Full Length Research Paper

ABSTRACT
Mexico is the fourth largest producer of mango and the leading exporter worldwide, and cv ‘Ataulfo’ is the largest area sown in the country. New cultivars are required to maintain production, ensure competitiveness, and mitigate problems with pests and diseases. The purpose of the present study was to identify microsatellite (SSR) markers in zygotic and nucellar seedlings originated by the largest embryo in mango seeds, cultured in vitro. DNA was extracted from the foliar tissue of 18 seedlings from four different populations, and its origin was identified with six SSRs: MIAC-4, MIAC-5, mMiCIR003, mMiCIR030, LMMA1, and LMMA9. The number of alleles found varied from two to three, and allele size ranged from 95 to 325 bp. Based on the polymorphism taking place in microsatellites and genetic similarity index (GSI) data, more than 15% of the analyzed seedlings were found to be different from the female parent. Based on Nei’s genetic distance and UPGMA cluster analysis, the genetic variability among the four populations was 6.2%.
Key words: Mangifera indica L., polyembryony, microsatellites.
INTRODUCTION
Mango (Mangifera indica L.) is the sixth most important fruit tree in terms of production and the second most important tropical crop in terms of economic importance worldwide, surpassed only by bananas (FAO, 2016). Mexico is the fourth largest producer (SIAP, 2018) and the leading exporter around the world (SAGARPA, 2017). New mango cultivars are required to maintain production, ensure competitiveness, and avoid problems associated with pests and diseases (Khan et al., 2015).
Most mango cultivars are bred based on a selection of plants obtained by open and controlled pollination (Ruiz, 2020). In Mexico, the predominant type of cultivar is Ataulfo. This cultivar is polyembryonic, that is, a zygotic embryo and several nucellar embryos may be present in the seed to maintain the genetic constitution of the mother plant (Johri et al., 1992; Hand and Koltunow, 2014; Wang et al., 2017). Nucellar embryony is favorable for propagation by grafting since it allows the production of clonal plants and helps to maintain uniformity in plantations (Ruiz et al., 2020). However, it is undesirable for breeding programs because differentiating zygotic seedlings from nucellar seedlings is difficult at early stages; in addition, the development of nucellar embryos can inhibit zygotic embryos and cause their abortion (Naumova and Vielle-Calzada, 2001; Batygina and Vinogradova, 2007; Hand and Koltunow, 2014). Using random amplified polymorphic DNA (RAPD) markers, Martínez et al. (2012), identified two or three zygotic embryos in 14.3% of seeds, although the evaluated seeds were few and from only one population.
Given that microsatellite markers are specific, highly polymorphic, and codominant, they are more efficient than RAPD markers for studies involving diversity or intraspecific relationships (Duval et al., 2005; Azmat et al., 2016); thus, the purpose of the present study was to identify zygotic seedlings produced by the largest embryos in seeds from Oaxaca, Guerrero, Nayarit, and Sinaloa (México). The analysis used simple sequence repeat (SSR) microsatellites. Polyembryony was assumed to be significantly affected by the environment (Hand and Koltunow, 2014) and to have an influence on the survival of the zygotic embryo. In 2004, Andrade et al. (2004) carried out a study to determine the zygotic or nucellar origin in Citrus volkameriana using RAPD markers, obtaining satisfactory results. In the same way, in the case of the Mexican lemon (Citrus aurantifolia), RAPD markers have been used successfully, highlighting the use of the OPH15 decamer (Mondal and Saha, 2014); without there being reports in which other types of molecular markers, such as SSRs, have been used.
MATERIALS AND METHODS
Vegetative material
Ataulfo mangoes were harvested from the states of Guerrero and Oaxaca in the month of May 2017 and from cultivars in Nayarit and Sinaloa in July of the same year. One hundred fruits were harvested randomly per tree and harvest site when physiologically mature. Fruits were transferred to the laboratory (Genetic Transformation and Molecular Genetics, Campus Montecillo, Colegio de Postgraduados, Texcoco, Mexico) for in vitro culture and data collection.
