Full Length Research Paper

ABSTRACT
Piper chaba is a flowering vine in the family of piperaceae. It is a spicy plant widely used as food and herbal medicine in the South and Southeast Asian countries. The study evaluated anti-inflammatory and immunomodulatory activities of a major alkaloid extract, piperine, from P. chaba fruits on human colonic epithelial cells. The effects of piperine extract on cell viability, pro-inflammatory cytokine inhibition, and immunomodulation on inflammatory genes were evaluated in vitro using lipopolysaccharide (LPS)-induced inflammatory model in Caco-2 cells. T-test and ANOVA were used for statistical analysis. Piperine, in the doses of non-cytotoxicity (1, 10, 50 µg/ml), reversed effect of LPS on pro-inflammatory cytokines; tumor necrosis factor (TNF)-α, interleukin (IL)-1β, and interleukin (IL)-6 production. In addition, it also confirmed the activity on mediating IκB-α/NF-κB signaling proteins by activating IκB-α and suppressing NF-κB expression leading to attenuate pro-inflammatory cytokine release during the inflammatory process. The study concludes that piperine from P. chaba inhibits LPS-induced inflammation in intestinal epithelial cells by modulating cytokine gene expression via down regulation of IκB-α/NF-κB signaling pathway.
Key words: Piperine, Pro-Inflammatory Cytokine, Iκb-Α, Nf-Κb, Caco-2 Cells.
INTRODUCTION
Inflammation is part of the body reaction to invasive stimuli. The immune cells are responsible for inflammatory response by such as macrophages, neutrophils, lymphocytes and epithelial cells (Cheng et al., 2018). When the host is stimulated by micro-organisms or tissue injury, the guard cells react on acute-phase response by releasing acute-phase proteins including serum amyloid, chemokines, mediators, or inflammatory cytokines, to play a concert in inflammatory process leading to several systemic responses (Buamann and Gauldie, 1994; Laskin and Pendino, 1995; Gabay and Kushner, 1999). The significant pro-inflammatory cytokines or mediators participating at the beginning of inflammatory process are tumor necrosis factor (TNF)-α, interleukin (IL)-1β, and interleukin (IL)-6, nitric oxide (NO) or prostaglandin (PG) E2 (Buamann and Gauldie, 1994; Gabay and Kushner, 1999).
Gastrointestinal (GI) epithelia also play an important role in immunity because they are the first line of barrier against external environment, the GI content, located in our body. The ingested foods, as an external environment, contain invasive pathogens, chemicals, and toxins that can produce inflammation (Chen et al., 2006). These inflammatory stimulators activate innate and acquired immune response from immune cells residing in the gut wall, or intestinal epithelial cells lining intestinal mucosa, resulting in mucosal inflammation. If an inflammation is not removed, it will bring the situation into inflammatory disease such as inflammatory bowel diseases (IBD) which is caused by pathogens, non-pathogens, or autoimmune disorders (Chakravortty and Kuma, 1999; Panja et al., 1998). For bacterial cause, lipopolysaccharides (LPS) are inflammatory factors derived from outer part of gram-negative bacteria and are active parts to stimulate inflammation. Therefore, They are usually used in the study of inflammatory mechanism in in vitro model (Choi et al., 2014).
Plants are naturally potential sources of medicinal agents nowadays. Previous studies showed many beneficial effects of plants as herbal medicine and as natural sources of pharmaceutical active compounds. For herbal medicine, they can be potentially used as crude of both a single plant or incorporated into herbal recipe. Piper chaba is a flowering vine in the family of piperaceae. It is a spicy plant of pepper family which is widely used as food and herbal medicine in South and Southeast Asian countries. It is a creeper plant spreading on the ground or growing around the large trees. The fruit looks similar to long pepper. It turns red when ripe and turns dark brown or black when dry. In Thailand, P. chaba is called ‘Dee Plee”. The stem, root, and fruit of Dee Plee, are commonly used in the recipe of folk medicine (Islam et al., 2020).
