Full Length Research Paper

ABSTRACT
Noni (Morinda citrifolia) is a fruit species which is well adapted to different soil and climate conditions, and may be a good option for farmers in arid and semi-arid regions. To evaluate the tolerance of noni plant in the initial phase to salinity and the accumulation of inorganic solutes in its organs, an experiment was carried out in a greenhouse under hydroponic system. The experimental design was completely randomized, with five levels of NaCl (0, 25, 50, 75 or 100 mM) in the nutrient solution, and four replicates. Plant height, stem diameter, number of leaves, dry matter of leaves, stems and roots, leaf area, leaf succulence, sclerophylly index and the contents of Na+, Cl- and K+ in different plant organs were evaluated after 40 days of stress. Salinity reduced the growth of all plant organs in the initial phase, in contrast, leaf succulence and sclerophylly index increased by 32% at the highest saline level, compared with control plants. The contents of Na+ and Cl- in leaves, stems and roots increased, while K+ contents decreased with the increment in salinity. The highest K+ contents were observed in the roots and the highest Na+ and Cl- contents were observed in the leaves. Salinity induced disturbances in the ionic homeostasis of noni plants, which can explain, at least in part, the salt-induced growth reduction.
Key words: Salinity, sodium chloride, toxic ion.
INTRODUCTION
The cultivation of salt-tolerant species has called the attention of researchers and farmers in order to achieve an economically viable production in salt-affected areas. The saline soils are located mainly in arid and semi-arid regions. In these areas, the soil and climatic conditions and inadequate irrigation management favor the salt accumulation in soil and water. In Brazil, the semi-arid region occupies an area of 980,133.079 km², located mainly in the Northeast of the country, with about 53 million inhabitants (IBGE, 2011; Santos et al., 2014). Therefore, the introduction of salt-tolerant plants is essential to maintain the crop production in these areas. The noni plant acclimates to the most diverse environmental conditions, including high levels of salts in the soil (Nelson and Elevitch, 2006). Under favorable conditions, the plant produces fruits nine to twelve months after planting (Chan-Blanco et al., 2006), which turns the noni (Morinda citrifolia) as an interesting crop to be introduced in salt-affected areas.
Noni is an exotic plant, belonging to the Rubiaceae family, and a fruit crop with medicinal and nutritional value that has been cultivated in several states of Brazil (Correia, 2010). This plant has been used for thousands of years by the Polynesians in the combat of various diseases. Noni seeds, peel and pulp have significant amounts of carbohydrates, proteins, vitamin C, caro-tenoids and phenolic compounds, which have antioxidant activity in vitro (Costa et al., 2013).
Salinity affects plant development inducing osmotic and ionic stress (Munns and Tester, 2008). However, the plant can develop mechanisms to survive to these conditions. Na+ and Cl- are the main ions found in saline soils. These salts, when absorbed and accumulated by plants, can contribute to osmotic adjustment or be toxic; however, the amounts of salts and the capacity of the plant to cope with salt stress will determine the crop production. Therefore, understanding the mechanisms of plant tolerance to salinity is fundamental for the introduction of this crop in salt-affected areas. Thus, the study aimed to evaluate the effect of salinity on the initial growth and accumulation of inorganic solutes in the different parts of noni plants in order to know its tolerance level to salt stress.
MATERIALS AND METHODS
The experiment was carried out in a greenhouse of the Center of Soil and Water Engineering, at the Federal University of Recôncavo of Bahia, in Cruz das Almas-BA, Brazil (12º40'19"S, 39º 6'23"W). The mean values of temperature, relative air humidity and photosynthetically active radiation (at noon) during the experimental period were 25° C, 81% and 1200 µmol m-2 s-1, respectively.
