
K. pneumoniae 105, 106 and 107 broth culture dilutions recorded inhibition zone ranges of 15.0 to 20, 16.0 to 20.0 and 13.0 to 20.0 mm, respectively with mean±SE of 17.6±9.5, 14.4±6.0 and 13.8±9.3 mm, respectively against the same drugs.
Broth culture dilutions (105, 106, 107) of S. aureus produced zero zones of inhibition to the tested drugs apart from ofloxacin which recorded 14.0, 17.0 and 14.0 zones of inhibition, respectively. Mean±SE of 3.4±7.1, 2.8±6.9 and 2.8±5.7 mm were however recorded for the broth dilutions, respectively.
Vertically, 105, 106 and 107 dilutions of all pure bacterial pathogens recorded 10.0±15.9, 13.7±23.3 and 9.3±15.0 mm zones of inhibition, respectively to gentamicin. To the same dilutions of all three pathogens, zones of inhibition of 9.0±7.1, 11.0±11.5 and 10.7±5.6 mm were recorded by nalidixic acid in that same order. In that same order also, nitrofurantoin recorded 13.7±10.7, 7.7±11.0 and 11.3±12.3mm zones of inhibition. Whereas, 105, 106 and 107 broth culture dilutions of all pathogens recorded 18.7±4.5, 17.7±10.0 and13.0±7.5 mm zones of inhibition, respectively to ofloxacin, the mean ± SE zones of inhibition recorded by cotrimoxazole for the same broth dilutions were 6.7±21.2, 6.7±21.2 and 6.7±21.2 mm, respectively.
In Table 2, the indications of 50% (or more) resistance marker reduction of clinical bacterial pathogens after temperature treatment of 30°C are shown. Although, there was 0.0% resistance marker (RM) reduction for 105, 106 and 107 E. coli broth culture dilutions, the mean ± SE zones of inhibition indicated 0.0, 76.2 and 0.0% for the dilutions, respectively suggesting that 106 E. coli broth culture dilution produced a RM reduction of 76.2% which was much higher than the 50.0% bench mark used in this study.
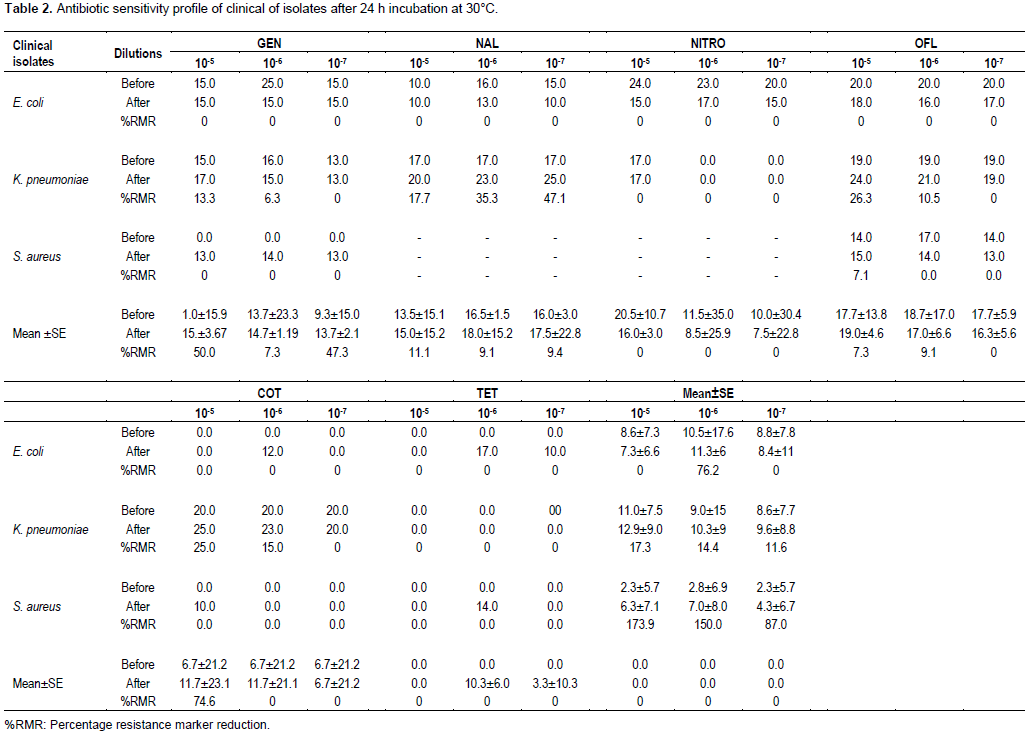
K. pneumoniae 105 broth culture dilution at 30°C temperature treatment produced RM reductions of 13.3 to 26.3% with a mean ± SE zone of inhibition of 17.3±8.3% by all tested drugs. Broth culture dilutions of 106 and 107 of the same organism recorded RM reductions of 6.3 to 35.3% and 0.0 to 47.1% respectively with means±SE zones of inhibition of 14.4±12.0 and 11.6±8.3%, respectively.
