Review

ABSTRACT
Under natural conditions, the plants are, usually, surrounded by a series of potential enemies. They had created strategies of defense against pathogens and herbivores' attacks, allowing its perpetuation throughout evolution. These defense mechanisms are closely associated with the synthesis of secondary metabolites, which are also worldwide used in several areas of industry. This bibliographic review was conducted aiming to better understand how plants synthesize these substances in response to biotic or abiotic stimuli. The results discussed here revealed that synthesis of plant secondary metabolites is dependent on changes in several metabolic pathways, being often directly associated with the primary metabolism. Injury by herbivores or pathogens, temperature, CO2 levels, solar radiation and drought, are the factors related to the environment that express the most significant signs of inhibition or synthesis of plant secondary metabolites. Global climatic changes recently observed can affect the metabolic pathways network and, consequently, secondary metabolites synthesis. Nowadays, genomic tools have been useful alternatives that are leading to a new revolution of plant breeding, allowing the overexpression or inhibition of these substances. Some limitations and challenges to be achieved upon the dynamics of secondary metabolite synthesis in plants are presented.
Key words: Jasmonic acid, transcription factors, inhibition, synthesis.
INTRODUCTION
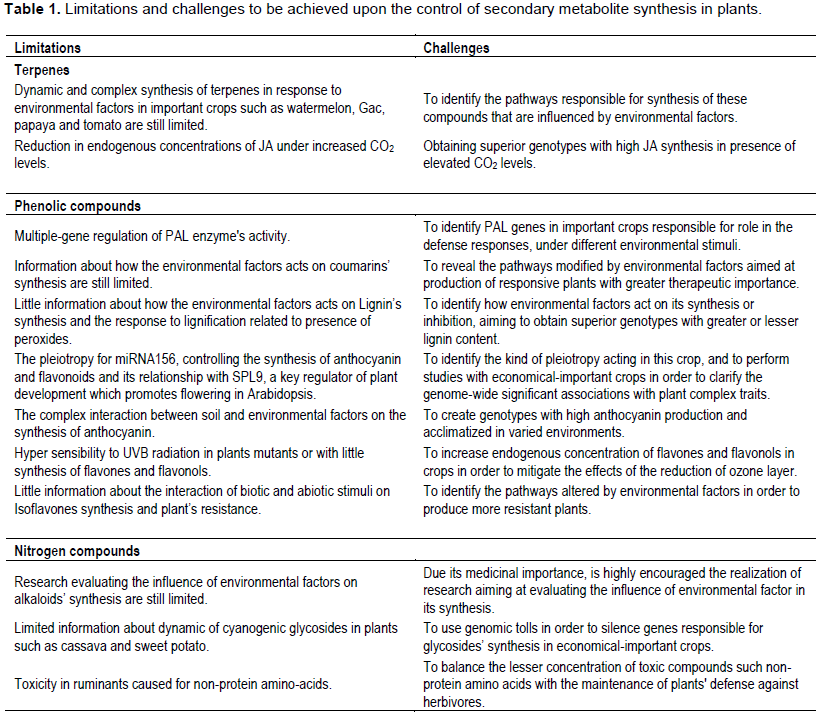
SECONDARY METABOLITES

CONFLICT OF INTERESTS
REFERENCES
|
Adamczyk B, Kitunen V, Smolander A (2013). Response of soil C and N transformations to condensed tannins and different organic N-condensed tannin complexes. Appl. Soil Ecol. 64:163-170. |
|
|
Aerts RJ, De Luca V (1992). Phytochrome is involved in the light-regulation of vindoline biosynthesis in Catharanthus. Plant Physiol. 100(2):1029-1032. |
|
|
Al-Qudah MA, Saleh AM, Al-Jaber HI, Tashtoush HI, Lahham JN, Abu Zarga MH, Abu Orabi ST (2015). New isoflavones from Gynandriris sisyrinchium and their antioxidant and cytotoxic activities. Fitoterapia 107:15-21. |
|
|
Arimura G, Ozawa R, Shimoda T, Nishiokd T, Boland W, Takabayashi J (2000). Herbivory-induced volatiles elicit defence genes in lima bean leaves. Nature 406(6795):512-515. |
|
|
Blanche CA, Lorio JPL, Sommers RA, Hodges JD, Nebeker TE (1992). Seasonal cambial growth and development of loblolly pine: Xylem formation, inner bark chemistry, resin duct and resin. For. Ecol. Manage. 49(1-2):151-165. |
|
|
Blum E, Liu K, Mazourek M, Yoo E Y, Jahn M, Paran I (2002). Molecular mapping of the C locus for presence of pungency in Capsicum. Genome 45(4):702-705. |