Piperine (Figure 1) is a major compound derived from natural herbs such as curcumin, P. chaba and Piper nigrum (black pepper) (Naz et al., 2012). These pepper plants are usually used as spice in foods and as phytomedicine. As a dietary medicine, piperine extract has been shown to exhibit many pharmaceutical effects such as anti-oxidant (Prakash and Srinivasan, 2010; Rauscher et al., 2000) anti-inflammation (Srinivasan, 2007; Tasleem et al., 2014), antispasmodic activity (Doucette et al., 2013; Mehmood and Gilani, 2010), anti-angiogenesis (Lu et al., 2012), antimalarial activity (Thiengsusuk et al., 2018), anti-diarrhea by inhibiting cAMP-mediated Cl- secretion in human colonic epithelial cells (Pongkorpsakol et al., 2015), and prevention of gallstones (Li et al., 2015a). However, the role of piperine on anti-inflammation on human intestinal epithelial cells has not been reported. The study evaluated an effect of piperine extracted from P. chaba fruits on LPS-induced pro-inflammatory cytokine release and the immunomodulation in Caco-2 cells (Figure 1).
MATERIALS AND METHODS
Plant extraction
P. chaba fruits were collected, identified, and extracted by the Thai Medicine Development Center, National Science and Technology Development Agency (NSTDA), Pathum Thani, Thailand, 12120. Piperine extraction was done using high performance liquid chromatography (HPLC). The yellow crystals of piperine extract were done for quantitative analysis and the purification obtained is 95%.
Chemicals
Dimethyl sulfoxide (DMSO) was purchased from Sigma-Aldrich (St. Louis, MO, USA). MTT (3-(4,5-dimethylthaizol-2-yl)-2,5-diphenyltetrazolium bromide) was purchased from Invitrogen Thermo Fisher Scientific (Waltham, MA, USA). Materials for cell culture were purchased from Gibco-BRL (Grand Island, NY, USA). Enzyme-linked immunosorbent assay (ELISA) kits for TNF-α, IL-1β, and IL-6 quantification were brought from Merck Millipore (Etobicoke, Ontario, Canada). InstantOne TM ELISA kits for inhibitory kappaB (IκB)-α and nuclear factor-kappaB (NF-κB) (p65) analysis were purchased from Affymetrix eBioscience (Waltham, MA, USA).
Cell culture
Caco-2 cells were purchased from American Type Culture Collection (ATCC) (Manassas, VA, USA). They were cultured in dulbecco’s modified eagle medium (DMEM)/F12 containing 10% fetal bovine serum (FBS) (Gibco-BRL, Grand Island, NY, USA), penicillin (100 U/ml) and streptomycin (100 µg/ml) at 37°C in a humidified atmosphere of 5% CO2. Piperine extract was dissolved in DMSO and was diluted to stock concentrations with free FBS culture medium. Cells were incubated with piperine in various concentrations and were stimulated with LPS for the incubation time.
Cell viability assay
The cell viability was measured based on MTT colorimetric assay. Caco-2 cells of 1×104 cells/ml were grown in 96-well plates until confluence. Fresh media were added and the cells were further incubated at 37°C with piperine (0.1, 1, 10, 50 and 100 µg/ml) for 8 h in a humidified atmosphere of 5% CO2. At the end of the incubation time, fresh medium and tetrazolium dye (MTT) solution (5 mg/ml) were added. The cells were further incubated for 4 h at 37°C in a humidified atmosphere of 5% CO2. A 200 µl aliquot of DMSO was added and the absorbance at 540 nm was measured in a Bio Tex Power Wave XS automatic microplate reader (Bio Tex Instrument Inc., Winooski, VT, USA).
Pro-inflammatory cytokine determination
Caco-2 cells of 5×105 cells/ml were grown in 24-well plates until they reached confluence. After that, they were incubated with various concentration of piperine extract (1, 10, and 50 µg/ml) in an absence or presence of LPS (1 µg/ml) for 24 h at 37°C in a humidified atmosphere of 5% CO2. Supernatant fluids were collected and stored at -80°C until cytokines were analyzed. The pro-inflammatory cytokines were determined by using commercial human TNF-α, IL-1β and IL-6 ELISA kits according to the manufacturer’s instructions.
Signaling protein determination
Caco-2 cells of 1×104 cells/ml were seeded in 96-well plate and were incubated with only LPS (1 µg/ml) and LPS plus various concentrations of piperine (1, 10, and 50 µg/ml) for 24 h at 37°C in a humidified atmosphere of 5% CO2. The cells were washed with phosphate buffer saline (PBS) and then lysed with lysis buffer included with InstantOne TM ELISA kits. The signaling proteins of IκB-α and NF-κB (p65) were determined by using commercial human InstantOne TM ELISA kits according to the manufacturer’s instructions. The absorbance at 450 nm for O.D. values was measured in a Bio Tex Power Wave XS automatic microplate reader (Bio Tex Instrument Inc., Winooski, VT, USA).