Dilutions of S. aureus (105, 106 and 107) recorded RM reductions of 0.0 to 7.1, 0.0 and 0.0%, respectively and means±SE zones of inhibition of 173.9±6.4, 150±7.3, and 87.0±6.2%, respectively for all eight antibiotics tested. Of note, antibiotic resistance reversal was recorded for gentamicin after 30°C temperature treatment for all dilutions. A similar situation occurred for 105 dilution with cotrimoxazole.
Vertically, 105, 106 and 107 dilutions of all three pure broth bacterial cultures recorded 50.0±9.8, 7.3±12.2 and 47.3±8.5% RM reductions, respectively to gentamicin. The same broth culture dilutions in that order produced resistance marker (RM) reductions of 11.1±15.0, 9.1±8.4 and 9.4±12.9%, respectively to nalidixic acid. Mean ± SE resistance marker reduction of 0.0, 0.0 and 0.0% were recorded for the same dilutions with nitrofurantoin.
Resistance marker reductions of 7.3±9.2, 9.1±11.8 and 0.0% were recorded for 105, 106 and 107 broth culture dilutions, respectively of all pathogens with ofloxacin and 74.6±22.2, 74.6±21.0 and 0.0% RM reductions for all three dilutions, respectively were recorded for cotrimoxazole.
Table 3 displays the data recorded for all three pathogens after 35°C temperature treatment. There was 0.0% RM reduction in E. coli broth culture dilution of 105 for all the tested antimicrobial agents with equally 0.0% Mean ± SE RM reduction. Apart from 6.7% RM reduction recorded for 10ˉ7 broth culture dilution, there was also 0.0 and 0.0% RM reduction and 0.0 and 0.0 mean± E RM reduction for 10ˉ6 and 10ˉ7 E. coli broth culture dilutions, respectively.
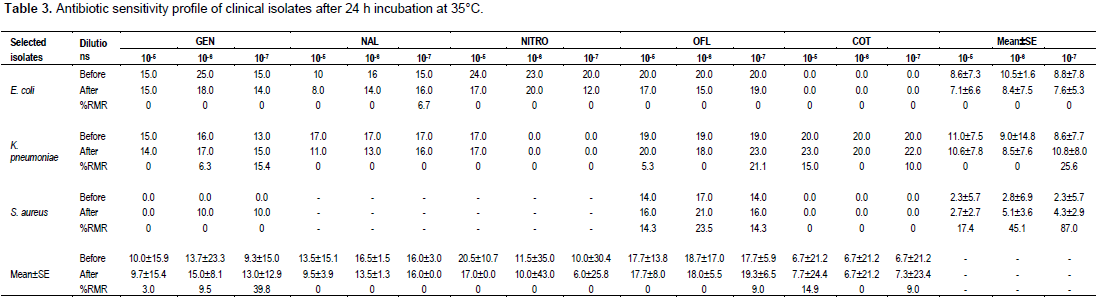
For all the eight antibiotics tested, broth culture dilutions 10ˉ5 and 10ˉ6 of K. pneumoniae recorded RM reductions of 5.3 to 15.0% and 0.0 to 6.3%, respectively with mean±SE RM reductions of 0.0 and 0.0%, respectively, while 10ˉ7 dilution showed a RM reduction of 10.0 to 21.1% with a mean ± SE of 25.6%. Broth culture dilutions of S. aureus produced 0.0% RM reduction for all antibiotics tested apart from ofloxacin which recorded RM reductions of 14.3, 23.5 and 14.3% for 105, 106 and 107, respectively with mean±SE RM reductions of 17.4, 45.1 and 87.0%, respectively. A significant RM reduction of 87.0% was therefore recorded for 107 dilutions.