|
|
Bortesi L, Fischer R (2015). The CRISPR/Cas9 system for plant genome editing and beyond. Biotechnol. Adv. 33(1):41-52. |
|
|
Bozokalfa MK, Esiyok D, Turhan K (2009). Patterns of phenotypic variation in a germplasm collection of pepper (Capsicum annuum L.) from Turkey. Span. J. Agric. Res. 7(1):83-95. |
|
|
Budovská M, Pilátová M, Varinská L, Mojžiš J, Mezencev R (2013). The synthesis and anticancer activity of analogs of the indole phytoalexins brassini 1-methoxyspirobrassinol methyl ether and cyclobrassinin. Bioorg. Med. Chem. 21(21):6623-6633. |
|
|
Chan BG, Waiss AC, Lukefahr M (1978). Condensed tannin an antibiotic chemical from Gossypium hirsutum. J. Insect. Physiol. 24(2):113-118. |
|
|
Chang TL, Chiang HY, Shen JY, Lin SW, Tsai PJ (2015). Phenolic compounds stage an interplay between the ubiquitin–proteasome system and ubiquitin signal autophagic degradation for the ubiquitin-based cancer chemoprevention. J. Funct. Foods 17:857-871. |
|
|
Cheng Y, Liu L, Zhao G, Shen C, Yan H, Guan J, Yang K (2015). The effects of modified atmosphere packaging on core browning and the expression patterns of PPO and PAL genes in "Yali" pears during cold storage LWT. Food Sci. Technol. 60(2:2):1243-1248. |
|
|
Chini A, Fonseca S, Fernández G, Adie B, Chico JM, Lorenzo O, Micol JL (2007). The JAZ family of repressors is the missing link in jasmonate signaling. Nature 448(7154):666-671. |
|
|
Clausen TP, Provenza FD, Burritt EA, Reichardt PB, Bryant JP (1990). Ecological implications of condensed tannin structure: A case study. J. Chem. Ecol. 16(8):2381-2392. |
|
|
Consortium TTG (2012). The tomato genome sequence provides insights into fleshy fruit evolution. Nature 485(7400):635-641. |
|
|
Dar TA, Uddin M, Khan MMA, Hakeem KR, Jaleel H (2015). Jasmonates counter plant stress: A Review. Environ. Exp. Bot. 115:49–57. |
|
|
Darvill AG, Albersheim P (1984). Phytoalexins and their elicitors-a defense against microbial infection in plants. Ann. Rev. Plant Phys. 35(1):243-275. |
|
|
De Jong F, Hanley S J, Beale MH, Karp A (2015). Characterization of the willow phenylalanine ammonia-lyase (PAL) gene family reveals expression differences compared with poplar. Phytochemistry 117:90-97. |
|
|
De Luca V, Fernandez JA, Campbell D, Kurz WGW (1988). Developmental regulation of enzymes of indole alkaloid biosynthesis in Catharanthus roseus. Plant Physiol. 86(2):447-450. |
|
|
De Luca V, Laflamme P (2001). The expanding universe of alkaloid biosynthesis. Curr. Opin. Plant Biol. 4(3):225-233. |
|
|
Ejike CECC, Gong M, Udenigwe CC (2013). Phytoalexins from the Poaceae: Biosynthesis function and prospects in food preservation. Food Res. Int. 52(1):167-177. |