In vitro culture
Under aseptic conditions, seed-bearing endocarps were disinfested in a sodium hypochlorite solution (Cloralex®) at 50% v/v under a laminar flow hood for 5 min and placed on sterile paper towels (Sanitas®) to remove excess moisture. Subsequently, the endocarp was removed, the seeds were numbered and then immersed in 80% ethanol for 1 min and 10 s and in 5% sodium hypochlorite solution for 3 min. Finally, the seed coat was removed, and embryos were seeded in culture medium (20 g·L-1 sucrose, 9 g·L-1 agar-Phytagel, 1 g·L-1 activated charcoal, 1 mg·L-1 thiamine, and 50 mg·L-1 myo-inositol; pH: 5.7) and stored in a dark incubation room at 24°C (Figure 1).
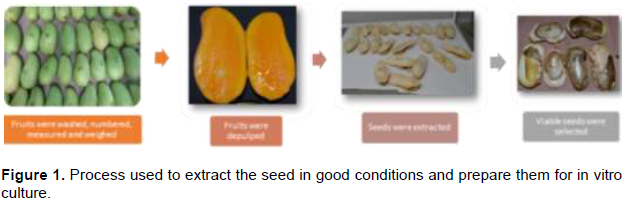
After two weeks, when embryos began to separate naturally due to imbibition, they were removed from the culture medium and classified by size, assigning the value of 1 to the largest embryo (Figure 2). Embryos of sizes 2, 3, and 4 were subcultured in culture medium (1 mg·L-1 benzyladenine, 1 mg·L-1 indoleacetic acid, 4 mL·L-1 ammonium nitrate (1 M), 20 mL·L-1 calcium nitrate (1 M), 50 mg·L-1 myo-inositol, 1 mg·L-1 thiamin, 20 g·L-1 sucrose, 9 g·L-1 PhitagelTM, 1 g·L-1 activated carbon; pH: 5.7) and stored in the incubation room at 24°C under a 16-h photoperiod and a light intensity of 30 μmol m-2s-1. The larger embryos were submerged in fungicide (Benomilo 1 g·L) for 5 min and transplanted in boxes with agrolite to be transferred to the "Vivero Cazones”, a plant nursery located in Cazones, Veracruz, Mexico. When seedlings originating from small embryos (sizes 2 to 4) reached between 5 and 7 cm in length, they were transplanted using the procedure described for the larger embryo. All embryos were placed on a substrate composed of pine bark, tepezil, earth, and vermicompost (30:30:25:15 v/v/v/v). Each plant was identified by number of embryo (1-4), number of seed (all seeds were numbered), and population where the fruit was collected (Oaxaca, Guerrero, Nayarit, or Sinaloa).

Extraction of DNA
Young (reddish-violaceous) and mature leaves from 72 plants originating from the largest embryos (18 samples per state) were collected, placed in perforated glassine bags, and transported in an ice box to the experimental site. These samples were frozen at -80°C and placed in the lyophilizer (Labconco Freezone® freeze dryer) for 24 to 48 h, ground (Retcsh® MM 400 mixer mill) at 35 Hz for 3 min, and stored at -20°C.
DNA was extracted using as a reference the methods of Saghai-Maroof et al. (1984) for species with high content of secondary metabolites and Doyle and Doyle (1987) based on the use of CTAB. 15 mg of lyophilized tissue were placed in 1.6-mL Eppendorf tubes with 600 μL saline buffer (1 M Tris-HCl, 2 M NaCl, 0.5 M EDTA pH 8, 0.05% serum albumin). The mixture was vortexed and allowed to stand for 15 min at room temperature and centrifuged at 5000 RCF for 4 min. The supernatant was decanted and 600 μL of 2% CTAB buffer and 3 μL β-mercaptoethanol were added; the mixture was vortexed until completely homogenized and incubated at 55°C for 1 h. 400 μL of chloroform: isoamyl alcohol (24:1) was added, and the mixture was vortexed and centrifuged at 12000 RCF for 10 min. The aqueous phase was transferred to a new Eppendorf tube, where 600 μL of (cold) isopropanol was added, and the phase was homogenized by inversion and allowed to stand for 20 min at -20°C. The phase was centrifuged at 5000 RCF for 4 min, and the supernatant was discarded. The formed pellet was added 600 μL of 75% ethanol (cold) and centrifuged at 5000 RCF for 4 min; the supernatant was decanted, and the tube was left at room temperature until the ethanol was evaporated entirely. Finally, the pellet was resuspended in 100 μL of sterile distilled water, added 5 μL of RNase (10 mM/mL), and incubated at 35°C for 30 min and then at 65°C for 5 min to inactivate the RNase.
