ABSTRACT
African mahogany (Khaya senegalensis) is a good option for the growing demand of the furniture industry since it is resistant to Hypsipyla grandella Zeller, a borrowing insect which causes damage to plants of the genus Swietenia in the American continent. The objective of this paper was to verify the effects of potassium doses on the growth of young African mahogany plants growing under two hydric conditions. The doses consisted of 0, 50, 100, 150, and 200 mg dm-3 of K in combination with hydric conditions of 30 and 70% of the soil maximum water holding capacity in pot, the treatment replications being distributed according with a completely random design and a 5 × 2 factorial scheme with 5 replications. At 180 days, the plants showed higher efficiency of potassium use, stomata conductance, transpiration, photosynthesis, height, diameter, leaf area, root dry matter, stem, and leaves at 70% of moisture. At the moisture of 30%, the potassium doses increased leaf area and leaf dry matter. Maximum water use efficiency was attained at 71 mg dm-3 of K. At 70% of soil moisture, increasing doses of K caused the reduction in N concentration in the leaves and a concomitant increment in phosphorus and potassium concentrations. Young African mahogany plants do not demand high doses of K as long as the other mineral necessities are met. Under hydric restriction, potassium increases the efficiency of water use and leaf biomass.
Key words: Khaya senegalensis, argisol, hydric restriction, potassium fertilization, photosynthesis.
Khaya senegalensis A.Juss. (African mahogany), a member of the Meliaceae family, is of natural occurrence in Africa, between latitudes of 15 and 18°N, along a line parallel to the equator including mainly Senegal, South Sudan, and the northern regions of Cameroon and Uganda (Lamprecht, 1990). It is found in river and swamp forests and dispersed through high pluvial precipitation forest savannas. Its habitat may be found between altitudes of 0 and 1800 m and rainfall precipitation between 700 and 1750 mm/year. It is the highest drought resistant species of the Khaya genus. Humid areas, though, favor its growth. It is the tallest among the savanna trees as well as the most valued noble wood since it is hard dense and long lasting (Orwa et al., 2009).
African mahogany is a good option for the growing demand of the furniture industry since it is resistant to Hypsipyla grandella Zeller, a borrowing insect which causes damage to plants of the genus Swietenia in the American continent (Pinheiro et al., 2011).
Nowadays, studying growth and development of plants under conditions of restricted water availability has become very important since conclusions from several research works referring to climatic changes in the world suggest that dry periods will increase in intensity, frequency, and duration (Misson et al., 2010). Under those conditions, the species, variety and source options as well as studies concerning physiology and nutrition of plants are of fundamental importance.
Swietenia macrophylla King in Hook. plants, under conditions of water deficiency, showed good control of transpiration and stomata conductance, in addition to having recovered leaf turgor after 12 h, showing a satisfactory tolerance to drought (Cordeiro et al., 2009). Khaya ivorensis A.Chev. also showed tolerance to moderate hydric deficiency since, 14 days after irrigation suppression, it recovered its hydric status three days after rehydration and, a little later, the gaseous exchanges (Albuquerque et al., 2013).
Potassium may influence the plant responses to water deficiency. It is related to the osmotic potential of plants since it influences cell expansion and ion transport in addition to being fundamental for the stomata movement. Potassium well-nourished plants use the soil water more efficiently whereas the ones with potassium deficiency have a poorer photosynthetic performance due to the irregular opening of the stomata which causes reduction in the amount of CO2 taken in (Prado, 2008).
It was verified in Eucalyptus grandis plantlets (going through their hardening period) growing under hydric stress, that fertilizing them with potassium brought about a reduction in the amount of water lost by the plant (Silva et al., 2004).
So, the objective of this paper was to evaluate the effect of doses of potassium on the growth of young African mahogany plants growing under two hydric conditions.
Description of the experimental environment
The experiment took place between August, 2013 and March, 2014. It was carried out in a 30 m long, 4.5 m high, and 5 m wide greenhouse which was covered by a screen capable of intercepting up to 30% of the sun light.
Seed provenance
Seeds of K. senegalensis were provided by the Vasconcelos Florestal company, which is found in the municipality of Monte Alto, state of São Paulo, Brazil. The origin of seeds is the Indian settlement of Tiakane, Burkina Faso (Africa), localized at 11° 11’ 10’’ of N latitude and 1° 12’ 14’’ of W longitude. Burkina Faso climate is tropical semi-arid with high temperatures with a mean annual temperature of 30°C and potential annual evapotranspiration of approximately 2000 mm. A long dry period (from October to May) and a short rainy one (from July to September) are the most important climatic characteristics of that region (Couteron and Kokou, 1997).