Statistical analysis
Data from at least three individual experiments in each protocol were analyzed, and are presented as mean±SEM. Statistical significance was determined using one-way ANOVA and student t-test. P value less than 0.05 is statistically significant.
RESULTS
Cytotoxic effect of piperine
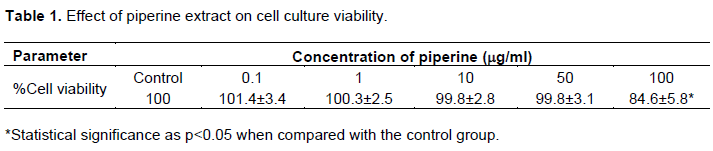
To study the cytotoxicity of piperine extract on culture cells, the cells were treated with progressively increase in piperine concentration (0.1, 1, 10, 50,100 µg/ml). Cell survival was evaluated by MTT calorimetric assay. The results are shown in Table 1. Piperine extract in the doses of 0.1 to 50 µg/ml shows survival rate of nearly 100% compared to non-treated control group, while the concentration of 100 µg/ml reveals significantly decreased cell viability.
Effect of piperine on LPS-induced pro-inflammatory cytokine production
Piperine extract, in the concentration of non-cytotoxicity (0.1, 1, 10, 50 µg/ml), is further used to evaluate the effect on pro-inflammatory cytokine production. The concentrations of TNF-α, IL-1β, and IL-6 were measured in cell culture media of LPS-treated and non-treated groups after 24 h incubation by ELISA method. The cells were divided into four groups: Group 1, growing without both LPS and piperine; Group 2, growing with only LPS (1 µg/ml); Group 3, growing with only piperine in various concentrations; and Group 4, growing with LPS plus piperine of non-cytotoxic concentrations. The results demonstrate that piperine alone has no effect on pro-inflammatory cytokine stimulation; however, LPS alone acts as a potent inflammatory stimulator by activating pro-inflammatory cytokine release. Piperine extract clearly reveals a protective effect on inflammation by suppressing pro-inflammatory cytokine release during LPS stimulation in a dose-dependent manner (Figure 2).

Effect of piperine on LPS-induced signaling protein, expression
LPS stimulates inflammatory response via NF-κB/Rel family signaling proteins. In the resting state, NF-κB forms dimer with IκB-α and locates in the cytosol. During LPS activation, IκB-α dissociates from NF-κB and is degraded in the cytosol while NF-κB moves into the nucleus to stimulate various inflammatory genes. To investigate the role of piperine extract on cellular mechanism of pro-inflammatory cytokine production, IκB-α and NF-κB expression was evaluated using instant One ELISA which the principle is close to semi-quantitative analysis of western blot. The levels of signaling proteins, IκB-α and NF-κB, in various groups of study are shown in Figure 3 as O.D. values compared with those of positive control of instant One ELISA.
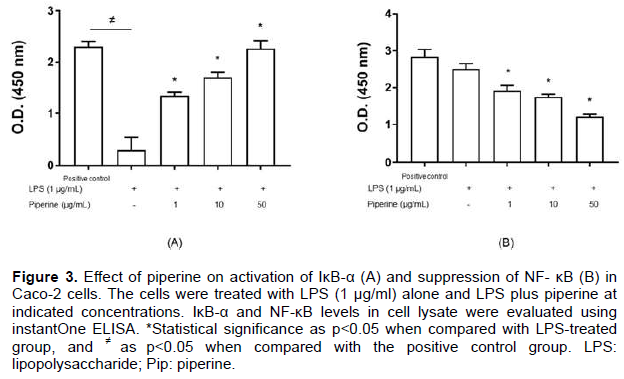
LPS alone significantly suppressed IκB-α expression in Caco-2 cells. However, the concentrations of cytosolic IκB-α were significantly resumed in a concentration-dependent manner when the cells were added with piperine extract. An expression of NF-κB, on the other hand, was exponentially increased when the cells were incubated with only LPS. This protein levels were significantly reduced in a concentration-dependent manner when the cells were co-cultured with piperine extract.