|
|
Espinosa RR, Inchingolo R, Alencar SM, Rodriguez-Estrada MT, Castro IA (2015). Antioxidant activity of phenolic compounds added to a functional emulsion containing omega-3 fatty acids and plant sterol esters. Food Chem. 182:95-104. |
|
|
Facchini PJ (2001). Alkaloid biosynthesis in plants: biochemistry cell biology molecular regulation and metabolic engineering applications. Annu. Rev. Plant Physiol. Plant Mol. Biol. 52(1):29-66. |
|
|
Facchini PJ, Bird DA, St-Pierre B (2004). Can Arabidopsis make complex alkaloids? Trends Plant Sci. 9(3):116-122. |
|
|
Farmer EE, Ryan CA (1990). Interplant communication: airborne methyl jasmonate induces synthesis of proteinase inhibitors in plant leaves. Proc. Natl. Acad. Sci. USA. 87(19):7713-7716. |
|
|
Fits L, Memelin J (2000). ORCA3: A jasmonate-responsive transcriptional regulator of plant primary and secondary metabolism. Science 289(5477):295-297. |
|
|
Franceschi VR, Krokene P, Christiansen E, Krekling T (2005). Anatomical and chemical defenses of conifer bark against bark beetles and other pests. New Phytol. 167(2):353-376. |
|
|
Gaj T, Gersbach CA, Barbas CF (2013). ZFN, TALEN and CRISPR/Cas-based methods for genome engineering Trends. Biotechnology 31(7):397-405. |
|
|
Gou JY, Felippes FF, Liu CJ, Weigel D, Wang JW (2011). Negative regulation of anthocyanin biosynthesis in Arabidopsis by a miR156-Targeted SPL transcription factor. Plant Cell 23(4):1512-1522. |
|
|
Gumul D, Korus J, Achremowicz B (2007). The influence of extrusion on the content of polyphenols and antioxidant/antiradical activity of rye grains (Secale cereale L.). Acta Sci. Pol. Technol. Alimen. 6:103-111. |
|
|
Gundlach H, Müller MJ, Kutchan TM, Zenk MH (1992). Jasmonic acid is a signal transducer in elicitor-induced plant cell cultures. Proc. Natl. Acad. Sci. USA 89(6):2389-2393. |
|
|
Gurung T, Techawongstien S, Suriharn B, Techawongstien S (2011). Impact of environments on the accumulation of capsaicinoids in Capsicum spp. HortScience 46(12):1576-1581. |
|
|
Hall DE, Zerbe P, Jancsik S, Quesada AL, Dullat H, Madilao LL, Yuen M, Bohlmann J (2013). Evolution of conifer diterpene synthases: diterpene resin acid biosynthesis in lodgepole pine and jack pine involves monofunctional and bifunctional diterpene synthases. Plant Physiol. 161(2):600-616. |
|
|
Heleno SA, Martins A, Queiroz MJRP, Ferreira ICFR (2015). Bioactivity of phenolic acids: metabolites versus parent compounds: A review. Food Chem. 173:501-513. |
|
|
Hernández V, Hellín P, Fenoll J, Flores P (2015). Increased temperature produces changes in the bioactive composition of tomato depending on its developmental stage. J. Agric. Food Chem. 63(9):2378-2382. |
|
|