The three broth culture dilutions (105,106 and 107) of all three pathogens together (E. coli, S. aureus and K. pneumoniae) recorded mean±SE RM reductions of 3.0±15.7, 9.5±15.7 and 39.8±14.0%, respectively when gentamicin was applied. The application of nalidixic acid for the same dilutions and for all three pathogens recorded mean±SE RM reductions of 0.0, 0.0 and 0.0%, respectively. Similar RM reductions were recorded with nitrofurantoin. Ofloxacin recorded 9.0±22.3% RM reductions for 105, 106 and 107 broth cultures dilutions, respectively for all three pathogens together.
Data obtained at 40°C temperature exposure of E. coli broth culture dilutions are shown in Table 4. Broth culture dilutions of 105 for all eight antibiotics tested recorded 0.0 to 50.0% RM reduction while 106 and 107 broth dilutions recorded 0.0 to 18.8 and 13.3 to 33.3% RM reductions, respectively. The mean ± SE for 105, 106 and 107 E. coli broth culture dilutions after 40°C temperature treatment were 2.3±7.5, 0.0 and 2.3±7.9%, respectively. Indication of ≤50.0% RM reduction was absent. The 105, 106 and 107 broth culture dilutions of K. pneumoniae for all the eight tested antibiotics recorded RM reductions of 0 to 11.8, 0.0 to 6.3 and 5.3 to 30.8%, respectively with mean±SE RM reductions of 7.2±7.8, 1.1±11.7 and 23.3±7.4%, respectively. Again, there was absence of ≤50.0% RM reduction.
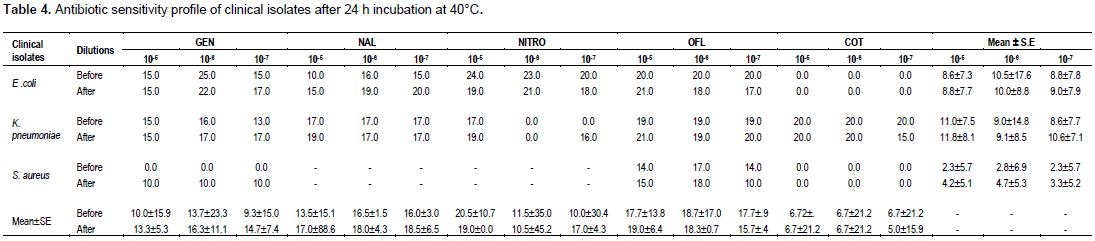
As was the case with 35°C temperature treatment, S. aureus at 40°C treatment recorded 0.0% RM reduction in 105, 106 and 107 broth culture dilutions for all the antibiotics tested apart from ofloxacin for which 7.1, 5.9, and 0.0% RM reductions were recorded for the three dilutions, respectively. Also, the mean ± SE RM reductions recorded for the three dilutions were however 121.7±5.4, 89.3±6.1, and 126.1±5.5%, respectively. All three dilutions produced greater than 50% RM reductions.
Broth culture dilutions (105, 106 and 107) of E. coli, K. pneumoniae and S. aureus with the application of gentamicin recorded 33.0±10.6, 19.0±17.25 and 58.1±11.25% RM reductions, respectively. The antimicrobial action of nalidixic acid on all three pathogens combined recorded 25.9±11.9, 9.1±2.95 and 15.6±4.3% RM reductions, respectively. Whereas the antibiotic activity of nitrofurantoin recorded 70.0±17.4% RM reduction at 107 dilution, ofloxacin recorded 7.3±10.1% RM reduction after 40°C temperature treatment of all three pathogens indications for ≤50.0% RM reduction were recorded for 107 dilution (58.1%) and 107 dilution (70.0%) with gentamicin and nitrofurantoin applications, respectively.
In Table 5, data recorded after 45°C temperature treatments are shown. E. coli broth culture dilution of 105 for all eight antimicrobials tested recorded 13.3 to 70.0% RM reduction and mean ± SE RM reduction of 57.0±14.3%. Broth dilutions of 106 and 107 recorded resistance marker (RM) reductions of 0.0 to 15.0% and 5.0 to 10.0%, respectively and mean±SE RM reductions of 22.9±24.3 and 37.5±7.3%, respectively. K. pneumoniae 105, 106 and 107 broth dilutions after 45°C temperature treatment recorded 6.7 to 10.5, 0.0 and 0.0 to 7.7% RM reductions, respectively with mean ± SE RM reductions of 0.0, 0.0 and 2.3±7.0%, respectively. S. aureus broth culture dilutions at 45°C treatment recorded 0.0% RM reduction for all antibiotics used apart from ofloxacin for which 0.0, 11.8 and 21.4% RM reductions and mean±SE reductions of 65.2±5.4, 71.4±6.9 and 95.7±6.05 were recorded, respectively for 105, 106 and 107 broth dilutions, respectively.
