Full Length Research Paper

ABSTRACT
This study aimed to differentiate and identify the best soil field capacity for good plant growth and optimization of yield. Tomato is an herbaceous crop that needs an adequate amount of water for growth and yield optimization. This experiment was conducted in pots in a greenhouse to examine the effect of water stress on the growth, yield, and shelf-life of five tomato varieties namely Padma F1, Cobra F1, Symbal F1, Titanium F1 and Nkansah GH. This research was conducted at the project site of the NEIP Envirodome greenhouse at the Dawhenya irrigation scheme in Ghana, from January 2017 to May 2019 for 5 cropping cycles. The tomato varieties were subjected to soil field capacity of 80-100FC%, 70-75FC%, 60-65FC% and 50-55FC% with three replications in a randomized complete block design. Plant height, stem diameter, internode length, leaf relative water content, stomata conductance, transpiration rate, yield, and shelf-life were the parameters measured to compute the effect of water stress on the different tomato varieties. Results from this study revealed that water stress decreased significantly leaf relative water content, stomata conductance, and transpiration rate at p<0.05. In conclusion, moderate water stress at field capacity 60-65 FC% resulted in optimizing plant morphological characteristics, physiological response, yield, shelf-life, and total soluble salt.
Key words: Water stress, tomato, crop physiology, crop morphology, yield, stomatal conductance, transpiration rate, relative leaf water content.
INTRODUCTION
Globally, water for irrigation is increasingly becoming limited because of global climatic change and the aggressive exploitation of natural resources (soil, water, and biodiversity). According to Fischlin et al. (2007), global warming is expected to increase water shortage by increasing evapotranspiration and the incidence and intensity of drought from 1 to 30% by 2100. Climate change is making water stress become a severe problem mostly limiting plant growth in arid and semi-arid regions (Fischlin et al., 2007; Knapp et al., 2001; Trenberth et al., 2015). Irrigation scheduling is a management strategy to improve the amount and time of irrigation water application as it contributes to water use efficiency and irrigation profitability (Alordzinu et al., 2017). Effective irrigation scheduling considers many factors, such as crop water requirements, crop growth stage, crop species, and variety, climatic factors, and soil physical and chemical properties that determine its water holding capacity. Furthermore, the crop's root mass and depth, affect the crop’s evapotranspiration process. The evapotranspiration rate of crops is similar to its water requirements, and this is the amount of water that the crop needs to compensate for the water lost through evapotranspiration (ET) (Allen et al., 1998; Nouri et al., 2013; Yadav and Sharma, 2016). Plants use several physiological adaptive mechanisms such as hormonal changes, cellular or molecular adaptive mechanisms to survive prolonged water stress (Bohnert and Jensen, 1996; Pedrol et al., 2000; Yadav and Sharma, 2016). Water stress is a major setback to global food production and food security (Hamouda et al., 2019; Lakmali et al., 2015; Nuruddin et al., 2003). Plant ontology is very sensitive to unplanned water stress because of the detrimental effect it has on growth, fruit quality, fruit size, and shelf-life especially when the plant reaches its peak physiological development (Shan and Zhao, 2015). Tomatoes (Solanum lycopersicum) are herbaceous vegetables from the Solanaceae family. It is an essential and the second most popular vegetable which is widely cultivated worldwide especially in arid and semi-arid regions. According to Bjarnadottir (2019) fresh tomato contributes significantly to the human diet because of its nutritional values, it is rich in vitamins A and C, water, protein, carbohydrate, ascorbic acid, sugar, fiber, fat, etc. Espinoza et al. (2017) reported that timely identification of water stress symptoms in crops is very vital to maintain good plant growth; the most common symptom of water stress in tomato is wilting. Tomato plants require a well regulated supply of water throughout the growing period for optimal quality and higher yield as tomato does not like “too dry or wet feet”, substantial reduction in plant vegetative growth, leaf relative water content, stomata conductance, yield, and other physiological and morphological plant characteristics may be as a result of severe water stress (Giuliani et al., 2017; Kirnak et al., 2001) however, plants may yield less fruit (weight, size, and number per cluster per plant) and quality when exposed to severe water stress. Tomato plants develop more root mass in soil with water content somewhat less than field capacity (Nuruddin et al., 2003), also, Liu et al. (2019) and Nyabundi and Hsia (2009) reported that tomato under water stress at field conditions inhibits its growth vegetatively but enhances fruit quality and number of fruits per plant.
This research is to examine the effect of water stress on the growth, yield, and shelf-life of tomato grown in pots in greenhouse.
MATERIALS AND METHODS
Study area
This research was carried out in Ghana at the Dawhenya irrigation scheme as a pot experiment conducted in the NIEP Envirodome greenhouse. Geographical location 0010’W 504’N with a mean annual temperature of 26.6°C, mean annual rainfall of 809 mm, and mean annual humidity is 81%. These experiments were conducted from 2017 to 2019 (5 growing cycles).