Homoki JR, Nemes A, Fazekas E, Gyémánt G, Balogh P, Gál, F, Remenyik J (2016). Anthocyanin composition antioxidant efficiency and α-amylase inhibitor activity of different Hungarian sour cherry varieties (Prunus cerasus L.). Food Chem. 194:222–229. |
|
|
Hopke J, Donath J, Blechert S, Boland W (1994). Herbivore-induced volatiles: The emission of acyclic homoterpenes from leaves of Phaseolus lunatus and Zea mays can be triggered by a β-glucosidase and jasmonic acid. FEBS Lett. 352(2):146-150. |
|
|
Julsing MK, Koulman A, Woerdenbag HJ, Quax WJ, Kayser O (2006). Combinatorial biosynthesis of medicinal plant secondary metabolites. Biomol. Eng. 23(6):265-279. |
|
|
Kaushik P, Andújar I, Vilanova S, Plazas M, Gramazio P, Herraiz FJ, Brar NS, Prohens J (2015). Breeding vegetables with increased content in bioactive phenolic acids Molecules 20(10):18464-18481. |
|
|
Kovac A, Wegrzy JL, Parr G, Hol C, Bruenin GE, Loopstr CA, Hartigan J, Yandell M, Langley CH, Korf I, Neal DB (2010). The Pinus taeda genome is characterized by diverse and highly diverged repetitive sequences. BMC Genomics 11:420. |
|
|
Kurth C, Welling M, Pohnert G (2015). Sulfated phenolic acids from Dasycladales siphonous green algae. Phytochemistry 117:417-423. |
|
|
Li J, Ou-Le TM, Raba R, Amundson RG, Last RL (1993). Arabidopsis Flavonoid Mutants Are Hypersensitive to UV-B Irradiation. Plant Cell 5(2):171-179. |
|
|
Li L, Shen QP, Liu CB, Wang Y, Yao JJ, Zhang T, Zhang FM, He P, Shi XX, Liu ZH, Miao MM, Yang GY (2015). Isoflavones from the leaves of Nicotiana tabacum and their anti-tobacco mosaic virus activities. Phytochem. Lett. 13:156-159. |
|
|
Li T, Jia KP, Lian HL, Yang X, Li L, Yang HQ (2014). Jasmonic acid enhancement of anthocyanin accumulation is dependent on phytochrome A signaling pathway under far-red light in Arabidopsis. Biochem. Biophys. Res. Commun. 454(1):78-83. |
|
|
Logemann E, Parniske M, Hahlbrock K (1995). Modes of expression and common structural features of the complete phenylalanine ammonia-lyase gene family in parsley. Proc. Natl. Acad. Sci. USA 92(13):5905-5909. |
|
|
Maldonado AM, Doerne P, Dixo RA, Lam CJ, Camero RK (2002). A putative lipid transfer protein involved in systemic resistance signalling in Arabidopsis. Nature 419(6905):399-403. |
|
|
Martellini F, Giorni E, Colzi I, Luti S, Meerts P, Pazzagli L, Gonnelli C (2014). Can adaptation to metalliferous environments affect plant response to biotic stress? Insight from Silene paradoxa L. and phytoalexins. Environ. Exper. Bot. 108:38-46. |
|
|
Mayer KF, Waugh R, Brown JW, Schulman A, Langridge P, Platzer M, Fincher GB, Muehlbauer GJ, Sato K, Close TJ, Wise RP, Stein N (2012). A physical genetic and functional sequence assembly of the barley genome. Nature 491(7426):711-716. |