Mature noni fruits were selected for collection of seeds, prioritizing the characteristics of plant sanitary conditions and size. After the removal of seeds, dormancy breaking was performed by immersing the seeds for 48 h in water (Leite et al., 2012). After 50 days of sowing, the seedlings with two cotyledonary leaves were transferred to 500-mL plastic cups, containing a mixture of Red-Yellow Latosol and humus (1:1), irrigated daily with well water and weekly with 50 mL of nutrient solution (Furlani, 1998).
Noni seedlings three months old with four pairs of leaves were transferred to containers with 12 L of Furlani nutrient solution, in a floating hydroponic system, where they remained for four days for acclimation. After this period, the seedlings were subjected to five treatments, which consisted in the addition of 0, 25, 50, 75 or 100 mM of NaCl in the nutrient solution. The NaCl was gradually added (25 mM day-1), in order to avoid the osmotic shock. The volume of the solutions was completed daily with water and the renewal was performed weekly. The pH was maintained at 6.0 ± 0.2 by adding NaOH or HCl. The system was maintained under intermittent aeration of 15 min every hour, using an air compressor coupled to a timer.
At the end of the experimental period (40 days), the plants from all treatments were carefully removed from the nutrient solution, the roots were washed with distilled water and the plants were divided into different organs for fresh mass (FM), dry mass (DM), and leaf area (LA) determinations. LA was measured with the WinDIAS image analysis system, model W-C110-PC (Delta-T Devices Ltd, Cambridge, UK). The leaf (LDM), stem (SDM), and root (RDM) dry masses were determined after drying of plant parts in an oven at 65°C, for 72 h. These data were used to calculate the shoot to root dry mass ratio (S/R). The values of leaf fresh mass (LFM), LDM and LA were used to calculate leaf succulence and sclerophylly index (SI), according to the equations proposed by Benincasa (2003):
Succulence = (LFM-LDM)/LA
SI = LDM (mg)/LA (cm2).
For determination of Na+, K+ and Cl- contents, the extracts were prepared as described by Jones (2001), with minor modifications. 100 mg of leaf, stem or root ground material were mixed with 10 mL of deionized water in test tubes. The tubes were maintained for 1 h at 80°C in water bath, with shaking every 15 min. After this period, the tubes were centrifuged at 5,000 × g for 15 min, at room temperature. The Na+ and K+ contents were determined by flame photometry (Faithfull, 2002) and the Cl- contents by spectro-photometry (Jones, 2001).
The experiment was conducted in a completely randomized design, with five salinity levels (0, 25, 50, 75 or 100 mM of NaCl) and four replicates. The data were subjected to analysis of variance and regression using SISVAR 4.6 statistical software (Ferreira, 2011). The models were selected based on significance level by F test and determination coefficient (R²).
RESULTS AND DISCUSSION
The increase of NaCl in the nutrient solution linearly decreased the plant height, stem diameter and number of leaves, with reductions of 0.225 cm, 0.041 mm and 0.255, respectively, per mM NaCl in solution (Figure 1). Thus, among biometric variables, the reduction in 100 mM NaCl was higher in the plant height (62%), number of leaves (62%) and lower in the stem diameter (35%), as compared to the control. Similar results were also reported by Souto et al. (2013), in noni plants grown in soil and irrigated with saline water. The reduction in plant height, stem diameter and number of leaves may affect biomass production. Studies on noni have demonstrated that salt-induced changes in growth variables directly affect crop yield (Souto et al., 2015, 2016).