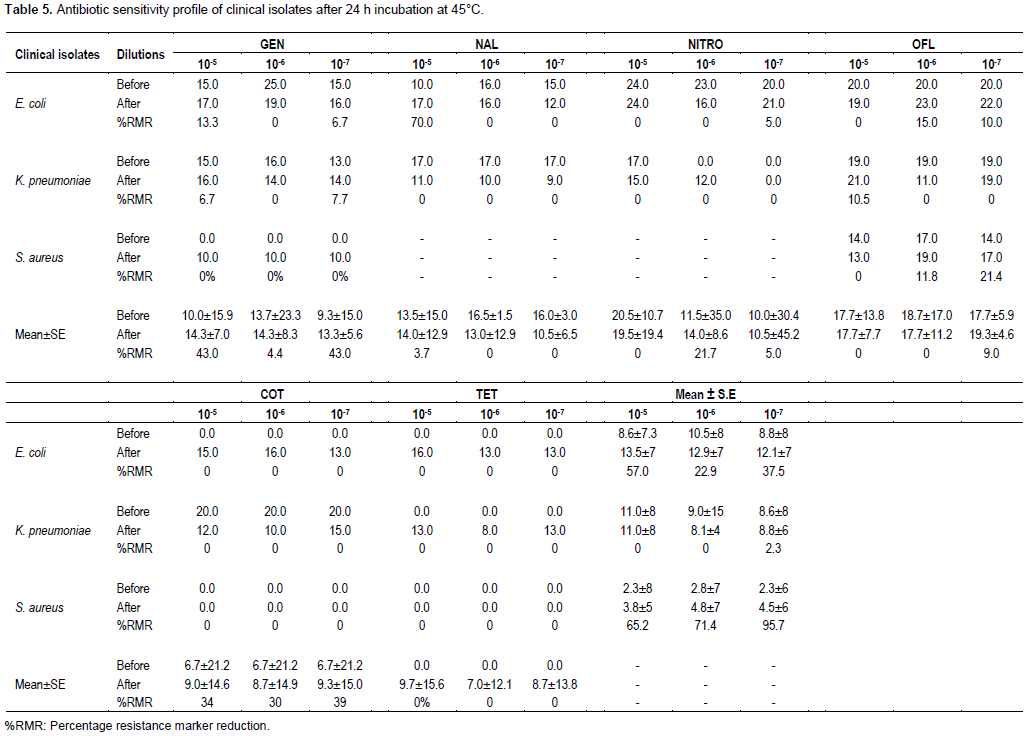
The 105, 106 and 107 broth dilutions of the three pathogens recorded 43.0±11.5, 4.4±15.8 and 43.0±10.3% RM reductions, respectively due to the antimicrobial action of gentamicin. Nalidixic acid activity produced RM reductions of 3.7±14.0, 0.0 and 0.0% for the three dilutions, respectively. RM reductions of 0.0, 21.7 and 5.0% were recorded, respectively for the three dilutions in that order as a result of the antibacterial activity of nitrofurantoin. The effect of ofloxacin was 9.0% RM reduction for 107 dilutions, while cotrimoxazole activity produced RM reductions of 34.3±17.9, 29.9±18.1 and 38.8±18.1% for 105, 106 and 107 broth culture dilutions, respectively of all three bacterial pathogens.
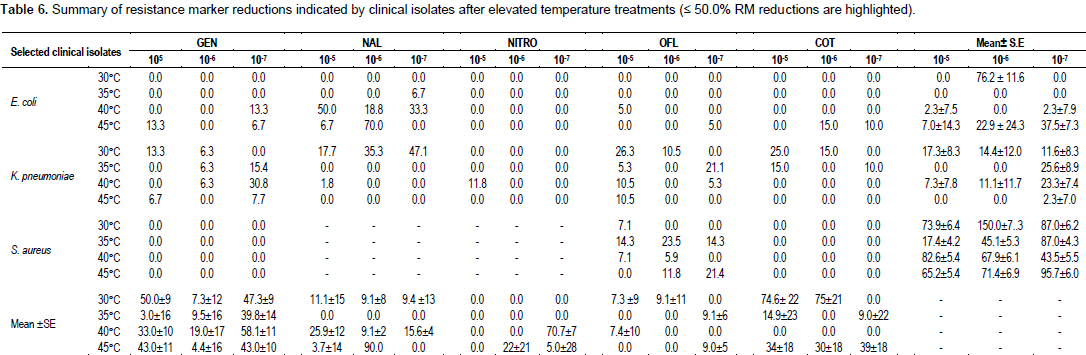
Table 6 shows the summary of resistance marker reduction at the various applied temperature treatments.