|
|
McSweeney CS, Collins EMC, Blackall LL, Seawright AA (2008). A review of anti-nutritive factors limiting potential use of Acacia angustissima as a ruminant feed. Anim. Feed. Sci. Technol. 147(1-3):158-171. |
|
|
Moreira X, Sampedro L, Zas R (2009). Defensive responses of Pinus pinaster seedlings to exogenous application of methyl jasmonate: Concentration effect and systemic response. Environ. Exper. Bot. 67(1):94-100. |
|
|
Moreira X, Zas R, Sampedro L (2012). Differential allocation of constitutive and induced chemical defenses in pine tree juveniles: a test of the optimal defense theory. PLoS ONE 7(3):e34006. |
|
|
Nabity PD, Zavala JA, DeLucia EH (2013). Herbivore induction of jasmonic acid and chemical defences reduce photosynthesis in Nicotiana attenuate. J. Exper. Bot. 64(2):685-694. |
|
|
Niggeweg R, Michael AJ, Martin C (2004). Engineering plants with increased levels of the antioxidant chlorogenic acid. Nat. Biotechnol. 22(6):746-754. |
|
|
Nose M, Bernards MA, Furlan M, Zajicek J, Eberhardt TL, Lewis NG (1995). Towards the specification of consecutive steps in macromolecular lignin assembly. Photochemistry 39(1):71-79. |
|
|
Obaid N, Kortschot MT, Sain M (2016). Lignin-Based Foaming Materials Lignin. in Polymer Composite. Elsevier pp. 217-230; 217–232. |
|
|
Ohta Y (1960). Physiological and genetical studies on the pungency of Capsicum II Pungency under various growing conditions. Seiken Ziho.11:73-77. |
|
|
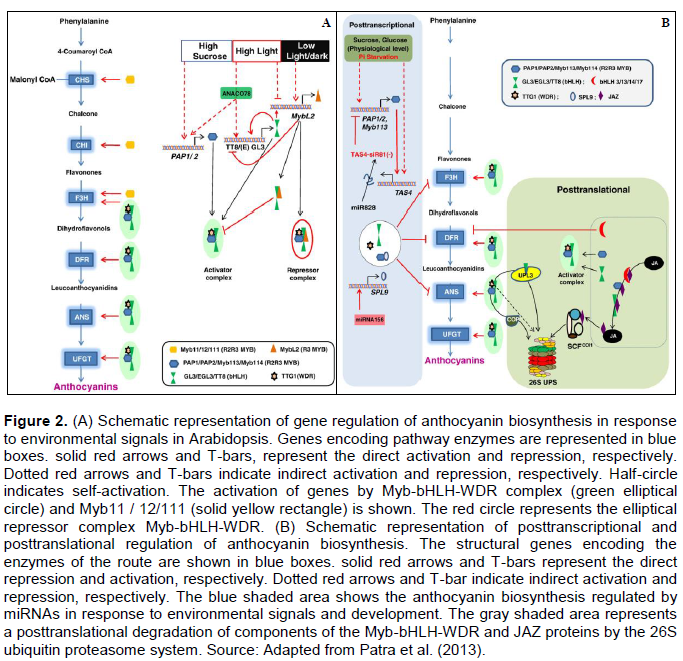
Patra B, Schluttenhofer C, Wu Y, Pattanaik S, Yuan L (2013). Transcriptional regulation of secondary metabolite biosynthesis in plants. BBA-Gene Regul. Met. 1829(11):1236-1247. |
|
|
Poulton J E (1990). Cyanogenesis in plants. Plant Physiol. 94(2):401-405. |
|
|
Prohens J, Rodríguez-Burruezo A, Raigón MD, Nuez F (2007). Total phenolic concentration and browning susceptibility in a collection of different varietal types and hybrids of eggplant: implications for breeding for higher nutritional quality and reduced browning. J. Am. Soc. Hortic. Sci. 132(5):638-646. |
|
|
Project IRGS (2005). The map-based sequence of the rice genome. Nature 436(7052):793-800. |
|
|
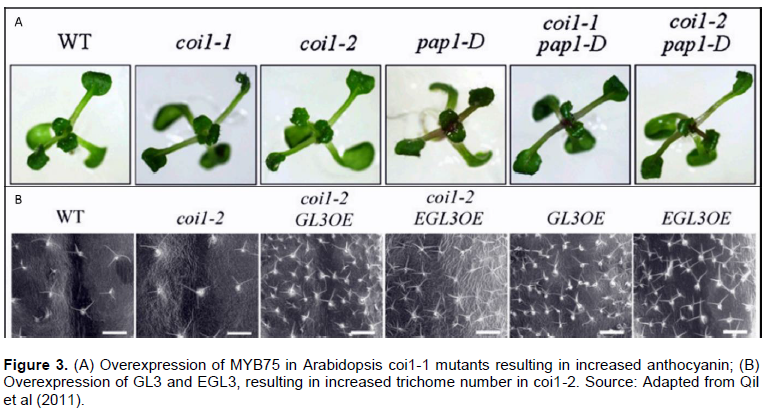
Qi T, Song S, Ren Q, Wu D, Huang H, Chen Y, Fan M, Peng W, Ren C, Xie D (2011). The Jasmonate-ZIM-Domain proteins interact with the WD-Repeat/bHLH/MYB complexes to regulate jasmonate-mediated anthocyanin accumulation and trichome initiation in Arabidopsis thaliana. The Plant Cell 23(5):1795-1814. |
|
|
Qiu Z, Guo J, Zhu A, Zhang L, Zhang M (2014). Exogenous jasmonic acid can enhance tolerance of wheat seedlings to salt stress. Ecotox. Environ. Safe 104:202-208. |