DISCUSSION
Our body has defensive mechanisms to protect ourselves from aggressive invaders such as pathogens, toxins, and chemicals. This is an innate immunity that attempts to remove attackers and to initiate healing process known as inflammation which is characterized by pain, swelling, redness, and hot. In the GI tract, the innate immunity is responsible by immune cells as well as epithelial cells lining the gut (Neurath et al., 1998). When the body is stimulated by pathogenic stimuli, these guard cells release chemokines such as leukotriene and histamine, inflammatory mediators such as prostaglandins (PGEs) and (NO), or inflammatory cytokines e.g. TNF-α, IL-1β, or IL-6 to act on inflammatory process. Interleukin-1β stimulates the expression of adhesive molecules on endothelial cells to allow the migratory action of leukocytes to an inflammatory site and also involves with hyperalgesia and fever (Jeong et al., 2014). TNF-α promotes the production of other inflammatory cytokines to join an inflammation that can lead to autoimmune disorders (Guadagni et al., 2007). The intestinal mucosa, as the first line of bodily protection, encounters toxins from pathogens which commonly cause inflammation. Chronic or recurrent inflammation, can cause inflammatory diseases.
There are documents supporting that the important pathways regulating inflammatory response in several cell types are MAPKs and IκB-α/NF-κB (Ying et al., 2013a, b). LPS, provokes an immune response in host cells via mediating membrane receptor protein such as Toll-like receptor 4 (TLR4) (Palsson-McDermott and O'Neill, 2004). This protein receptor passes the signal through cytosolic signaling proteins such as NF-κB/Rel family to the nucleus and activates several genes of inflammation and necrosis (Hayden et al., 2006; Beinke and Ley, 2004). Suppression on these genes has been accepted to become a good way for curing inflammatory diseases (Salminen et al., 2012; Nam, 2006). The nuclear protein NF-κB modulates the inflammatory genes by coordinating with cytosolic inhibitory protein, IκB-α. During unstimulated state, NF-κB protein forms dimer to IκB-α in the cytosol. When LPS binds to the membrane receptor and the cell is stimulated, IκB-α is phosphorylated and dissociates from NF-κB to the process of degradation (Hayden et al., 2006; Choi et al., 2014). The free NF-κB moves into the nucleus as transcription factor, binds to the target genes, and stimulates inflammatory cytokine release. Therefore during inflammation, the level of IκB-α could decrease and the level of NF-κB could increase.
Piperine is an alkaloid substance found in several medicinal plants such as, curcumin, black pepper, and long pepper (Yaffe et al., 2015). It has been proposed to have many therapeutic effects in traditional medicine especially anti-inflammation (Li et al., 2015b). The effect of piperine extract from P. chaba fruits on LPS-stimulated inflammation was investigated and its activity on the expression of inflammatory signaling proteins was evaluated. The results show that piperine extract attenuated LPS-induced pro-inflammatory cytokine releases in a concentration-dependent manner. In addition, during an inflammatory process (stimulated by LPS), piperine extract modulated the signaling proteins by elevating the levels of IκB-α, which is suppressed when LPS presents, and decreases levels of and decreasing the levels of NF-κB, which is stimulated by LPS.
These findings confirm that LPS stimulates inflammatory gene expression as shown in increased pro-inflammatory cytokine levels, compared to LPS-untreated cells. Piperine extract can inhibit LPS action by modulating IκB-α level during inflammatory process. This allows less number of free NF-κB to stimulate inflammatory genes as shown in low levels of pro-inflammatory cytokine expression. The results of the present study are supported by the studies on murine macrophage models showing that piperine inhibited LPS-induced PGE2, NO, TNF-α, and cox II gene expression by inhibiting NF-κB nuclear translocation (Ying et al., 2013a; Kim et al., 2012).
In Caco2 cells, piperine shows reverse effect of LPS on IκB-α and NF-κB expression. This may be due to piperine, has inhibitory action on the process of IκB-α phosphorylation and degradation. This leads to sustain a high level of this protein in the cytosol and restrains NF-κB transcription factor in an inactive form. As stated by the previous studies, piperine inhibited IκB-α phosphorylation and NF-κB nuclear localization (Jeong et al., 2014; Ying et al., 2013b). Taken together, in Caco-2 cells, piperine shows the same effects as in murine macrophage because it may also have an inhibitory effect on IκB-α phosphorylation. This makes the two signaling proteins stay as dimer in the cytosol. Therefore, the inflammatory genes are not activated if piperine is present. The present study gives scientific support to medicinal plants containing piperine on curing inflammatory diseases, especially the inflammatory diseases in the GI tract such as peptic ulcer and IBD. In IBD, an inhibition of NF-κB activation has been suggested as a treatment strategy (Dijkstra et al., 2002). The results of the present study could support the natural source of active compound for serving that treatment.