DISCUSSION
It has been reported that novobiocin, ethidium bromide, acriflavin, acridine orange, ascorbic acid and elevated temperatures are effective as curing agents (Ramesh et al., 2000). Elevated temperatures (43 to 44°C) have been successfully used not only to cure tetracycline resistant and penicillinase positive strains of S. aureus but also for curing F plasmids from E. coli (May et al., 1964). Also, it has been reported that certain physical treatments (such as elevated temperature treatment) and chemical compounds as well as growth conditions may increase the frequency of elimination of drug resistance R plasmids resulting in sensitive cells that were previously resistant to antibiotics (among other agents) have been used successfully to effect antibiotic sensitization of previously resistant bacteria (Chakrabartty et al., 1984; Gupta et al., 1980; Obaseki-Ebor, 1984; Reddy et al., 1986).
Any treatment that increases the frequency of elimination of plasmids will certainly enhance sensitivity of antibiotics. In this study, elevated temperatures of 30°C, 35°C, 40°C and 45°C were used to treat serially diluted pure broth cultures of E. coli, K. pneumoniae (both urinary tract pathogens) and S. aureus (a pathogen isolated from a wound infection). The plan was to see (in terms of resistance marker reduction or complete plasmids elimination), at what temperature(s) there was ≤50.0% RM reduction which is the significant benchmark (Akortha and Filgona, 2009). The three pathogens were selected simply based on their virulence and frequent occurrences in most human systemic and urinary infections. Hence, it may be reasonable to infer that patterns (treatment responses) recorded by these pathogens may be a general reflection of temperature treatment responses by other pathogens having same degree or higher degree of virulence.
Temperature treatments were carried out but in sterile nutrient broth medium and this was to enable the characteristic bacterial growth phases (of lag, log, stationary and decline phases) take their turn. Growth in liquid medium is synonymous with growth of selected bacterial pathogens and indeed, other bacterial pathogens in vivo (in the blood circulatory system, tissues and lymphatic system) of man and related primates. Incubation in water bath rather than conventional incubator was to ensure direct heat (temperature) contact with diluted broth cultures of pathogens selected for the study. Also, incubation for 18 h and not 24 h was to ensure that about 65% of cells in diluted broth cultures were in their actively growing (that is, viable or logarithmic) state to guarantee their optimal biochemical and physiological activities in the nutrient medium used.
Broth culture dilutions of 105, 106 and 107 (last three dilutions) were used for subculture after temperature treatment because the viable cells were probably well spaced out within their cultures to allow for maximal metabolic activities. Under 30°C temperature treatment, mean±SE resistance marker reduction recorded for 105, 106 and107 E. coli dilutions were 0.0, 76.2±11.6 and0.0%, respectively. For K. pneumoniae, it was 17.3±8.3, 14.4±12.0 and 11.6±8.3%, respectively and for broth dilutions of S. aureus, it was 173.9±6.4, 150.0±7.3 and 87.0±6.2%, respectively.
Significant RMr eductions were recorded for106 E. coli broth dilution and all three dilutions of S. aureus after 30°C treatment. The reason why at 106 dilution, E. coli recorded more than 50.0% RM reductions was not clear, but it may be because at that dilution, enzymatic activities increased above the specific temperature that is characteristic of the heat stability of the particular enzyme, thus enabling permeability of the tested drugs in line with the report of Hardy (1986).
Similarly, it was not clear why 105, 106 and 107 broth culture dilutions of S. aureus produced more than twice 50.0% RM reduction on average with 30°C temperature treatment. This is in view of the fact that more than 60.0% of resistance genes of S. aureus are chromosome-mediated and it has been reported that genes located on chromosomal DNA are not affected by temperature treatment compared to genes encoded by plasmid DNA (Radi and Rahman, 2010). Present findings is also not consistent with the report of Ochei and Kolhatkar (2008) which stated that Staphylococci displays a faster growth rate of resistant strains when incubated at 30°C rather than at 37°C. The present finding is open to validation. No significant RM reduction was recorded by K. pneumoniae dilutions suggesting that 30°C temperature treatment has no effect on the pathogen by other authors.