DNA quantity and quality were evaluated using a NanoDrop® 1000 spectrophotometer (Thermo Scientific).
Microsatellite markers
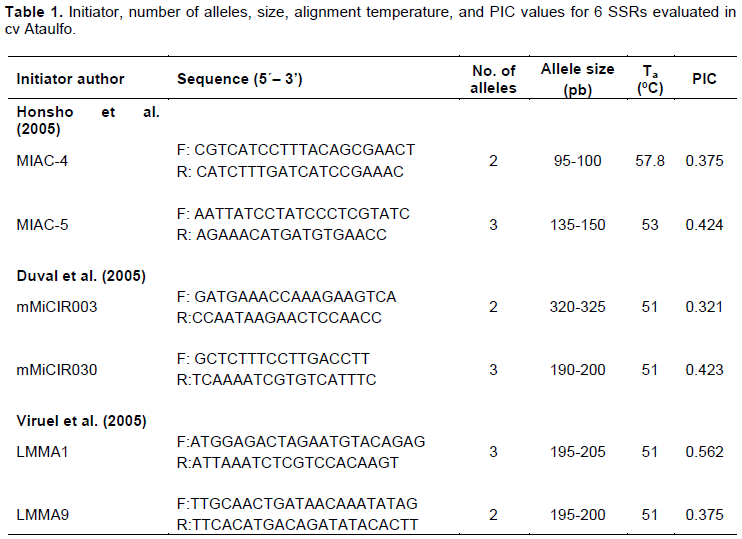
The present study focused on six microsatellite loci: MIAC-4, MIAC-5, mMiCIR003, mMiCIR030, LMMA1, and LMMA9, proposed by Arellano-Durán (2017). The PCR mixture (20 μL) contained 50 ng of DNA, 1X PCR Buffer (-Mg), 1.5 mM MgCl2, 2.5 mM dNTPs, 0.5 U DNA polymerase (Taq DNA Polymerase Invitrogen®), and 0.2 μM of each primer. The PCR reaction was performed in a MaxyGene® II Thermal Cycler (Axygen), and it consisted of an initial denaturation phase at 94°C for 4 min, followed by 30 cycles at 94°C for 45 s, at the defined alignment temperature for each initiator for 1 min (Table 1), at 72°C for 1 min, and a cycle at 72°C for 8 min. The PCR product was mixed with 5 μL loading buffer (50 mM Tris, 5 mM EDTA pH 8, 25% sucrose; 0.2% w/v bromophenol blue) and separated by vertical electrophoresis using an MGV-216-33 Dual Triple Wide Mini-Vertical electrophoresis system (CBS Scientific) and two-phase (running acrylamide and stacking acrylamide) polyacrylamide gel; running buffer was TG 1X. Bands were visualized by staining with silver nitrate (0.2%) (CIMMYT, 2006).
Statistical analysis
Nucellar plants were identified when compared with the SSR of the female parent; the presence of a band was given the value of '1' and the absence of a band was given a value of '0'; the studied genotypes were considered homozygous or heterozygous when one or two fragments were present per locus, respectively (Callen et al., 1993). The different amplifications to the female parent were recorded as polymorphisms. In order to determine the ability of each marker to differentiate the origin of the plants (nucellar or zygotic), the polymorphism information content (PIC) was calculated according to Novelli et al. (2006) , using CERVUS 3.0.7 software (Kalinowski et al., 2007). The genetic similarity index (GSI) between individuals of the same population was determined using the formula recommended by Mannen et al. (1993):
ISG=(2p+q)I2N,
where p is the total number of bands defined as S = 1 and q is the total number of bands defined as S = 0.5. UPGMA cluster analysis and Nei’s (1978) similarity coefficient were used for calculating genetic relationships between populations; all these analyses were carried out using the NTSYS v. 2.2 Statistical Package (Rohlf, 2000).
RESULTS
The number of alleles found with 6 primers varied from two to three and their size from 95 to 325 bp. The alignment temperatures used in 5 markers corresponded to report by Arellano-Durán (2017), except for MIAC-5 (Table 1).