Test crop varieties
Five tomato varieties were used namely; Cobra F1 from Agriseed GH, Tythenium F1from Dzingoff GH, Symbal F1 from Agriseed GH, Padma F1 from East-West seeds, and Nkansah GH from the University of Ghana Crop Research Center.
Soil data
The soil type used in this research was the Akuse series. The soil was collected at a depth of 30 cm using a 200 cc core cutter from the Dawhenya irrigation scheme farmland. Soil samples collected were air-dried, rounded, and sieved with a 2 mm sieve to remove all debris and this was used to determine the physical and chemical properties as used by Osei et al. (2017). The soil physical and chemical properties of the sampled soil in August 2017 are shown in Tables 1 and 2.
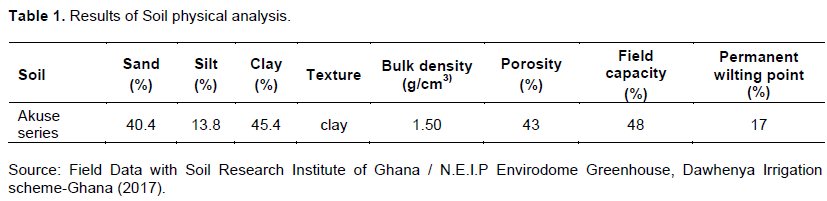
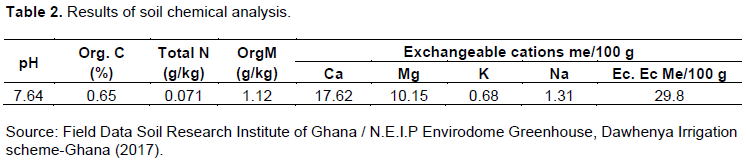
Soil moisture measurement
IMKO, Trime-Pico TDR was used to determine the soil moisture content in each pot. The instrument was calibrated and moisture readings were taken directly by inserting the instrument into the desired depth of soil through access tubes that were already installed in holes.
The volume of water applied to soil to reach each soil FC limit was calculated based on the following equation by Jackson (1982) and Martínez-Fernández et al. (2015).
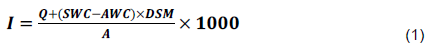
Where: (I) is the irrigation water (mm), Q is the volume of ponded water SWC is the water content on the required of the soil (%), AWC is the actual water of the soil when irrigating (%), DSM is the dry soil mass (kg), A is the pots surface area (mm2).
Soil drainability
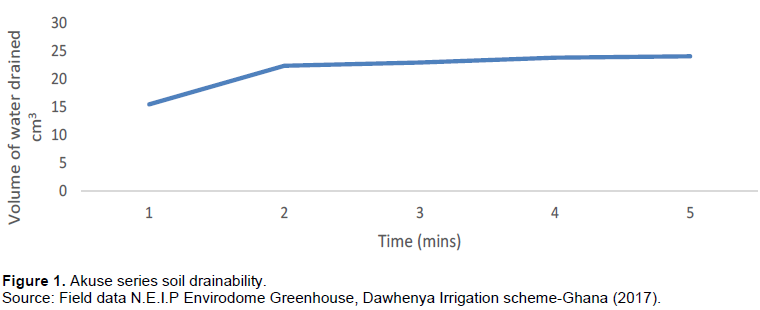
It is the volume of water that drains through the soil voids per unit time. When the soil pore volume of water is removed (or added) when the water table is lowered or raised in response to gravity without evaporation drainable porosity is said to occur (Osei et al., 2017). The soil drainability for this soil is shown in Figure 1.
Crop data
Morphological parameters
The physical and external structure of plants that were used to estimate water stress effect on the growth of tomato include plant height (Ph) (cm) which was measured by a meter stick with the zero mark placed at the ground level of the pot to the highest point of the plant, this measurement was carried out every three days and the value recorded for the entire growing season, plant height was calculated using growth formula (Poorter et al., 2012).

Where S1 is the first measurement, S2 is the second measurement and T is the number of days between each measurement. Stem Diameter (DS) (mm) was measured around the base of the plant using calipers and Internode Length (LI) (cm) measured between trusses using meter rule.
Physiological parameter
These parameters are responsible for the growth, development, and production of economic yield by the crop. Leaf Relative Water Content (LRWC), Transpiration rate (E), and Stomata Conductance were the physiological parameters used to estimate the water stress effects on the growth, yield, and shelf-life of tomato.
Leaf relative water content (LRWC)
This is an important indicator to estimate the amount of water contained in plants because it reflects the steady relationship between water available in plant leaf tissue and water lost through transpiration (Chakroun et al., 2015; Hamouda et al., 2019; Van Loon, 1981; Lugojan and Ciulca, 2011). This parameter was used to evaluate the existing water content of the sampled leaf tissue relative to the highest amount of water the leaf tissue can hold at full turgidity. The fresh mass of the sample leaves was weighed and recorded; the leaves were immersed in distilled water in a Petri dish. The immersed leaves were removed from the petri dish and the surface water was whipped cleaned and weighed to obtain the turgid leaf mass. The Samples were oven-dried at 70°C for 48 h to constant weight. Leaf relative water content was calculated using the equation by Pieczynski et al. (2013),

Where, LRWC %= Leaf Relative Water Content, FM= fresh leaf mass, DM= dry leaf mass, TM= turgid leaf mass.