Temperature treatment of 35°C recorded 0.0% RM reduction for all three broth dilutions of E. coli and less than 30% (25.6±8.9%) RM reductions in 107 K. pneumoniae broth cultures dilutions. Only 107 S. aureus broth culture dilution yielded 87.0±4.3% RM reduction (more than 50.0%) due to the antimicrobial activity of all the drugs combined. Resistance marker reduction by as much as 87.0% implies that 35°C temperature treatment will enhance S. aureus susceptibility to gentamicin, ofloxacin, cotrimoxazole, tetracycline, erythromycin and augmentin (due probably to increased permeability to these drugs) when diluted to 107 in broth culture. Again, this finding is inconsistent with the report of Ochei and Kolhatkar (2008) which stated that Staphylococci displays a faster growth rate of resistant strains when incubated at 30°C than at 37°C.
Antimicrobial action of all the selected antibiotics and 40°C temperature treatment recorded less than 5.0% RM reduction on 105, 107 broth dilutions as well as 0.0% reduction on 106 dilution of E. coli. Less than 30% RM reduction was recorded for all broth culture dilutions of K. pneumoniae. Also at 40°C treatment, whereas more than 50.0% RM reduction was recorded separately for 105 and 106 broth dilutions of S. aureus, less than 50.0% reduction was recorded for 107 dilution. The increased (elevated) temperature treatment of 40°C did not yield a marked difference in resistance marker reduction compared to 35°C temperature treatment for E. coli and K. pneumoniae. This, ostensibly suggest that elevated temperature exposures of 35°C and 40°C have no effect on both pathogens in terms of RM reduction under the antibacterial activities of the selected antibiotics.
There was however significant RM reduction in broth dilutions of S. aureus. According to Degerman et al. (2013), temperature increase and addition of nutrients can shorten lag phase (response time) in a broth culture. This response time could be in terms of increased permeability of the selected antibiotics. Conversely, a decreased temperature can result in a prolonged lag phase (Buchanan and Klawitter, 1992; Kirchman and Rich, 1997). Kheder (2002) reported that at high temperature treatment between 40 and 46°C, microbial enzymes are sensitive to enzymes. Present finding is in agreement with the report of Hajdu et al. (2009) which established that antibacterial activity of antimicrobial agents against some strains of S. aureus was significant but differentially enhanced by increasing the ambient temperature. Incubation of S. aureus strains with antibiotics at 40°C or resulted in significant reductions in bacterial growth (Hajdu et al., 2009). A moderate increase in the ambient temperature increased the antibacterial activities of antibiotics such as daptomycin, vancomycin, tigecycline, fosfomycin and cefamandole (Hajdu et al., 2009).
Again, 82.6% and 67.9% (more than 50.0%) RM reductions in 105 and 106 S. aureus broth culture dilutions, respectively after 40°C temperature treatment reported in present study is not in tandem with the published work of Stanier et al. (1984) which stated that whereas elevated temperature of 40°C alone was ineffective in curing Pseudomonas putida, the plasmid or resistance marker of this organism was cured up to 2.63% when elevated temperature (40°C) and acridine orange combined were used.
At 45°C temperature treatment, the antibacterial action of all the selected antibiotics produced 57.0±14.3% (more than 50.0%) RM reduction in 105 E. coli broth culture dilution and less than 50.0% reduction in 106 and 107 broth dilutions. The 57.0% (more than 50.0%) RM reductions at 105 dilutions may be because E. coli attains optimal metabolic (catabolic and anabolic) activities at 44°C as well as increased antibiotic permeability into the cells may be responsible.
It was completely different with all three K. pneumoniae broth culture dilutions at 45°C temperature treatment. This was because 0.0% RM reduction was recorded for both 105 and 106 dilutions and less than 3.0% reduction in 107 dilution. Again, this may be suggestive that 45°C is not optimal for resistance marker reduction (that is, enhanced antibiotic sensitization) of multidrug resistant strains of the clinical pathogen.