|
|
Rajendran L, Ravishankar GA, Venkataraman LV, Prathiba KR (1992). Anthocyanin production in callus cultures of Daucus carota as influenced by nutrient stress and osmoticum Biotech. Lett. 14(8):707-712. |
|
|
Robil JLM, Tolentino VS (2015). Histological localization of tannins at different developmental stages of vegetative and reproductive organs in Medinilla magnifica (Melastomataceae). Flora – Morphology, Distribution, Funct. Ecol. Plant 217:82-89 |
|
|
Rodríguez-García A, Martín JA, López R, Mutke S, Pinillos F, Gil L (2015). Influence of climate variables on resin yield and secretory structures in tapped Pinus pinaster Ait. in central Spain. Agric. Forest. Meteorol. 202:83-93. |
|
|
Rouholami S, Zahed B, Nazarian-Firouzabad F, Sae A (2015). Expression analysis of anthocyanin biosynthesis key regulatory genes involved in pomegranate (Punica granatum L.). Scientia Horticultura 186:84-88. |
|
|
Ruel JJ, Ayres MP, Lorio J, Peter L (1998). Loblolly pine responds to mechanical wounding with increased resin flow. Can. J. For. Res. 28(4):596-602. |
|
|
Saheb DN, Jog JP (1999). Natural fiber polymer composites: a review. Adv. Polym. Sci. 18(4):351-363. |
|
|
Shulaev V, Silverman P, Raskin I (1997). Airborne signalling by methyl salicylate in plant pathogen resistance. Nature 385(6618):718-721. |
|
|
Simon JR, Martin DL (1973). The effects of l-2:4-diaminobutyric acid on the uptake of gamma-aminobutyric acid by a synaptosomal fraction from rat brain. Arch. Biochem. Biophys. 157(2):348-355. |
|
|
Singh A, Pandey B, Kumari S, Agrawal M (2015). Nitrogen availability modulates CO2-induced responses of Catharanthus roseus: Biomass allocation, carbohydrates and alkaloids profile. J. Appl. Res. Med. Aromat. Plants 2:160-167. |
|
|
Siritunga D, Sayre R (2004). Engineering cyanogen synthesis and turnover in cassava (Manihot esculenta). Plant Mol. Biol. 56(4):661-669. |
|
|
Sivankalyani V, Feygenberg O, Diskin S, Wright B, Alkan NN (2016). Increased anthocyanin and flavonoids in mango fruit peel are associated with cold and pathogen resistance. Postharv. Biol. Technol. 111:132–139. |
|
|
Skalicka-Woźniak K, Orhan IE, Cordell GA, Nabavi SM, Budzyńska B (2015). Implication of coumarins towards Central Nervous System disorders. Pharmacol. Res. 103:188-203. |
|
|
Soriano IR, Rilye IT, Potter MJ, Bowers WS (2004). Phytoecdysteroids: A Novel Defense Against Plant-Parasitic Nematodes. J. Chem. Ecol. 30(10):1885-1899. |
|
|
Spencer DF, Ksander GG (1990). Influence of temperature, light and nutrient limitation on anthocyanin content of Potamogeton gramineus L. Aquatic Bot. 38(4):357-367. |
|
|