Stomata conductance (gl)
This parameter measures the rate of water vapor evolving through the plant leaf stomata and this is a function of the aperture of the stomata, the size of the stomata, and stomata density. Transpiration reduces with the reduction of stomatal conductance (Massonnet et al., 2007). If plant leaf size reduces it may bring about stomatal closure hence lowering stomatal conductance (Damour et al., 2010). Three plants were sampled from each treatment and fully matured leaves from this plant were also sampled for data collection, handheld leaf porometer was used to measure the stomatal conductance in mmol/m2s after every irrigation application on a full bright day as this indicator is dependent on light.
Transpiration rate (E)
It is the loss of water from plants' stomata in the form of water vapor. This was estimated by the gravimetric method because the volume of water added to the soil and the volume of water lost was known by weighing the pot with the plant before irrigating and weighing after irrigating with a weighing scale with an accuracy of ±5 g. The measurements were taken for all the five growing seasons from 2017 to 2019 and this was compared to the moisture values from the moisture meter for consistency. Transpiration rates were calculated using the water balance approach. Kirnak et al. (2001), reported that the appropriate technique for estimating the transpiration rate of plants grown in pots is gravimetric because the volumes of water applied to the plant root zone in the pot and volumes of water drained from the pots is known.
Crop water stress index (CWSI)
The crop water stress values were calculated according to (Idso, 1982; Ihuoma and Madramootoo, 2019; Jackson et al., 2004) procedure, CWSI measures crop canopy temperature by measuring the minimum canopy temperature expected under non-water- stress conditions and the maximum temperature under severe water stress. The non-water-stressed starting point for the lower limits for the canopy-air temperature difference (Tc-Ta) versus the Vapor Pressure Deficit (VPD) relationship was determined using the data collected from the control treatment (T1- 80 to100%) only. The higher limits were calculated using canopy temperatures of the fully stressed plants (T4-50 to 55 %). CWSI was calculated for all the treatments, using the equation proposed by (Idso, 1982).

Where, D1 is the maximum canopy and air temperature difference for a stressed crop (the maximum stressed baseline, °C), D2 the lower limit canopy, and air temperature difference for a well-watered crop (the non-water-stressed baseline, °C), Tc is the measured canopy surface temperature (°C), Ta is the air temperature (°C). According to Idso (1982), vapor pressure deficit is calculated as:
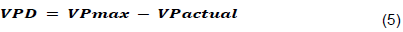
where: VP max is the maximum vapor pressure for a given air temperature and pressure, VP is the actual vapor pressure.
Water stress application
The crops were subjected to four (4) field capacity percentages treatment levels at FC range of 50-55% FC, 60-65%FC, 70-75% FC and 80-100%FC. To maintain the preferred percentage FC in the soil, water was added whenever the water level is lower than the required FC%, plastic mulch was used to cover the pots to avoid water loss through evaporation. IMKO, Trime-Pico TDR supplied by micromodultechnik GMBH with model code 100, HW:1,.2 FW:1, 120313 was used to measure the available moisture (volumetric water content) in the soil every day. This is useful for the estimation of the crop transpiration rate, relative water content, moisture content of the fruits, osmotic pressure, fruit quality, and brix of the fruit. The average volume of water applied per irrigation to maintain the soil %FC includes 250 ml/irrigation application for 80-100% F.C., 200 ml/irrigation application for 70-75% F.C. 150 ml/irrigation application for 60-65% F.C. 100ml/irrigation application for 50-55% F.C. with a scheduled irrigation interval of 3 days.
Nutrient application
Chicken manure was applied in the pot at the rate of 15 t/ha and inorganic fertilizer N, P, and K at the rate of 220-200-150 kg/ha, 70% of the total amount of potash, phosphorus, and 50% the nitrogen was incorporated during soil preparation and sterilization for the pot experiment. The remaining 30% of the P and K and 50% of N were applied as topdressing and supplementary nutrition was supplied as foliar at every 15days interval for 4 times in the growing season.
Measurement of crop yield, quality, and shelf-life parameters
The total number of fruits per plant was counted manually from 5 randomly sampled tomato plants from all five varieties and the averages were determined and recorded for the entire growing seasons. Harvesting of the matured tomato fruits at full ripening was carried out twice a week for 9 weeks for each growing season in all the 5 five cropping cycles (2017 to 2019) by manual hand picking.