Resistance marker reductions of 65.2±5.4, 71.4±6.9 and 95.7±6.0% (all more than 50.0%) were recorded for 105, 106 and 107 S. aureus broth culture dilutions, respectively after 45°C temperature treatment. This finding is supported by Bondi et al. (1958) whose report stated that growth of S. aureus at 45°C increased the rate of loss of penicillinase production. The present finding is also consistent with the report that growth at elevated temperature (44°C) increased the rate of loss of penicillinase resistance in some strains of Staphylococci (Fairbrother et al., 1954). May et al. (1964) showed that loss at both penicillin and tetracycline resistance during growth of a strain of S. aureus occurred at 43°C to 44°C. Some authors have reported that 46°C temperature treatment of three strains of P. aeruginosa efficiently cured all the plasmids being harboured within the genome of these organisms and conceded that elevated temperature (46°C) is more efficient to cure all plasmids than chemical agents (Radi and Rahman, 2010).
Antimicrobial action of gentamicin on 105 and 106 broth culture dilutions of E. coli, K. pneumoniae and S. aureus combined yielded 50.0±9.8 and 58.1±11.2% RM reduction at 30°C and 40°C temperatures treatment, respectively. Less than 50.0% reduction was recorded at the other temperature treatment. The antibacterial activity of nalidixic acid did not yield up to 30.0% RM reduction in all broth dilutions of the pathogens at the tested elevated temperatures. A similar situation (to nalidixic acid) occurred with ofloxacin. The antibacterial action of nitrofurantoin however, produced a 70.7±17.4% (more than 50.0%) RM reduction (at 40°C) in 107 broth culture dilution of the pathogens used in the study. The action of cotrimoxazole produced 74.6±22.2 and 74.6±21.0% reductions in 105 and 106 broth dilutions, respectively of the pathogens at 30°C temperature treatment In all, no single antibiotic action on all three pathogens produced significant (≤50.0%) RM reduction after 45°C temperature treatment.
CONCLUSION
Elevated temperature exposures of 30 and 40°C of pure broth culture of E. coli can produce significant (≤50.0%) resistance marker reduction under the activity of selected narrow and broad spectrum antibiotics. None of the tested elevated temperature (that is, 30°C, 35°C, 40°C and 45°C) seemed to have any effect on pure broth culture of K. pneumoniae as no significant (≤50.0%) RM reduction was recorded. All tested elevated temperatures produced significant (≤50.0%) RM reduction in S. aureus pure broth culture under the action of the selected antimicrobials. Individually, gentamicin action yielded significant RM reduction on broth cultures of all three pathogens at 30 and 40°C temperature treatments. Nitrofurantoin activity yielded significant RM reduction at 40°C, while cotrimoxazole action resulted in significant reduction at 30°C.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Akortha EE, Filgona J (2009). Transfer of gentimicin resistance genes among enterobacteriaceae isolated from the outpatients with urinary tract infections attending 3 hospitals in Mubi, Adamawa State. Sci. Res. Essays 4(8):745-752. |
|
|
Asheshov EH (1966). Loss of antibiotic resistance in Staphylococcus aureus resulting from growth at high temperature. J. Gen. Microbiol. 42(3):403-410. |
|
|
Asheshov EH (1966). Chromosomal location of the genetic element controlling Penicillinase production in a strain of Staphylococcus aureus. Nature 210(5038):804-806. |
|
|
Bauer AW, Kirby WM, Sherris JC, Turk M (1966). Antibiotic susceptibility testing by a standardized single disc method. Am. J. Clin. Pathol. 45:493-496. |
|
|
Buchanan RL, Kawitter LA (1992). The effect of incubation temperature,Initial pH and sodium chloride on the growth kinetics of Escherichia coli 0157:h7. Food Microbiol. 9:185-196. |
|
|
Chakrabartty PK, Mishra AK, Chakrabarti SK (1984). Loss of plasmid Linked drug resistance after treatment with iodo-deoxyuridine. Indian J. Exp. Biol. 22:333-334. |
|
|
Degerman R, Dinasquet J, Riemann L, de Luna SS, Andersson A (2013). Effect of resource availability on bacterial community responses to increased temperature. Sci. Rep. 68:131-142. |