CONCLUSION
The findings of the study confirm that piperine from P. chaba fruits can inhibits LPS-induced gastrointestinal epithelial cell inflammation by reversing LPS effect on inflammatory gene expression. This would suggest that P. chaba fruits could be further developed as a biomedicine for curing inflammatory diseases of the GI tract. It could benefit supplementary natural product to reduce the use of synthetic steroid drugs that have several side effects. However, further study as a use in medicine is still needed.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENT
The author appreciates Thammasat University, Pathum thani, Thailand, for funding the research.
REFERENCES
|
Buamann H, Gauldie J (1994). The acute phase response. Immunology Today 15(2)74-80. |
|
|
Beinke S, Ley SC (2004). Functions of NF-kappaB1 and NF-kappaB2 in immune cell biology. The Biochemical Journal 382(Pt 2):393-409. |
|
|
Chakravortty D, Kumar KS (1999). Interaction of lipopolysaccharide with human small intestinal lamina propria fibroblasts favors neutrophil migration and peripheral blood mononuclear cell adhesion by the production of proinflammatory mediators and adhesion molecules. Biochimica et Biophysica Acta 1453(2):261-272. |
|
|
Chen J, Ng CP, Rowlands DK, Xu PH, Gao JY, Chung YW, Chan HC (2006). Interaction between enteric epithelial cells and Peyer's patch lymphocytes in response to Shigella lipopolysaccharide: effect on nitric oxide and IL-6 release. World Journal of Gastroenterology 12(24):3895-3900. |
|
|
Cheng N, Yurong L, Du X, Ye RD (2018). Serum amyloid A promotes LPS clearance and suppresses LPS-induced inflammation and tissue injury. EMBO Report 19:e45517. |
|
|
Choi WK, Jeong JW, Kim SO, Kim GY, Kim BW, Kim CM, Seo YB, Kim WY, Lee SY, Jo KH, Choi YJ, Choi YH, Kim GD (2014). Anti-inflammatory potential of peat moss extracts in lipopolysaccharide-stimulated RAW 264.7 macrophages. International Journal of Molecular Medicine 34(4):1101-1109. |
|
|
Dijkstra G, Moshage H, Jansen (2002). Blockade of NF-kappaB activation and donation of nitric oxide: new treatment options in inflammatory bowel disease? Scandinavian Journal of Gastroenterology (Suppl) 236:37-41. |
|
|
Doucette CD, Hilchie AL, Liwski R, Hoskin DW (2013). Piperine, a dietary phytochemical, inhibits angiogenesis. The Journal of Nutritional Biochemistry 24(1):231-239. |
|
|
Gabay C, Kushner I (1999). Acute-phase proteins and other systemic responses to inflammation. New English Journal of Medicine 340(6):448-454. |
|
|
Guadagni F, Ferroni P, Palmirotta R, Portarena I, Formica V, Roselli M (2007). TNF/VEGF cross-talk in chronic inflammation-related cancer initiation and progression: An early target in anticancer therapeutic strategy. In Vivo 21(2):147-161. |
|
|
Hayden MS, West AP, Ghosh S (2006). NF-kappaB and the immune response. Oncogene 25(51):6758-6780. |
|
|
Islam MT, Hasan J, Snigdha HMSH, Ali ES, Sharifi-Rad J, Martorell M, Mubarak MS (2020). Chemical profile, traditional uses, and biological activities of Piper chaba Hunter. Journal of Ethnopharmacology 16(257):112853. |
|
|
Jeong JW, Lee HH, Han MH, Kim GY, Hong SH, Park C, Choi YH (2014). Ethanol extract of Poria Cocos reduces the production of inflammatory mediators by suppressing the NF-kappaB Signaling Pathway in lipopolysaccharide-stimulated RAW 264.7 macrophages. BMC Complementary and Alternative Medicine 14:100. |
|
|
Kim HG, Han EH, Jang WS, Choi JH, Khanal T, Park BH, Tran TP, Chung YC, Jeong HG (2012). Piperine inhibits PMA-induced cyclooxygenase-2 expression through downregulating NF-kappaB, C/EBP and AP-1 signaling pathways in murine macrophages. Food and Chemical Toxicology 50(7):2342-2348. |
|
|
Laskin DL, Pendino KJ (1995). Macrophages and inflammatory mediators in tissue injury. Annual Review of Pharmacology and Toxicology 35:655-677. |