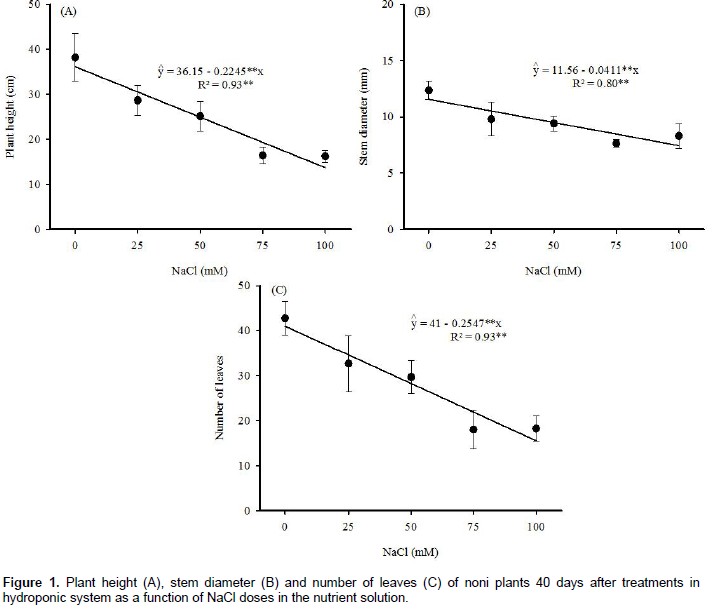
Figure 2 shows that LDM, SDM and TDM presented decreasing linear response, with reductions of 0.146, 0.074 and 0.329 g per mM NaCl, hence reductions of 72, 49 and 68% at the highest NaCl level, respectively. However, for the S/R, succulence and SI, there were increases of 0.0319, 0.0823 mg H2O cm-2 and 0.0202 mg of DM cm-2 for each increment in salinity, that is, increases of 117, 32 and 32% at 100 mM of NaCl, in relation to control treatment.
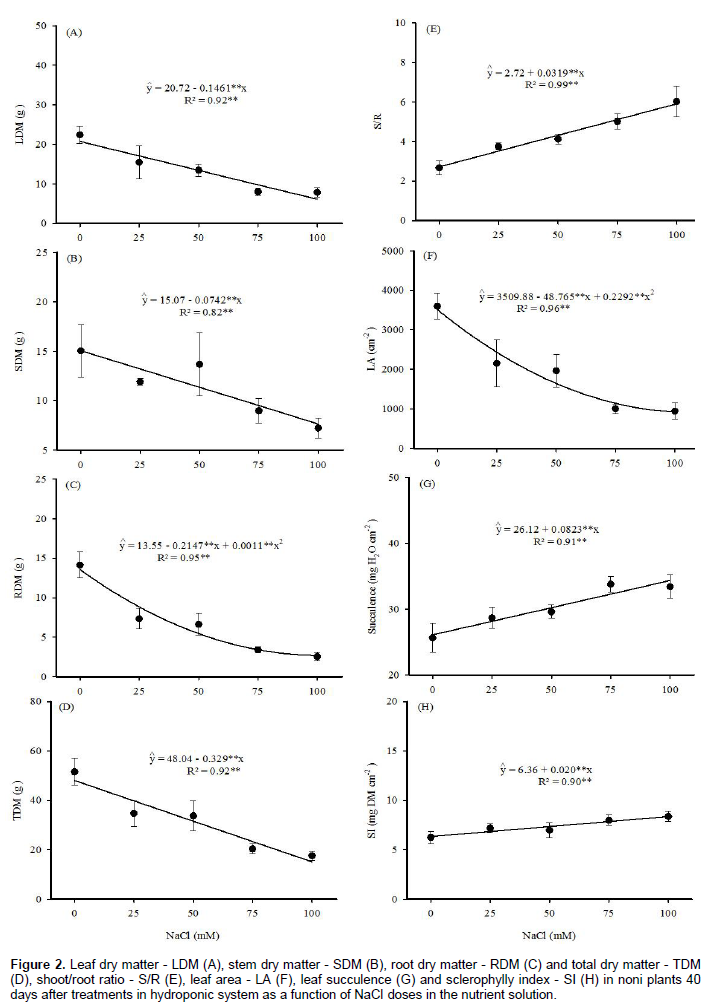
RDM and LA followed a quadratic polynomial model, with more pronounced reductions (35 and 31%) at 25 mM NaCl in comparison with control, and reductions of 77 and 74%, respectively in 100 mM NaCl. The reduction in noni LA may decrease photosynthesis, absorption of water and nutrients, and consequently, affect growth and biomass production (Souto et al., 2013).