Stuiver MH, Custers JHHV (2001). Engineering disease resistance in plants. Nature 411(6839):865-868. |
|
|
Sudha G, Ravishankar GA (2003). Influence of methyl jasmonate and salicylic acid in the enhancement of capsaicin production in cell suspension cultures of Capsicum frutescens. Curr. Sci. 85(8):1212-1216. |
|
|
Sun Y, Guo H, Zhu-Salzman K, GF (2013). Elevated CO2 increases the abundance of the peach aphid on Arabidopsis by reducing jasmonic acid defenses. Plant Sci. 210:128-140. |
|
|
Sung Y, Chan Y-Y, Ni-Lu T (2005). Capsaicin biosynthesis in water-stressed hot pepper fruits. Bot. Bull. Acad. Sin. 46:35-42. |
|
|
Taiz L, Zeiger E (2010). Plant Physiology, Fifth ed. Sinauer Associates Inc., Massachusetts. |
|
|
Taofiq O, Calhelha RC, Heleno S, Barros L, Martins A, Santos-Buelga C, Queiroz MJRP, Ferreira ICFR (2015). The contribution of phenolic acids to the anti-inflammatory activity of mushrooms: Screening in phenolic extract individual parent molecules and synthesized glucuronated and methylated derivatives. Food Res. Int. 76(3):821-827. |
|
|
Thakur A, Singla R, Jaitak V (2015). Coumarins as anticancer agents: A review on synthetic strategie mechanism of action and SAR studies. Eur. J. Med. Chem. 101:476-495. |
|
|
Thines B, Katsir L, Melotto M, Niu Y, Mandaokar A, Liu G, Nomura K, He SY, Howe GA, Browse J (2007). JAZ repressor proteins are targets of the SCFCOI1 complex during jasmonate signaling. Nature 448(7154):661-665. |
|
|
Trapp S, Croteau R (2001). Defensive Resin Biosynthesis in Conifers. Annu. Rev. Plant. Physiol. Plant. Mol. Biol. 52(1):689-724. |
|
|
Veitch GE, Boyer A, Ley SV (2008). The Azadirachtin Story. Angew. Chem. Int. Ed. 47(49):9402-9429. |
|
|
Vranová E, Coman D, Gruissem W (2012). Structure and dynamics of the isoprenoid Pathway Network. Mol. Plan. 5(2):318-333. |
|
|
Wang C, Gu F, Chen J, Yang H, Jiang J, Du T, Zhang J (2015a). Assessing the response of yield and comprehensive fruit quality of tomato grown in greenhouse to deficit irrigation and nitrogen application strategies. Agric. Water Manage. 161:9-19. |
|
|
Wang Q, Li Y, Sun F (2015b). Tannins improve dough mixing properties through affecting physicochemical and structural properties of wheat gluten proteins. Food Res. Int. 69:64-71. |
|
|
Wink M (1988). Plant breeding: importance of plant secondary metabolites for protection against pathogens and herbivores. Theor. Appl. Genet. 75(2):225-233. |
|
|
Winkel BSJ (2004). Metabolic Channeling in Plants. Ann. Rev. Plant Biol. 55(1):85-107. |
|
|
Woo JW, Kim J, Kwon SI, Corvalán C, Cho SW, Kim H, Kim SG, Kim ST, Choe S, Kim JS (2015). DNA-free genome editing in plants with preassembled CRISPR-Cas9 ribonucleoproteins Nat. Biotechnol. 33(11):1162-1164. |
|
|