All tomato fruits from selected pots with different water stress levels which were considered as marketable yield were weighed separately using an electronic weighing scale with a 0.001 g efficiency level. Total soluble solids (TSS) were measured twice on juice from 3 sampled tomato fruit from all five varieties, using a portable handheld refractometer Atago (ATAGO, Inc. Kirkland, WA, USA) and results were expressed as Brix value. Sampled fruits were sliced longitudinal into two halves (from stem end to calyx-end). The slice was squeezed to obtain the juice. 1 ml from the prepared fruit juice was placed onto the refractometer prism plate and the reading was taken and recorded, the prism plate was cleaned and dried After each test. Five tomato fruits from each variety from all the water stress treatments were sampled and stored on the shelf in a well-ventilated room at room temperature to determine the shelf-life, this was done for all the five growing seasons and the average yield, brix, and shelf-life determined. Data obtained from yield in terms of the number of fruits per plant was represented in the raw form in a table format to give a true reflection of the actual situation on the plant and the treatment imposed.
Experimental design
This research was conducted in ordinary time conditions as a pot experiment in a completely randomized block design (CRBD) with four treatments and three replications in the NIEP Envirodome Greenhouse. 15 L black plastic pots filled to 12 L with Akuse series black soil was used. The greenhouse was extensively examined for uniform distribution of light, temperature, air, and humidity to enable accurate data collection from experimental units. The data collected on plant morphology, physiology, yield, and shelf-life were subjected to Analysis of Variance (ANOVA) using SAS version 9.1. Significant means were separated using the Duncan Multiple Range Test (DMRT) at a = 0.05 probability level and the results were represented in tables and graphs.
RESULTS
Crop morphology
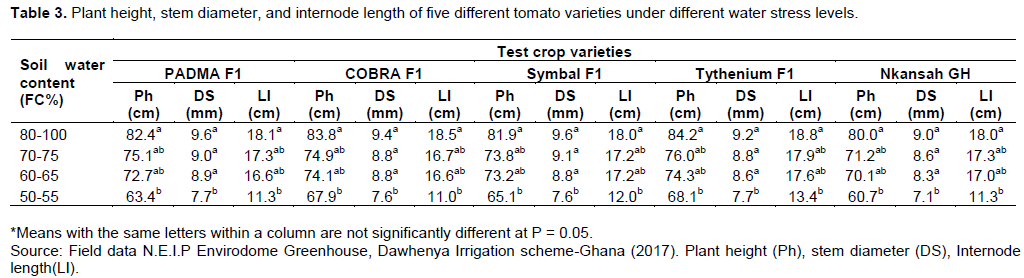
The data for each parameter was measured for all five tomato varieties and the means were determined and recorded separately. These means were compared with Duncan's least multiply range test (LSD) and this is shown in Table 3.
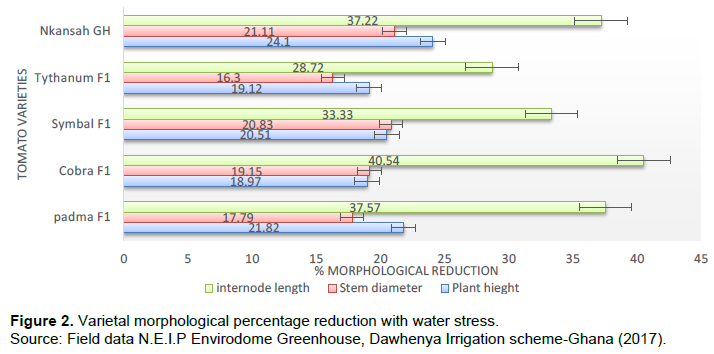
Varietal morphological percentage reduction with water stress
The five tomato varieties used as test crop for the research was further analyzed to compare the effect of water stress on their morphological development to determine the percentage reduction in plant height, stem diameter, and plant internode length (Figure 2).
Crops physiology
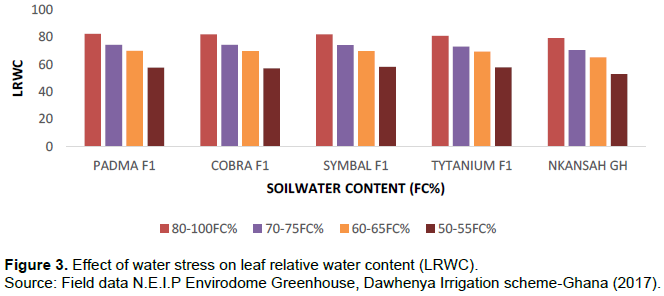
Leaf relative water content (LRWC)
LRWC was estimated using the equation explained by Pieczynski et al. (2013). Leaf Relative Water Content (LRWC (%) = [(FM–DM)/(TM–DM)] x100). The tested plant leaves were dependent on the pot soil field capacity (FC) levels which indicate a steady reduction (y = 92.78-4.84x; R2 = 85%) of the LRWC (%) with the respective water stress level (soil FC%) in the pots. The leaf relative water content was reduced by 25% in the most stressed plants (50-55% FC) compared to less water-stressed crops (80-100% of FC) and this is shown in Figure 3.
Stomata conductance
Plants with moderate to no water stress 60-65 FC % and above have generally higher stomata conductance ranging from 130 to 225 mmol/m2s compared to the plants with higher water stress 50-55 FC% having lower stomata conductance ranging from 100 to 112 mmol/m2s as shown in Table 4.