Comparing the different plant organs, the roots showed the greatest reduction in dry mass induced by the saline stress suggesting that the root is the most sensitive organ to the deleterious effects of salinity in noni plants. Rodrigues et al. (2014) observed a reduction of 52% in Ricinus communis roots, which was the most affected organ at 150 mM NaCl in the nutrient solution, compared with the control plants. Although Abreu et al. (2008) suggested that roots have greater capacity for osmotic adjustment and better protection against oxidative stress under saline stress conditions, this growth reduction may occur because the root is the first organ directly exposed to the salt stress (Azevedo Neto et al., 2006).
The higher sensitivity of roots to salt stress, in comparison to the shoots, led to a linear increase in the S/R, which varied from 2.72 in the control to 5.91 in the treatment of 100 mM NaCl. According to Marana et al. (2008), in coffee seedlings, a plant of the same family of noni, values between 4.7 and 7.0 are considered as satisfactory for plant development.
Plants, when subjected to abiotic stress, including salinity, can develop mechanisms to survive to environ-mental conditions, such as changes in leaf succulence and SI. The increment in succulence observed in the present study indicates an increase in the amount of H2O per unit of LA, which may be important for saving water through transpiration, water storage, and dilution of the toxic ions Na+ and Cl-. In turn, increased SI indicates that the leaves are thickened, with more strata of photosynthetic tissues, which may be important to reduce the deleterious effects of salinity on plant growth (Oliveira et al., 2012).
Reductions in growth variables of noni irrigated with saline water have recently been reported in the literature (Souto et al., 2013; Souza et al., 2014; Souto et al., 2015). These authors attributed the results to the increase in soil salinity, which affects water absorption, and to the accumulation of toxic ions and reduction of photosynthetic pigments. The accumulation of the toxic ions Na+ and Cl- in the different plant organs, the nutritional imbalance and/or the water deficit induced by salinity can affect the plant growth (Munns et al., 2006; Munns and Tester, 2008; Bosco et al., 2009).
The Na+ and Cl- contents in leaves, stems and roots of noni plants, as a function of NaCl concentrations in the nutrient solution are shown in Figure 3. It can be observed that the Na+ and Cl- contents increased quadratically, except for the Cl- contents in the roots, which followed a linear model. In relation to the control, the contents of Na+ and Cl- in 100 mM of NaCl increased, respectively, 15 and 6 times in the leaves, 31 and 4 times in the stem and 6 and 5 times in the roots. It can also be observed in Figure 3 that, at 100 mM of NaCl, the estimated values for Na+ and Cl- were 1.81 and 1.17 mmol g-1 DM in the leaves, 0.80 and 0.75 mmol g-1 DM in roots, respectively.