|
|
Fairbrother RW, Parker L, Eaton BR (1954). The stability of penicillinase producing strains of Staphylococcus aureus. J. Gen. Microbiol. 10:309-314. |
|
|
Goller C, Romeo T (2008). Environmental influences on biofilm development. Curriculum Top. Microbiol. Immunol. 322:37-66. |
|
|
Gupta TD, Bandyopathy T, Dastidar SG, Bandopadhyay M, Mistra A, Chakrabarty AN (1980). R plasmids of Staphylococcus and their Elimination by different agents. Indian J. Exp. Biol. 18:478-481. |
|
|
Hajdu S, Lassnigg A, Graninger W, Hirschl AM, Presterl E (2009). Effect of vancomycin, daptomycin, fosfomycin, tigecycline and ceftriaxone on Staphylococcus epidermidis biofilms. J. Orthop. Res. 27:136-1365. |
|
|
Hardy K (1986). Bacterial plasmids. Second edition. American Society for Microbiology. 1913 street N, W. Washington D.C. (20006) USA. |
|
|
Kheder AK (2002). Studies on antibiotic resistance by plasmid of Pseudomonas aeruginosa Ph.D. thesis. College of Education. University of Salahaddin, Irag. |
|
|
Kirchman DL, Rich JH (1997). Regulation of bacterial growth rates by Dissolved organic carbon and temperature in the equatorial Pacific Ocean. Microb. Ecol. 33:11-20. |
|
|
Lakshmi VV, Padma S, Polasa H (1989). Loss of plasmid antibiotic resistance in Escherichia coli on treatment with some compounds. FEMS Microbiol. Lett. 57:275-278. |
|
|
Madigan M, Martinko J, Parker J (2003). Brock biology of microorganisms (10th edition). Prentice Hall, Upper Saddle River, NJ. USA. 500 p. |
|
|
May JW, Houghton RH, Perret CJ (1964). The effect of growth at elevated temperatures on some heritable properties of Staphylococcus aureus. J. Gen. Microbiol. 2:56-61. |
|
|
McGowan AP, Wise R (2001). Establishing MIC breakpoints and the Interpretation of invitro susceptibility tests. J. Antimicrob. Chemother. 48:17-28. |
|
|
Neu HC (1989). Overview of mechanisms of bacterial resistance. Diagn. Microbiol. Infect. Dis. 12:109-116. |
|
|
Obaseki-Ebor EE (1984). Rifampicin curing of plasmids in Escherichia coli k12 rifampicin resistant host. J. Pharm. Pharmacol. 36:467-470. |
|
|
Ochei J, Kolhatka A (2008). Medical Laboratory Science Theory and Practice. 10th Edn. New Delhi: Tata Mc Graw hill Publishing Company Limited. 255 p. |
|
|
Okeke IN, Lamikanra A, Edelman R (1999). Socio economic and behavioural factors leading to acquired bacterial resistance to antibiotics in developing countries. Emerg. Infect. Dis. 5:18-27. |
|
|
Oskay M, Oskay D, Kalyoneu F (2009). Activity of some plant extracts against multidrug resistant human pathogens. Iran. J. Pharm. Res. 8(4):293-300. |
|
|
Rachid S, Ohlsen K, Wallner U, Hacker J, Hecker M, Ziebuhr W (2000). Alternative transcription factor is involved in regulation of biofilms. Bacteriology 182:6824-6826. |
|
|
Radi OR, Rahman FH (2010). Study of the effects of ethidium bromide, SDS and elevated temperature on stability of multiple antibiotic resistance plasmids of Pseudomonas aeruginosa. Iraqi J. Biotechol. 9(4):797-811. |
|
|
Ramesh A, Heami PM, Chandrasekhar A (2000). Ascorbic acid induced loss of a pediocin- encoding plasmid in Pediococus acidilactivi CFR K7. World J. Microbiol. Biotechnol.16:695-697. |
|
|
Reddy G, Shridhar P, Polasa H (1986). Elimination of Col. El (PBR322) and (PBR329) plasmids in Escherichia coli on treatment with hexamine ruthenium III chloride. Curr. Microbiol. 13:243-246. |
|
|
Stanier RY, Adelberg EA, Ingraham JL (1984). General Microbiology. 4th edition. The Placmilian Press LTD, London and Basingstoke. |
|
|
Trevors JT (1986). Plasmid curing in bacteria. FEMS Microbiol. Rev. 32:149-152. |
|
|
Viljanen P, Boratynski J (1991). The susceptibility of conjugative resistance transfer in gram negative bacteria to physiochemical and biochemical agents. FEMS Microbiol. Rev. 88:43-45. |
|
|
Zhao BY, Hu KA, Wu RJ (2001). Primary study on the phenollic modification of sodium lignosulphate. Polym. Mater. Sci. Eng. 16(1):158-161. |
|
|
Zielenkiewicz U, Ceglowski P (2001). Mechanisms of plasmids stable Maintainance with special focus on plasmid addiction system. Acta Biochem. Pol. 48(4):1003-1023. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