The PIC calculated for 6 SSR markers ranged from 0.321 to 0.562, with an average PIC of 0.413 (Table 1). According to Botstein et al. (1980) criteria, five initiators are "moderately informative" (PIC 0.25-0.5), and only one marker (LMMA1) was "highly informative" (PIC> 0.5).
Eleven plants out of 72 presented polymorphisms with one to five primers (Table 2), thus, 15.27% of seedlings were of zygotic origin. The seedling from the largest embryo (1) in the seed, identified with number 94 for the in vitro culture (Figure 2) and appearing as ‘02’ in the electrophoresis results (Figure 3), was the only one that showed polymorphism in 5 of 6 SSR, which indicate that it was homozygous.
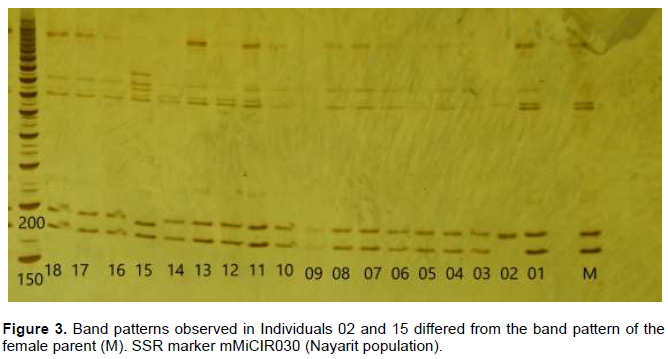
The GSI, proposed by Mannen et al. (1993), states that if values are greater than 95%, plants are considered to be genetically identical to their female parent. With this premise, 15.27% of seedlings were different, with GSI from 0.50 to 0.90 (Table 3). None of the genotypes was found to be identical to the mother plant: in Oaxaca, Guerrero, and Sinaloa populations, 3 out of 18 seedlings (16.6%) were different and 2 of 18 (11%) seedlings were different in the Nayarit population.
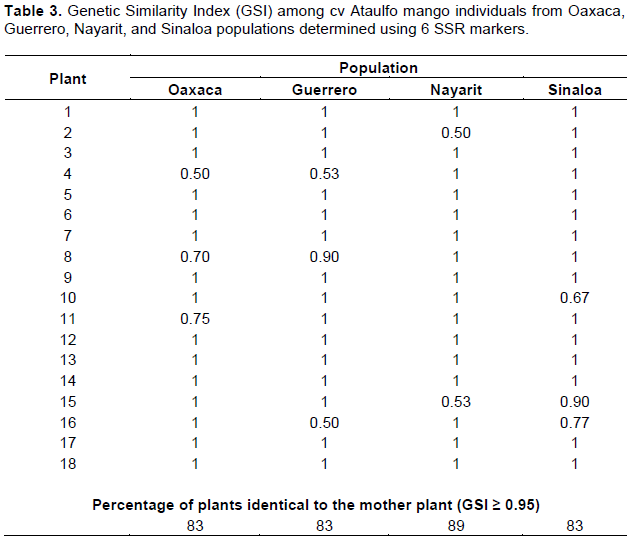
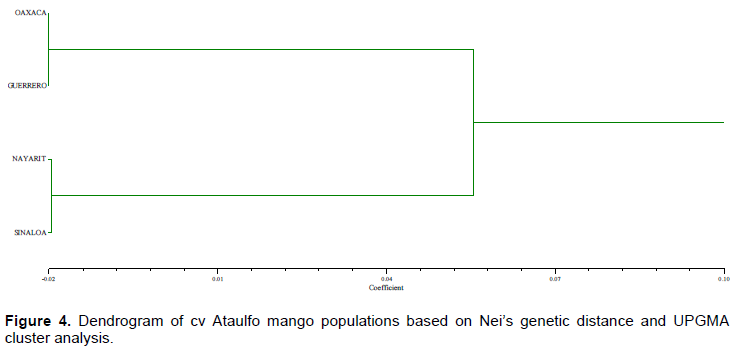
The genetic similarity among the four populations evaluated is high (93.8%). The populations from Guerrero-Nayarit are the most genetically distant (0.062), followed by Sinaloa-Guerrero (0.057), Oaxaca-Nayarit (0.054) and Sinaloa-Oaxaca (0.049). The distance between Oaxaca-Guerrero and Sinaloa-Nayarit was the lowest (-0.020).