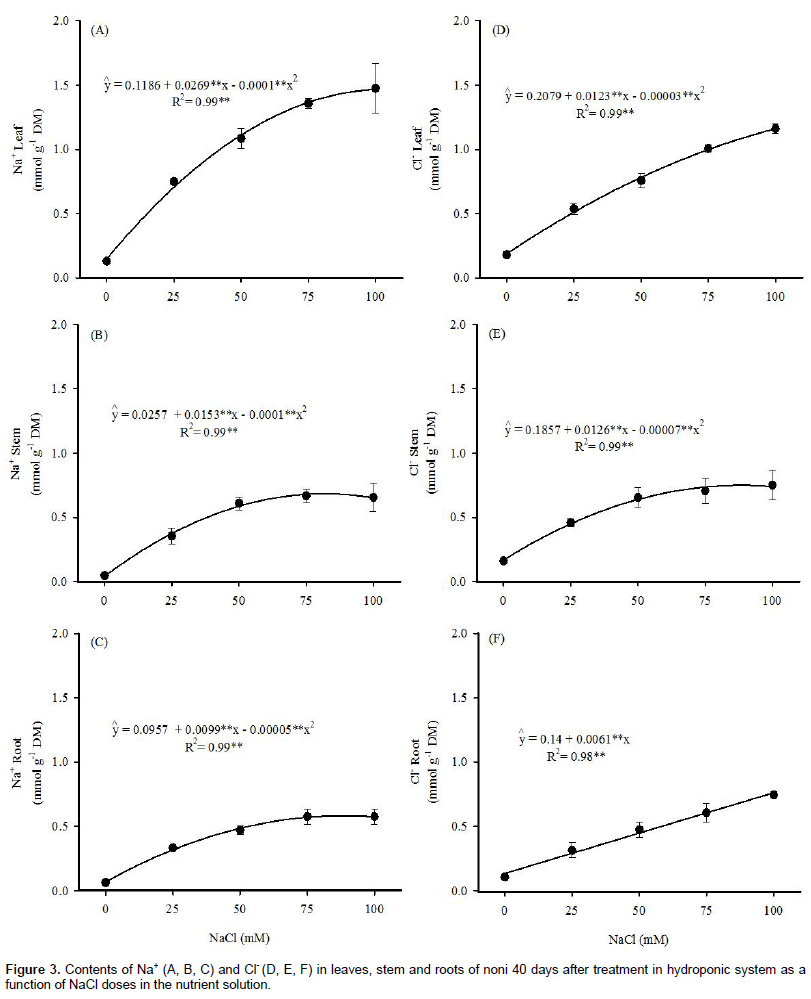
The high content of Na+ in the leaf tissue can affect physiological and biochemical processes, such as stomatal opening, respiration and synthesis of proteins, consequently affecting photosynthesis, growth and yield of the crops (Apse and Blumwald, 2007; Tavakkoli et al., 2010; Furtado et al., 2013). Thus, among the characteristics of salt-tolerant plants, the capacity to restrict the transport and accumulation of toxic ions in the leaves has been considered as the most important (Munns and Tester, 2008).
Chloride in high concentrations can reduce photo-synthetic capacity and quantum yield, due to chlorophyll degradation (Tavakkoli et al., 2010). On the other hand, Rodrigues et al. (2014) reported that the accumulation of Na+ and Cl- in leaves of R. communis played an important role in the osmotic adjustment.
Considering that the highest Na+ and Cl- accumulations were observed in the leaves, the results obtained in this study indicate that noni plants have no mechanisms of exclusion of these ions in the organs with smaller metabolic activity, such as stem and roots.
According to Taiz and Zeiger (2013), values above 100 mM of Na+ and Cl- become cytotoxic, causing protein denaturation and destabilization of membranes. Considering the data of LFM and LDM, the estimated concentrations of Na+ and Cl- in the leaf were, respectively, 188 and 134 mM at the lowest salinity level (25 mM NaCl). These values are similar to those reported by Azevedo Neto and Tabosa (2000) in the leaves of maize plants grown in nutrient solution with 100 mM NaCl. These authors considered that the ionic compartmentalization and cytoplasmic accumulation of compatible solutes was necessary for the reduction of ionic toxicity. Thus, the growth reduction observed in noni can, at least in part, be related to the toxic effects of Na+ and Cl- accumulation. However, since the NaCl levels were not lethal and did not cause early senescence of the leaves, the data of the present study also suggest that noni plant utilizes mechanism of Na+ and Cl- compartmentalization.
The K+ content and the K+/Na+ ratio in leaves, stems and roots of noni plants as a function of NaCl doses applied in the nutrient solution are shown in Figure 4. The K+ contents in the plant organs decreased with the increment of NaCl in the nutrient solution and fitted to an exponential regression model in the leaves, quadratic in the stem and linear in the roots, which represented reductions of 65, 59 and 41%, respectively, in 100 mM NaCl.