Transpiration rate (E)
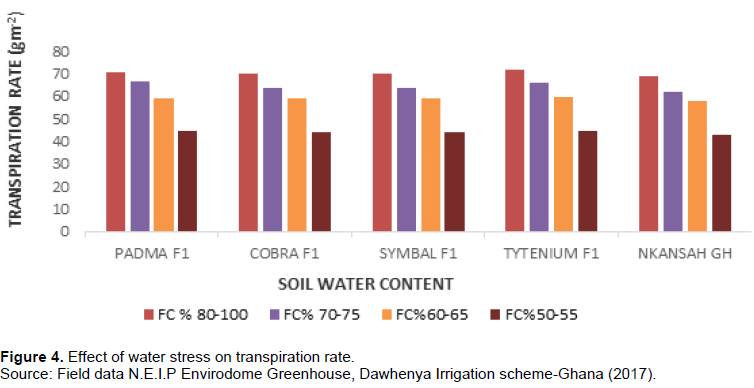
Transpiration rate for the high-water stress crops reduced by 44-45% at 50-55% FC compared to the less water stress crops of FC% ranging from 60-65% and above depending on the variety of tomato. The transpiration rate was very high for the crops that were subjected to no water stress 80-100% FC as shown in Figure 4.
Yield, brix, and shelf life parameters
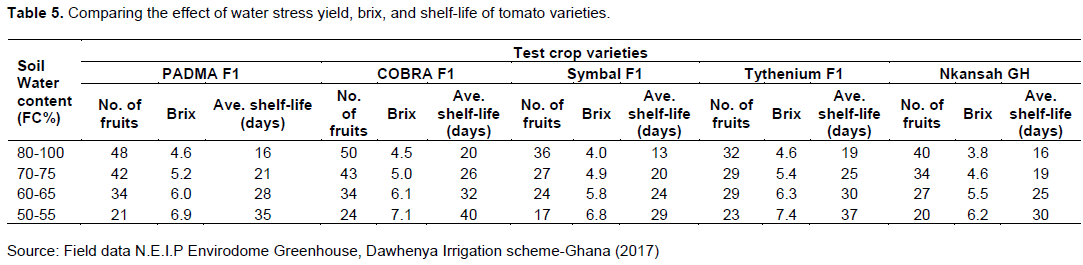
Results of tomato yield, brix, and shelf life for the five tomato varieties for the different water stress treatments are shown in Table 5.
DISCUSSION
Water stress on crop morphology as shown in Table 3 revealed that plants suffered water stress when soil water content becomes less than 60%, according to the results, plant height, stem diameter, and internode length decreased. This was evident in all five tomato varieties used in this experiment. These results confirm the results of (Nahar and Ullah, 2012; Nemeskéri et al., 2019), which indicated that plant height, internode length and stem diameter of water-stressed plants decreased compared to plants with no water stress. A similar result was also confirmed by (Giuliani et al., 2017) in deficit irrigation and partial root-zone drying techniques in processing tomatoes cultivated under Mediterranean climate conditions. Conversely when the soil water content was more than 60% plants had high internode length and stemdiameter which reflect changes in stem tissue hydration (Giuliani et al., 2017; Klepper et al., 1971). Furthermore, Fenglan et al. (2019) also confirmed these results from a report of changes in cell size and tissue hydration that, when soil water becomes lower than field capacity near-permanent wilting point, plant growth is usually decreased. Xu et al. (2016); Zeng et al. (2009) and Zeng et al. (2016) reported similar effects of water stress on dry matter of melon, other researchers also have proven that plant growth could be decreased with decrease soil water regardless of the complete maintenance of turgor in the growing regions as a result of the osmotic adjustment. Research has shown that when tomato plants are subjected to different levels of water stress under field conditions, vegetative growth is compromised (Nyabundi and Hsia, 2009). According to
Ibrahim (1990) and Giuliani et al. (2017), a greater reduction was seen in vegetative parts with decreased branch production, and correspondingly the main shoot became a more dominant component of the total shoot biomass. However, this is not always true as some other investigators such Kamrun et al. (2016) have observed and reported no significant difference in the growth of tomato plants subjected to different water stress levels. This argument was factual because the growth reduction of a plant will occur depending on the stage of growth, the time interval, or the duration of the stress.
It was observed from Figure 2 that the percentage reduction in plant height, stem diameter and internode length as affected by water stress revealed that Nkansah GH recorded the highest % reduction in terms of stem diameter and plant height with 21.11 and 24.10% respectively which is significantly different at p= 0.05 probability level from the other four varieties tested whereas Cobra F1 recorded the highest % reduction in internode length with 40.54%. These results confirm Harmanto et al. (2005); Liu et al., (2019) and Nyabundi and Hsia (2009) report which indicated that vegetative growth of tomato reduced under water stress at field conditions.