|
|
Li Y, Li K, Hu Y, Xu B, Zhao J (2015a). Piperine mediates LPS induced inflammatory and catabolic effects in rat intervertebral disc. International Journal of Clinical Experimental Pathology 8(6):6203-6213. |
|
|
Li Y, Li M, Wu S, Tian Y (2015b). Combination of curcumin and piperine prevents formation of gallstones in C57BL6 mice fed on lithogenic diet: whether NPC1L1/SREBP2 participates in this process? Lipids in Health and Disease 14:100. |
|
|
Lu JJ, Bao JL, Chen XP, Huang M, Wang YT (2012). Alkaloids isolated from natural herbs as the anticancer agents. Evidence-based Complementary and Alternative Medicine 2012:485042. |
|
|
Mehmood MH, Gilani AH (2010). Pharmacological basis for the medicinal use of black pepper and piperine in gastrointestinal disorders. Journal of Medicinal Food 13(5):1086-1096. |
|
|
Nam NH (2006). Naturally occurring NF-kappaB inhibitors. Mini-Reviews in Medical Biochemistry 6(8):945-951. |
|
|
Naz T, Mosaddik A, Rahman M, Muhammad I, Haque E, Cho SK (2012). Antimicrobial, antileishmanial and cytotoxic compounds from Piper chaba. Natural Product Research 26(11):979-986. |
|
|
Neurath M, Becker C, Barbulescu K (1998). Role of NF-KB in immune and inflammatory response in the gut. Gut 43(6):856-860. |
|
|
Palsson-McDermott EM, O'Neill LA (2004). Signal transduction by the lipopolysaccharide receptor, Toll-like receptor-4. Immunology 113(2):153-162. |
|
|
Panja A, Goldberg S, Eckmann L, Krishen P, Mayer L (1998). The regulation and functional consequence of proinflammatory cytokine binding on human intestinal epithelial cells. Journal of Immunology 161(7):3675-3684. |
|
|
Prakash UN, Srinivasan K (2010). Gastrointestinal protective effect of dietary spices during ethanol-induced oxidant stress in experimental rats. Applied Physiology Nutrition and Metabolism 35(2):134-141. |
|
|
Pongkorpsakol P, Wongkrasant P, Kumpun S, Chatsudthipong V, Muanprasat C (2015). Inhibition of intestinal chloride secretion by piperine as a cellular basis for the anti-secretory effect of black peppers. Pharmacological Research 100:271-280. |
|
|
Rauscher FM, Sanders RA, Watkins JB (2000). Effects of piperine on antioxidant pathways in tissues from normal and streptozotocin-induced diabetic rats. Journal of Biochemical and Molecular Toxicology 14(6):329-334. |
|
|
Salminen A, Kauppinen A, Kaarniranta K (2012). Phytochemicals suppress nuclear factor-κB signaling: impact on health span and the aging process. Current Opinions in Clinical Nutrition and Metabolism Care 15(1):23-28. |
|
|
Srinivasan K (2007). Black pepper and its pungent principle-piperine: A review of diverse physiological effects. Critical Reviews in Food Science and Nutrition 47(8):735-748. |
|
|
Tasleem F, Azhar I, Ali SN, Perveen S, Mahmood SA (2014). Analgesic and anti-inflammatory activities of Piper nigrum L. Asian Pacific Journal of Tropical Medicine 7S1:S461-S468. |
|
|
Thiengsusuk A, Muhamad P, Chaijaroenkul W, Na-Bangchang K (2018). Antimalarial activity of piperine. Journal of Tropical Medicine 6:9486905. |
|
|
Yaffe PB, Power CMR, Doucette CD, Walsh M, Hoskin DW (2015). Piperine, an alkaloid from black pepper, inhibits growth of human colon cancer cells via G1arrest and apoptosis triggered by endoplasmic reticulum stress. Molecular Carcinogenesis 54(10):1070-1085. |
|
|
Ying X, Chen X, Cheng S, Shen Y, Peng L, Xu HZ (2013a). Piperine inhibits IL-beta induced expression of inflammatory mediators in human osteoarthritis chondrocyte. International Immunopharmacology 17(2):293-299. |
|
|
Ying X, Yu K, Chen X, Chen H, Hong J, Cheng S, Peng L (2013b). Piperine inhibits LPS-induced expression of inflammatory mediators in RAW 264.7 cells. Cellular Immunology 285(1-2):49-54. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