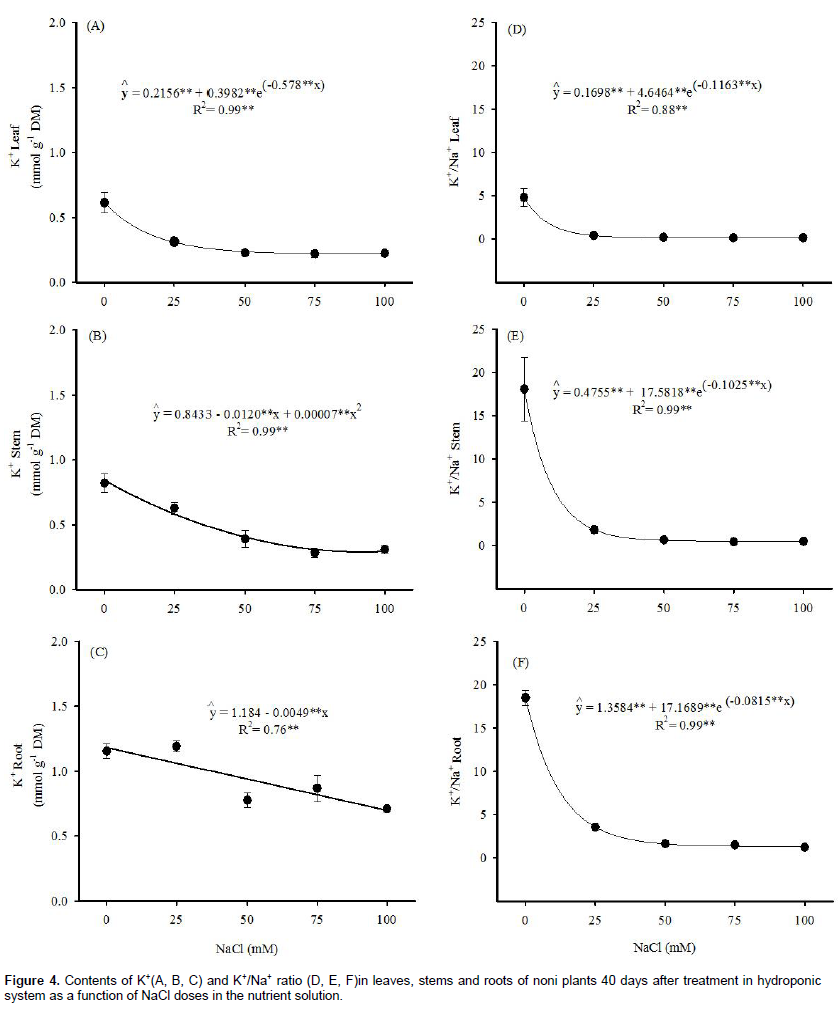
According to Bosco et al. (2009), high Na+ concen-trations can reduce K+ absorption by plants. Various factors can affect the absorption of this ion under saline conditions, especially the antagonism between K+ and Na+ for absorption sites in the plasmalemma and disorders in the membrane integrity (Apse and Blumwald, 2007). These results corroborate with those of Rodrigues et al. (2014), in castor bean subjected to increasing levels of NaCl in the nutrient solution, leading to a reduction of photosynthesis and dry mass yield and increase of S/R ratio.
The K+/Na+ ratio has been considered as an important variable in studies on the plant nutrition under salt stress (Bosco et al., 2009). In the present study, this variable decreased exponentially in all plant organs. Thus, the K+/Na+ ratio in leaf, stem and root decreased by 96, 97 and 93%, respectively, at 100 mM NaCl. The abrupt reduction of the K+/Na+ ratio in noni plants was due to the increase in Na+ uptake in conjunction with the decrease in K+ uptake. Many studies have shown that, under saline conditions, several crops alter the K+/Na+ ratio, causing nutritional imbalance, due to the antagonism between these elements (Silva et al., 2008; Bosco et al., 2009). According to Greenway and Munns (1980), the K+/Na+ ratio in glycophytes must be higher than 1.0 for the maintenance of ionic homeostasis and an optimal metabolic efficiency. Based on the regression equation of K+/Na+ ratio in the leaves, the maximum dose would be equal to 15 mM NaCl for the maintenance of a ratio adequate for the metabolism in noni.