The application of water stress influences significantly the physiology of plant and plant water relations (LRWC, stomata conductance, and transpiration rate). The crop receiving the lowest water stress upheld higher plant water status (LRWC, stomata conductance, and transpiration rate). It is clearly shown from the result of this experiment that plants showed a variation in leaf relative water content, stomatal conductance, and transpiration rate when subjected to different levels of water stress this is represented in Figure 3 which shows the trend of how water stress affects LRWC. The result from this research confirmed the findings of Chen et al. (2015); El Jaafari (2000) and Tahar et al. (2010) who stated that the leaf relative water content decreased due to high water stress levels and increasing resistance to water flow in stems and leaves. Water stress also showed a similar trend of stomatal conductance and transpiration rate in tomatoes as shown in Table 4 and Figure 4.
These revealed how stomata conductance and transpiration rate are affected by crop water status on the five tomato varieties. Transpiration rate and stomatal conductance decreased with increasing water stress. This result was also consistent with Blanke and Cooke (2004); Kirnak et al. (2001); Nemeskéri et al. (2015) and Osakabe et al. (2013) who observed that prolonged water stress decreases plant moisture content which reduces the leaf stomata opening and transpiration rate. These authors suggested that an increase in the ratio of leaf surface mesophyll tissue somewhat increases crop water use efficiency by increasing photosynthesis more than it increases transpiration. Results from this research revealed that total soluble salt (Brix) that was estimated ranged from 3.8 to 7.4 of tomato fruits from both high-water stress and low-water stress treatment conditions among the five tomato varieties this results confirm the findings of Lahoz et al. (2016).
Effect of water deficit on the agronomical performance and quality of processing tomato. Water stress levels had a highly significant impact on the total soluble salt content of the fruits. High water stress at 50-55% soil field capacity caused lower yield (number of fruits per pant, size, etc.), highest soluble solids, with good shelf-life. The fruits obtained from plants with moderate water stress (60-65% and 70-75% soil field capacity) to high water stress (80-100% soil field capacity) yielded good marketable value with high soluble salts. This is a result of less water uptake by the plant into the fruit under stress which lowers the percentage of water in the fruit hence higher solute concentration and firmness of fruits. This also confirms the findings of Birhanu and Tilahun (2010), that plants subjected to water stress decrease the number and sizes of tomato fruits per plant. Nielsen (1994) and Zhang et al. (2017) also reported that low water stress resulted in a maximum yield of tomato raw product, best viscosity, and low soluble solids. This is not different from Zotarelli et al. (2009) who observed that tomato yield decreases when soil field capacity is reduced by 50%. Cantore et al. (2016) also reported that low plant water status increases total soluble salts and the shelf life of deficit irrigated tomato. Tomato plants subjected to severe water stress showed a significant reduction in yield but an increase in total soluble salts and shelf life compared to plants with no water stress, this is because plants under severe water stress did not receive the required amount of water to optimize their physiological process.
CONCLUSION
Plant morphological, physiological characteristics, yield (quantity and quality), and shelf-life were significantly influenced by different water stress levels (pot soil FC%). It can be concluded that tomato plants under 60-65 FC% performed well in terms of plant growth, yield (quality and quantity), and shelf-life.
This characteristic response of tomato plants to these water stress levels can be used as a protocol for tomato production to adequately optimize plant yield to ensure food and water security to efficiently utilize water resources in arid and semi-arid regions where water sources are scarce and of high demand by other water users.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Allen G, Pereira S, Raes D, Smith M (1998). Crop Evapotranspiration. FAO Irrigation and Drainage Paper 56:300-315. |
|
|
Alordzinu KE, Sam-Amoah LK, Owusu-Sekyere JD (2017). Assessing the performance of an irrigation scheme in Okyereko, Ghana. Journal of Agricultural Extension and Rural Development 9(12):270-282. |
|
|
Birhanu K, Tilahun K (2010). Fruit yield and quality of drip-irrigated tomato under deficit irrigation. African Journal of Food, Agriculture, Nutrition and Development 10:2141-2148. |
|
|
Bjarnadottir A (2019). Tomatoes 101 nutrition facts and health benefits: Healthline pp. 1-5. |
|
|