Wu J, Wang Z, Shi Z, Zhang S, Ming R, Zhu S, Khan MA, Tao S, Korban SS, Wang H, Chen NJ, Nishio T, Xu X, Cong L, Qi K, Huang X, Wang Y, Zhao X, Wu J, Deng C, Gou C, Zhou W, Yin H, Qin G, Sha Y, Tao Y, Chen H, Yang Y, Song Y, Zhan D, Wang J, Li L, Dai M, Gu C, Wang Y, Shi D, Wang X, Zhang H, Zeng L, Zheng D, Wang C, Chen M, Wang G, Xie L, Sovero V, Sha S, Huang W, Zhang S, Zhang M, Sun J, Xu L, Li Y, Liu X, Li Q, Shen J, Wang J, Paull RE, Bennetzen JL, Wang J, Zhang S (2013). The genome of the pear (Pyrus bretschneideri Rehd.). Genome Res. 23(2):396-408. |
|
|
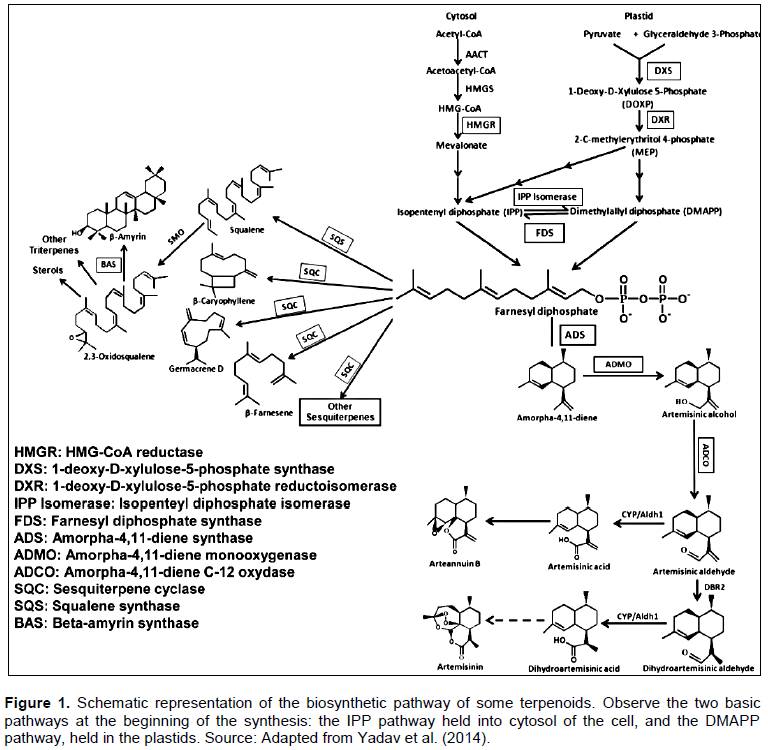
Yadav RK, Sangwan RS, Sabir F, Srivastava AK, Sangwan NS (2014). Effect of prolonged water stress on specialized secondary metabolite peltate glandular trichome and pathway gene expression in Artemisia annua L. Plant. Physiol. Biochem. 74:70-83. |
|
|
Yan Z, Li X, Chen J, Tam NFY (2015). Combined toxicity of cadmium and copper in Avicennia marina seedlings and the regulation of exogenous jasmonic acid. Ecotoxicol. Environ. Saf. 113:124-132. |
|
|
Yang Y, Huang S, Han Y, Yuan H, Gu C, Wang Z (2015). Environmental cues induce changes of steviol glycosides contents and transcription of corresponding biosynthetic genes in Stevia rebaudiana. Plant Physiol. Biochem. 86:174-180. |
|
|
Yu ZX, Li JX, Yang CQ, Hu WL, Wang LJ, Chen XY (2012). The Jasmonate-Responsive AP2/ERF Transcription Factors AaERF1 and AaERF2 Positively Regulate Artemisinin Biosynthesis in Artemisia annua L. Mol. Plant 5(2):353–365. |
|
|
Zewdie Y, Bosland PW (2000). Evaluation of genotyp environment and genotype-by-environment interaction for capsaicinoids in Capsicum annuum L. Euphytica 111(3):185-190. |
|
|
Zhang C, Huang Z (2013). Effects of endogenous abscisic acid, jasmonic acid, polyamine and polyamine oxidase activity in tomato seedlings under drought stress. Sci. Hortic. 159:172-177. |
|
|
Zhang C, Jia H, Wu W, Wang X, Fang J, Wang C (2015). Functional conservation analysis and expression modes of grape anthocyanin synthesis genes responsive to low temperature stress. Gene 574(1):168-177. |
|
|
Zhou C, Lin H, Ge X, Niu J, Wang J, Wang Y, Chen L, Huang Z, Yu W, Tan X (2015). The Effects of dietary soybean isoflavones on growt innate immune response hepatic antioxidant abilities and disease resistance of juvenile golden pompano Trachinotus ovatus. Fish Shellfish Immunol. 43(1):158-166. |
|
|
Zucker WV (1983). Tannins: Does Structure Determine Function? An Ecological Perspective. Am. Nat. 121(3):335-365. |
|
|
Zungu MM, Downs CT (2015). Effects of tannins on fruit selection in three southern African frugivorous birds. Behav. Process. 111:84-89. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