CONCLUSIONS
1. Salinity negatively affects biomass production in noni plant and the root is the main organ affected by the stress in the initial phase;
2. Salinity reduces leaf area and increases succulence and sclerophylly index as a mechanism of tolerance to salinity in noni plant;
3. Noni plant in the initial phase does not have mechanisms of restriction to the transport of Na+ and Cl- to the leaves, which show higher contents of these ions, while having the compartmentalization of Na+ and Cl- as a mechanism of tolerance to salinity;
4. The increment in salinity until the dose of 100 mM causes alterations in the ionic homeostasis of noni plants, which can, at least partially, explain the salt-induced growth reduction.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
The author would like to thank the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) and the Universidade Federal do Recôncavo da Bahia (UFRB) for the financial support.
REFERENCES
|
Abreu CEB, Prisco JT, Nogueira ARC, Bezerra MA, Lacerda, CF, Gomes-Filho, E (2008). Physiological and biochemical changes occurring in dwarf-cashew seedlings subjected to salt stress. Braz. J. Plant Physiol. 20(2):105-118. |
|
|
Apse MP, Blumwald E (2007). Na+ transport in plants. FEBS Letters. 581(1):2247-2254. |
|
|
Azevedo Neto AD, Prisco JT, Enéas-Filho J, Abreu CEB, Gomes-Filho E (2006). Effect of salt stress on antioxidative enzymes and lipid peroxidation in leaves and roots of salt-tolerant and salt-sensitive maize genotypes. Environ. Exp. Bot. 56(1):87-94. |
|
|
Azevedo Neto AD, Tabosa JN (2000). Salt stress in maize seedlings: Part II Distribution of cationic macronutrients and its relation with sodium. Rev. Bras. Eng. Agric. Ambient. 4(2):165-171. |
|
|
Benincasa MMP (2003). Análise de crescimento de plantas: noções básicas. 2.ed. Jaboticabal: Funep. Brazil 41 p. |
|
|
Bosco MRO, Oliveira AB, Hernandez FFF, Lacerda CL (2009). Influence of salt stress on the mineral composition of eggplant seedlings. Rev. Ciênc. Agron. 40(2):157-164. |
|
|
Chan-Blanco Y, Vaillant F, Perez AM, Reynes M, Brillouet J-M, Brat P (2006). The noni fruit (Morinda citrifolia L.): A review of agricultural research, nutritional and therapeutic properties. J. Food Compos. Anal. 19(1):645-654. |
|
|
Correia AAS (2010). Maceração enzimática da polpa de noni (Morinda citrifolia L.), Universidade Federal do Ceará, Fortaleza, Brazil 106 p. |
|
|
Costa AB, Oliveira AMC, Silva AMO, Macini-Filho J, Lima A (2013). Atividade antioxidante da polpa, casca e sementes do noni (Morinda citrifolia Linn). Rev. Bras. Frutic. 35(2):345-354. |
|
|
Faithfull NT (2002). Methods in agricultural chemical analysis: A practical handbook. Wallingford: CABI Publishing 266 p. |
|
|
Ferreira DF (2011). Sisvar: A computer statistical analysis system. Ciênc. Agrotec. 35(6):1039-1042. |
|
|
Furlani PR (1998). Instrução para o cultivo de hortaliça de folha pela técnica de hidroponia - NFT. Campinas: Instituto Agronômico, (Boletim Técnico, 168). |
|
|
Furtado GF, Soares LAA, Sousa JRM, Andrade EMG, Guerra HOC (2013). Physiological changes in cowpea under saline water and nitrogen fertilization. Rev. Verde Agroecologia Desenvol. Sustent. 8(3):175-181. |
|
|