Blanke M, Cooke D (2004). Effects of flooding and drought on stomatal activity, transpiration, photosynthesis, water potential, and water channel activity in strawberry stolons and leaves. Plant Growth Regulation 42(2):153-160. |
|
|
Bohnert H, Jensen R (1996). Strategies for engineering water-stress tolerance in plants. Trends in Biotechnology 14(3):89-97. |
|
|
Cantore V, Lechkar O, Karabulut E, Sellami MH, Albrizio R, Boari F, Todorovic M (2016). Combined effect of deficit irrigation and strobilurin application on yield, fruit quality, and water use efficiency of "cherry" tomato (Solanum Lycopersicum L.). Agricultural Water Management. |
|
|
Chakroun H, Mouillot F, Hamdi A (2015). Regional equivalent water thickness modeling from remote sensing across a tree cover/LAI gradient in Mediterranean forests of Northern Tunisia. Remote Sensing 7(15):1937-1961. |
|
|
Chen S, Zhou Z, Andersen M, Hu T (2015). Tomato yield and water use efficiency-coupling effects between growth stage specific soil water deficits. Soil and Plant Science 65(5):460-469. |
|
|
Damour G,Thierry S, Cochard H, Urban L (2010). An overview of models of stomatal conductance at the leaf level. Plant, Cell and Environment 33(9):1419-1438. |
|
|
El Jaafari S (2000). Durum wheat breeding for abiotic stresses resistance: Defining physiological traits and criteria. Options Mediterranéennes 40(31):251-256. |
|
|
Espinoza CZ, Khot LR, Sankaran S, Jacoby PW (2017). High resolution multispectral and thermal remote sensing-based water stress assessment in subsurface irrigated grapevines. Remote Sensing 9(3):961. |
|
|
Fenglan M, Barrett-Lennard EG, Tian CY (2019). Changes in cell size and tissue hydration ('succulence') cause curvilinear growth responses to salinity and watering treatments in euhalophytes. Environmental and Experimental Botany 159(43):87-94. |
|
|
Fischlin A, Midgley G, Price J, Leemans R, Gopal B, Turley C, Rounsevell M, Dube P, Tarazona J, Velichko AA (2007). Climate Change Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. In Ecosystems, Their Properties, Goods and Services, 3:121-134. |
|
|
Giuliani MM, Nardella E, Gagliardi A, Gatta G (2017). Deficit irrigation and partial root-zone drying techniques in processing tomato cultivated under Mediterranean climate condition. Sustainability. |
|
|
Hamouda YA, Shaghaleh H, Sheteiwy M, Guoa X, Elshaikha N, Khand N, Oumaroue A, Rahima, S (2019). Impact of alternative wetting and soil drying and soil clay content on the morphological and physiological traits of rice roots and their relationships to yield and nutrient use efficiency. Agricultural Water Management 233(19):3-9. |
|
|
Harmanto VM, Salokhe VM, Babel MS, Tantau HJ (2005). Water requirement of drip irrigated tomatoes grown in greenhouse in tropical environment. Agricultural Water Management 71(3):225-242. |
|
|
Ibrahim AK (1990). Studies on growth and yield of chickpea. Ph.D. thesis University of Wales, Bangor, UK. |
|
|
Idso S (1982). Non-water stressed baselines: a key to measuring and interpreting plant water stress. Agricultural Meteorology 27(5):59-70. |
|
|
Ihuoma SO, Madramootoo CA (2019). Sensitivity of spectral vegetation indices for monitoring water stress in tomato plants. Computers and Electronics in Agriculture 163: 04860-87. |
|
|
Jackson R (1982). Canopy temperature and crop water stress. In Advances in Irrigation 1:43-85. |
|
|
Jackson T, Chen D, Cosh M, Li F, Anderson M, Walthall C, Doriaswamy P, Hunt E (2004). Vegetation water content mapping using Landsat data derived normalized difference water index for corn and soybeans. Remote Sensing of Environment 92(4):475-482. |
|
|
Kamrun MA, Eqani SAMAS, Bibi S, Xu S, Amna R, Monis MFH, Chaudhary HJ (2016). Bioaccumulation of nickel by E. sativa and role of plant growth promoting rhizobacteria (PGPRs) under nickel stress. Ecotoxicology and Environmental Safety 126(43):256-263. |
|
|
Kirnak H, Kaya C, Tas I, Higgs D (2001). The influence of water deficit on vegetative growth, physiology, fruit yield, and quality in eggplants. Bulgarian Journal of Plant Physiology 27(4):34-46. |
|
|
Klepper B, Browning V, Taylor H (1971). Stem diameter in relation to plant water status. Plant Physiology 48(6):683-685. |
|
|
Knapp AK, Briggs JM, Koelliker JK (2001). Frequency and extent of water limitation to primary production in mesic temperate grassland. Ecosystems 4(2):19-28. |
|
|
Lakmali WAS, Gunawardena ERN, Dayawansa NDK (2015). Comparative Performance Assessment of Major Irrigation Systems in Upper Deduru Oya Basin. Tropical Agriculture Research 26(2):343-354. |
|
|
Liu J, Hu T, Feng P, Wang L, Yang S (2019). Tomato yield and water use efficiency change with various soil moisture and potassium levels during different growth stages. PLoS ONE. |
|
|
Lugojan C, Ciulca S (2011). Evaluation of Relative Water Content in Winter Wheat. Journal of Horticulture, Forestry and Biotechnology 15(2):173-177. |
|
|
Martínez-Fernández J, González-Zamora A, Sánchez N, Gumuzzio A (2015). A soil water based index as a suitable agricultural drought indicator. Journal of Hydrology 522(3):265-273. |
|
|