The UPGMA cluster analysis separated the populations into two groups: Oaxaca-Guerrero and Nayarit-Sinaloa (Figure 4).
DISCUSSION
Allele sizes found in the present study were comparable to data reported in the literature for the same SSRs in different mango cultivars of different provenances (India, Pakistan, Florida, Filipinas, Thailand, Mexico), both monoembryonic and polyembryonic (Duval et al., 2005; Honsho et al., 2005; Viruel et al., 2005; Azmat et al., 2016); however, the number of alleles was slightly lower, possibly due to the low genetic diversity resulting from evaluating only one cultivar, and considering that the largest embryos were, theoretically, of nucellar origin; genetic variability was further reduced.
Concerning polymorphic information content (PIC), the results differ from those reported by Azmat et al. (2016), who classified MIAC-5 and mMiCIR030 as "highly informative"; MIAC-4 as "moderately informative", and LMMA1, LMMA9 and mMiCIR003 as "not informative". The difference in PIC values may be due to the low number of alleles measured in the present study, since there is a significant positive correlation (0.66) between the number of alleles amplified by a particular marker and its PIC value (Azmat et al., 2016).
Each microsatellite is a locus; therefore, different coding and non-coding regions of the genome are analyzed when using several SSRs. These markers are highly specific, and their length polymorphism ranges from 1 to 2 bp. Consequently, plants whose band patterns were different from the female parent’s band pattern, at least in a microsatellite were zygotic. However, Cordeiro et al. (2006) and Martínez et al. (2012) have classified them as nucellar using RAPD markers. The fact that some seedlings are polymorphic in fewer microsatellites may be due to the greater kinship between the female and male parent in comparison with seedlings whose parents showed polymorphism with more microsatellites. The SSR markers constitute a useful tool for the study of the genetic diversity of citrus fruits due to their efficacy and information (Mohamed et al., 2011). Its use for the detection of hybrids (Mondal et al., 2015) is more practical, effective and advantageous, because the studies that used RAPD and ISSR (Andrade et al., 2004; Golein et al., 2011) generally obtain a large number of non-informative monomorphic bands that can hinder the data analysis. RAPD and ISSR, being dominant markers, do not allow differentiation of heterozygous individuals, unlike SSR, which are codominant markers.
All the plants that showed different banding patterns with respect to the female parent were considered polymorphic; consequently, the percentage of zygotic plants using the polymorphism of the SSR and the GSI was the same.
There was no microsatellite in which all the seedlings showed polymorphism, although three markers (MIAC-4, LMMA9, and MIAC-5 or mMiCIR030) were sufficient to identify the zygotic or nucellar origin of the 72 plants analyzed in the present study. Similarly, Viruel et al. (2005) point out that three selected microsatellites allow for the unambiguous identification of mango genotypes of different geographical origin and seed type.
It is important to mention that none of the different individuals in the population showed uniformity in all their progeny. Although the fruits could be distinguished morphologically, the generic similarity among the four populations was high since all of them were from cv Ataulfo. The populations from Oaxaca and Guerrero were very close both genetically and geographically, as well as the populations from Nayarit and Sinaloa; however, these two groups presented a certain degree of variability, which can be attributed to the selection process and the different environments where the populations were sampled. The first population could have been very different: it is believed that only one of the nine mango trees that were initially planted in Mr. Ataulfo Morales’s orchard had the best characteristics, but the material of the rest of the trees was also propagated (personal communication). Nevertheless, only the clone IMC-M2 Ataulfo spread extensively (Infante et al., 2011). In fact, Salvador et al. (2008) reported 4.3% of genetic variability among individuals in the father orchard of the cv Ataulfo, which was associated with cross-pollination.
The genetic variability found by the present study can also be due to the genetic origin of the propagated plant; two plants usually emerge from the planted seed, and nurserymen tend to choose the larger one assuming that the cutting is a "clone" of the progenitor. In the cv Rosinha, Cordeiro et al. (2006) found that 90% of the more vigorous plants were of zygotic origin; eighteen seeds out of 30 in the cv Ubá contained zygotic embryos, and the zygotic seedling was the more vigorous in six of the 18 seeds (20%) (Rocha et al., 2014). In the present study, 15.27% of seedlings, originated by the largest embryo in the seed, were classified as zygotic. Although the percentage of zygote seedlings was not as high as in the cv Rosinha, not all the highly vigorous cv Ataulfo seedlings or the larger embryos were clones of the mother plant.