Greenway H, Munns R (1989). Mechanisms of salt tolerance in non-halophytes. Annu. Rev. Plant. Physiol. 31(1):149-190. |
|
|
Instituto Brasileiro de Geografia e Estatística – IBGE (2011). Censo Demográfico. 2010. Rio de Janeiro: IBGE. |
|
|
Jones JB (2001). Laboratory guide for conducting soil tests and plant analysis. Boca Raton: CRC. Press pp. 205-206. |
|
|
Leite GA, Cunha PSCF, Mendonça LFM, Medeiroa PVQ, Mendonça V (2012). Superação de dormência de sementes de noni. Rev. Verde Agroecologia e Desenvolv. Sustent. 7(4):120-128. |
|
|
Marana JP, Miglioranza E, Fonseca EP, Kainuma HK (2008). Seedling quality in coffee grown in containers. Cienc. Rural. 38(1):39-45. |
|
|
Munns R, James RA, Lauchli A (2006). Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot. 57(5):1025-1043. |
|
|
Munns R, Tester M (2008). Mecanisms of salinity tolerance. Annu. Rev. Plant. Biol. 59(1):651-681. |
|
|
Nelson SC, Elevitch CR. (2006) Workshop manual to supplement Noni: The complete guide for consumers and growers for noni processing, marketing and field training Workshop for YAP. Permanent Agriculture Resources, Holualoa, Hawaii 18 p. |
|
|
Oliveira FA, Medeiros JF, Oliveira FRA, Oliveira MKT, Freire AG (2012). Sensibility of cotton crops to mepiquat chloride under saline conditions. Rev. Ciênc. Agron. 43(3):484-492. |
|
|
Rodrigues CRF, Silva EN, Moura RM, Anjos DC, Hernandez FFF, Viégas RA (2014). Physiological adjustment to salt stress in R. communis seedlings is associated with a probable mechanism of osmotic adjustment and a reduction in water lost by transpiration. Ind. Crops Prod. 54(1):233-239. |
|
|
Santos APS, Perez-Marin AM, Forero LFU, Moreira JM, Medeiros AML, Lima RCSA, Bezerra HA, Bezerra BG, Silva LL (2014). O Semiárido brasileiro riquezas, diversidades e saberes. Instituto Nacional do Semiárido, Campina Grande, Brazil. |
|
|
Silva EC, Nogueira RJMC, Araújo FP, Melo NF, Azevedo Neto AD (2008). Physiological responses to salt stress in young umbu plants. Environ. Exp. Bot. 63(1):147-157. |
|
|
Souto AGL, Cavalcante LF, Lima Neto AJ, Mesquita FO, Santos JB (2016). Biometrics in noni plants under irrigation with saline water and the leaching of salts from the soil. Rev. Ciênc. Agon. 47(2):316-324. |
|
|
Souto AGL, Cavalcante LF, Diniz BLMT, Mesquita FO, Nascimento JAM, Lima Neto AJ (2015). Saline water and bovine biofertilizer on yield and biomass allocation in noni plant (Morinda citrifolia L.). Rev. Bras. Pl. Med. 17(2):340-348. |
|
|
Souto AGL, Cavalcante LF, Nascimento JAM, Mesquita FO, Lima Neto AJ (2013). Behavior of noni plants (Morinda citrifolia L.) to salinity of irrigation water in soil with bovine biofertilizer. Rev. Irriga. 18(3):442-453. |
|
|
Souza MCMR, Lacerda CF, Amorim, AV, Menezes, AS (2014). Influence of organic matter and the environment in growth and in solute accumulation in Morinda citrifolia submitted to salinity. Rev. Bras. Frutic. 36(3):704-712 |
|
|
Tavakkoli E, Rengasamy P, Mcdonald GK (2010). High concentrations of Na+ and Cl- ions in soil solution have simultaneous detrimental effects on growth of faba bean under salinity stress. J. Exp. Bot. 61(15):4449-4459. |
|
|
Taiz L, Zeiger E (2013). Fisiologia vegetal. Artmed, Porto Alegre, BR 918 p. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