Massonnet E, Serge R, Erwin D, Regnard J (2007). Stomatal regulation of photosynthesis in apple leaves evidence for different water-use strategies between two cultivars Catherine. Annals of Botany 100(6):1347-1356. |
|
|
Nahar K, Ullah S (2012). Morphological and Physiological Characters of Tomato (Lycopersicon esculentum Mill) Cultivars under Water Stress. Bangladesh Journal of Agricultural Research 37(2):355-360. |
|
|
Nemeskéri E, Molnár K, Vígh R, Nagy J, Dobos A (2015). Relationships between stomatal behavior, spectral traits, and water use and productivity of green peas (Pisum sativum L.) in dry seasons. Acta Physiologiae Plantarum, 37(5):34-42 |
|
|
Nemeskéri E, Neményi A, Bocs A, Pék Z, Helyes L (2019). Physiological factors and their relationship with the productivity of processing tomato under different water supplies. Water (Switzerland) 11(13):586. |
|
|
Nielsen DC (1994). Non-water stressed baselines for sunflowers. Agricultural Water Management 26(7):265-276. |
|
|
Nouri H, Beecham S, Hassanli AM, Kazemi F (2013). Water requirements of urban landscape plants: A comparison of three factor-based approaches. Ecological Engineering. |
|
|
Nuruddin MM, Madramootoo CA, Doods GT (2003). Effects of water stress at different growth stages on greenhouse tomato yield and quality. American Society of Horticultural Science. |
|
|
Nyabundi J, Hsia T (2009). Effects of water stress on growth and yield of field-grown tomatoes. H. Biomass partitioning between vegetative and productive growth. East African Agricultural and Forestry Journal 55(2):53-61. |
|
|
Osakabe Y, Arinaga N, Umezawa T, Katsura S, Nagamachi K, Tanaka H (2013). Osmotic stress responses and plant growth controlled by potassium transporters in Arabidopsis. PlantCell. |
|
|
Osei BY, Agyiri M, Aseidu EK, Agyarko K (2017). Assessment of the relative suitability of three different soil for dry season lettuce production. International Journal of Environment, Agriculture and Biotechnology 2(5):2456-2553. |
|
|
Pedrol N, Ramos P, Reigosa M (2000). Phenotypic plasticity and acclimation to water deficits in velvet-grass: a long-term greenhouse experiment. Changes in leaf morphology, photosynthesis, and stress induced metabolites. Journal of Plant Physiology 157(4):383-393. |
|
|
Pieczynski M, Marczewski W, Hennig J, Dolata J, Bielewicz D, Piontek P, Wyrzykowska A, Krusiewicz D, Strzelczyk-Zyta D, Konokpa- Postupolska D, Krzeslowska M, Jarmolowski A, Szweykowska- Kulinska Z (2013). Down-regulation of CBP80 gene expression as a strategy to engineer a drought-toletant potato. Plant Biotechnology Journal 11(4):459-469. |
|
|
Poorter H, Fiorani F, Stitt M, Schurr U, Finck A, Gibon Y (2012). The art of growing plants for experimental purposes?: a practical guide for the plant biologist. Functional Plant Biology. |
|
|
Shan C, Zhao X (2015). Lanthanum delays the senescence of Lilium longiflorum cut flowers by improving antioxidant defense system and water retaining capacity. Scientia Horticulturae, 197:516-520. |
|
|
Tahar B, Abdellah A, Abdul K, Al-Shoaibi A, Ali M (2010). Effect of water stress on growth and water use efficiency (WUE) of some wheat cultivars (Triticum durum) grown in Saudi Arabia. Journal of Taibah University for Science 3(1):39-48. |
|
|
Trenberth KE, Zhang Y, Fasullo JT (2015). Relationships among top?of?atmosphere radiation and atmospheric state variables in observations and CESM. Advancing Earth and Space Science 120(19): 10074-10090. |
|
|
Van Loon CD (1981). The effect of water stress on potato growth and development and yield. American Potato Journal 58(11):51-69. |
|
|
Xu S, Zhao Y, Wang M, Shi X (2016). Comparison of multivariate methods for estimating selected soil properties from intact soil cores of paddy fields by Vis-NIR spectroscopy. Geoderma 310(6):29-43. |
|
|
Yadav S, Sharma KD (2016). Molecular and Morphophysiological Analysis of Drought Stress in Plants. (E. C. Rigobelo (ed.); Plant Grow). IntechOpen. Available at: https://doi.org/DOI: 10.5772/65246 |
|
|
Zeng CZ, Bie ZL, Yuan BZ (2009). Determination of optimum irrigation water for drip irrigated muskmelon (Cucumis melo. L) in plastic greenhouse. Agricultural Water Management 96(4):595-602. |
|
|
Zeng R, Xu LH, Gao S, Ni X, Chen CL, Chen JC, Dai F (2016). One-step reverse transcription loop-mediated isothermal amplification assay for rapid detection of melon yellow spot virus. European Journal of Plant Pathology 145(1):119-124. |
|
|
Zhang C, Filella I, Liu D, Ogaya R, Llusià J, Asensio D, Peñuelas J (2017). Photochemical Reflectance Index (PRI) for Detecting Responses of Diurnal and Seasonal Photosynthetic Activity to Experimental Drought and Warming in a Mediterranean Shrubland. Remote Sensing. |
|
|
Zotarelli L, Dukes M, Scholberg JM, Munoz-Carpena R (2009). Tomato nitrogen accumulation and fertilizer use efficiency on a sandy soil, as affected by nitrogen rate and irrigation scheduling. Agricultural Water Management 96(8):1247-1258. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