Results found by Cordeiro et al. (2006), Martínez et al. (2012), and Rocha et al. (2014) show that the zygotic embryo does not always degenerate and can give rise to the most vigorous plant of the seed.
In mango, more than two zygotic seedlings have been identified per seed (Martínez et al., 2012; Rocha et al., 2014); consequently, identifying the genetic origin of embryos 2 to 4 using SSR markers is advisable as an addition to existing information, to verify previously obtained data, and to identify a possible alternative apomictic route in addition to the nucellar.
CONCLUSION
The 15% of seedlings originated by the largest embryo in the seed are not clones of the mother plant. Markers SSR, MIAC-4, MIAC-5, mMiCIR030, mMiCIR003, LMMA9, and LMMA1 are recommended to identify the genetic origin of mango seedlings. Three SSRs were sufficient to identify the zygotic or nucellar origin of the 72 plants analyzed in this work.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
The authors express their profound gratitude to the National Council of Science and Technology (CONACYT) for financing the project and to Colegio de Postgraduados for the facilities and support provided during the development of the study.
REFERENCES
|
Andrade RM, Villegas MA, Gutiérrez EMA, Carrillo CG, García VA (2004). Polyembryony and RAPD markers for identification of zygotic and nucellar seedlings in Citrus. Agrociencia 39(4):371-383. |
|
|
Arellano-Durán N (2017). Poliembrionía en mango Manila y Ataulfo: análisis molecular y desarrollo de embriones cigóticos y nucelares. Tesis de Doctorado. Colegio de Postgraduados, México 100 p. |
|
|
Azmat MA, Khan AA, Iqrar AK, Rajwana IA, Cheema HMN, Sattar AK (2016). Morphological characterization and SSR based DNA fingerprinting of elite commercial mango cultivars. Pakistan Journal of Agricultural Sciences 53:321-330. |
|
|
Batygina TB, Vinogradova GY (2007). Phenomenon of polyembriony. Genetic heterogeneity of seeds. Russian Journal of Developmental Biology 38(3):126-151. |
|
|
Botstein D, Raymond LW, Skolnick M, Davis RW (1980). Construction of a genetic linkage map in man using restriction fragment length polymorphisms. American Journal Human Genetic 32(3):314-331. |
|
|
Callen DF, Thompson AD, Shen Y, Phillips HA, Richards RI, Mulley JC, Sutherland GR (1993). Incidence and origin of 'null' alleles in the (AC) n microsatellite markers. American Journal Human Genetic 52(5):922-927. |
|
|
CIMMYT (2006). The applied biotechnology center manual of laboratory protocols. Third Edition. CIMMYT, D.F., México. |
|
|
Cordeiro MCR, Pinto ACQ, Ramos VHV, Faleiro FG, Fraga LMS (2006). Identificação da origem genética de plântulas em sementes poliembriônicas de mangueira (Mangifera indica L.) cv. Rosinha por meio de marcadores RAPD. Revista Brasileira de Fruticultura 28(3):454-457. |
|
|
Duval MF, Bunel J, Sitbon C, Risterucci AM (2005). Development of microsatellite markers for mango (Mangifera indica L). Molecular Ecology Notes 5(4):824-826. |
|
|
Food and Agriculture Organization (FAO) (2016). FAOSTAT, Dirección de Estadística. Consultado en línea: |
|
|
Golein B, Fifaei R, Ghasemi M (2011). Identification of zygotic and nucellar seedlings in citrus interspecific crosses by inter simple sequence repeats (ISSR) markers. African Journal of Biotechnology 10(82):18965-18970. |
|
|
Hand ML, Koltunow AMG (2014). The Genetic Control of Apomixis: Asexual Seed Formation. Genetics 197(2):441-450. |
|
|
Honsho C, Nishiyama K, Eiadthong W, Yonemori K (2005). Isolation and characterization of new microsatellite markers in mango (Mangifera indica). Molecular Ecology Notes 5(1):152-154. |
|
|
Infante F, Quilantán J, Rocha F, Esquinca H, Castillo A, Núñez GI, Palacio V (2011). Mango Ataulfo: Orgullo chiapaneco. CONABIO. Biodiversitas 96:1-5. |
|
|
Johri BM, Ambegaokar KB, Srivastava PS (1992). Comparative embryology of angiosperms. Springer-Verlag. Berlín, Heidelberg, New York, London, Paris, Tokyo 2:463-466. |
|
|
Kalinowski ST, Taper ML, Marshall TC (2007). Revisando cómo el programa de computadora CERVUS se adapta al error de genotipificación aumenta el éxito en la asignación de paternidad. Ecología Molecular 16:1099-1106. |
|
|
Khan AS, Ali S, Khan IA (2015). Morphological and molecular characterization and evaluation of mango germplasm: An overview. Scientia Horticulturae 194:353-366. |
|
|
Mannen H, Tsuji S, Mukai F, Goto N, Ohtagaki S (1993). Genetic similarity using DNA fingerprinting in cattle to determine relationship coefficient. Journal of Heredity 84(3):166-169. |
|
|
Martínez OEC, Andrade RM, Rocandio RM, Villegas MA (2012). Identification of zygotic and nucellar seedlings in polyembryonic mango cultivars. Pesquisa Agropecuaria Brasileira 47(11):1629-1636. |
|
|
Mohamed HA, Manosh KB, Zongwen Z, Wen-Wu G (2011). Exploitation of SSR, SRAP and CAPS-SNP markers for genetic diversity of Citrus germplasm collection. Scientia Horticulturae 128(3):220-227, ISSN 0304-4238, |
|
|
Mondal B, Pramanick S, Saha R, Karmakar M (2015). Application of simple sequence repeat markers for demarcation of Citrus reticulata nucellar and hybrid seedlings. International Journal of Biosciences 6(2):128-133. |
|
|
Mondal B, Saha R (2014). Identification of zygotic and nucellar seedling of Citrus aurantifolia and Citrus reticulata using RAPD. International Journal of Advanced Biotechnology 15(1):25-30. |
|
|
Naumova TN, Vielle CJP (2001). Ultrastructural analysis of apomictic development. In: Savidan, Y., J.G. Carman, and T. Dresselhaus (eds). The Flowering of Apomixis: From Mechanisms to Genetic Engineering. México, D.F., CIMMYT, IRD, European Commission DG VI (FAIR) pp. 44-66. |
|
|
Nei M (1978). Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 89(3):583-590 |
|
|
Rocha A, Salomão TMF, Siqueira DL, Cruz CD, Salomão LCC (2014). Identification of Ubá mango tree zygotic and nucellar seedlings using ISSR markers. Revista Ceres 61(5):597-604. |
|
|
Rohlf FJ (2000). NTSYS-pc: Numerical Taxonomy and Multivariate Analysis System version 2.2 Excenter Sofware, New York, USA. |
|
|
Ruiz M, Oustric J, Santini J, Morillon R (2020). Synthetic Polyploidy in Grafted Crops. Frontiers in Plant Science 11:540894. |
|
|
SAGARPA (2017). MANGO Mexicano, Planeación Agrícola Nacional 2017-2030. Ciudad de México, México. Consultado en línea: |
|
|
Salvador FM, Torres DR, Ovando MI, Vázquez OJA, Adriano AML (2008). Análisis de variabilidad genética del huerto padre del mango (Mangifera indica Linn) variedad Ataúlfo. Quehacer CIENTÍFICO en Chiapas, Enero-Junio 05:29-34. |
|
|
SIAP (2018). Atlas Agroalimentario 2012-2018. SAGARPA, México. Consultado en línea: |
|
|
Viruel MA, Escribano P, Barbieri M, Ferri M, Hormaza JI (2005). Fingerprinting, embryo type and geographic differentiation in mango (Mangifera indica L., Anacardiaceae) with microsatellites. Molecular Breeding 15(4):383-393. |
|
|
Wang X, Xu Y, Zhang S, Cao L, Huang Y, Cheng J, Wu G, Tian S, Chen C, Liu Y, Yu H (2017). Genomic analyses of primitive, wild and cultivated citrus provide insights into asexual reproduction. Nature Genetics 49(5):765-772 |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