Experiment installation
The plants grew in black polyethylene 6.5 dm3 vases covered with newspaper to reduce excessive warming by solar radiation. These vases had their bottom covered with a 0.05 mm mesh to reduce soil losses.
A total of 5.5 dm3 of an Argisol was placed in each vase. Each soil sample was taken from a 20 cm of topsoil in the “São Gabriel” farm, in the municipality of Monte Alto (21° 17’ 16’’ S and 48° 28’ 33’’ W), state of São Paulo, Brazil, where, formerly, there were pastures. The soil granulometric analyses showed the following values: sandy texture with 8.6% of clay, 4.9% of silt, 30% of fine sand, and 56.5% of coarse sand. Soil fertility analyses, following Raij et al. (2001) procedures, were of P resin: 2 mg dm-3; OM: 10 g dm-3; pH (CaCl2): 4.7; K+: 1.0 mmolc dm-3; Ca2+: 9.0 mmolc dm-3; Mg2+: 5 mmolc dm-3; H + Al: 28 mmolc dm-3; CEC at the pH of 7: 43 mmolc dm-3 and basis saturation (V): 35%.
Taking into consideration the soil chemical analysis results, calcium carbonate and calcium magnesium were applied following the Ca:Mg proportion of 3 : 1 so as to raise the soil base saturation (V) up to 50%.
Making use of the simple superphosphate in its powder form, 80 mg dm-3 of P were applied to the soil and after that the soil was moistened so as to reach 70% of its total water retention capacity. Such condition was kept during the 30 days period of incubation (time required for soil reaction with lime and fertilizers applied). Fifteen days later, the following fertilization doses were applied to each vase: 25 mg dm-3 of N (ammonium sulfate), 0.25 mg dm-3 of B (boric acid), 0.5 mg dm-3 of Zn (zinc sulfate), and 0.5 mg dm-3 of Cu (copper sulfate). The following N (alternating the source of N between urea and ammonium sulfate) doses were applied on the soil surface: 25 mg dm-3 at 45 days after sowing, 25 mg dm-3 at 75 days, 50 mg dm-3 at 110 days, and 50 mg dm-3 at 130 days, totalizing the application of 225 mg dm-3 of N.
Experimental design
The treatment replications were distributed according to a randomized complete block design with five potassium doses (0, 50, 100, 150, and 200 mg dm-3 of K2O) and two levels of water availability: 30 and 70% of the soil total water holding capacity. These treatment combinations resulted in a 5 × 2 factorial scheme with five replications. Each replication consisted of one vase bearing two plants, totaling 100 plants in the experiment.
Conduct and evaluation of the experiment
The potassium to the plants was provided by potassium chloride (60% of K2O). The potassium dose was split in three times; the first one took place 30 days after period of incubation (20% of the treatment potassium concentration), the second, 60 days after incubation (40% of the treatment potassium concentration), and the third 120 days after incubation (40% of the treatment potassium concentration). Water to the soil was supplied so that the levels of 30 and 70% of the total soil retention capacity were reached.
Soil maximum water retention capacity was determined by weighing 10 vases with mesh in the base with 5.5 dm3 of soil in each one. The vases with dry soil were weighed and, after that, water was added so as to soak the soil. After a period of 48 h of drainage, the vases were again weighed. The soil maximum water retention capacity was considered to be the difference between the mass after drainage and the mass of the dry soil.
Four untreated seeds were sown per vase. The vases were covered with a screen which reduced to 50% the sun light reaching the soil surface during seed germination; moisture level in the soil was maintained for 35 days at 70%. At the end of that period, only two plants per vase were left. Starting 40 days after sowing, the procedures to bring about soil moisture variation begun.
In order to guarantee the levels of 30 and 70% of soil moisture during the experiment, the vases were daily weighed and the difference in weight from one day to the next was considered to be the needed amount of irrigation water.
At 180 days after germination, the concentrations of nitrogen, phosphorus, and potassium were measured in the plant leaves. The potassium use efficiency in the plant leaves was also determined. Nitrogen determination made use of the semi-micro Kjeldahl method after sulfuric digestion while the nitro-perchloride method was used to measure P and K contents. Phosphorus and potassium were measured by the metavanadate colorimetry and flame photometry methods, respectively, according to the methodology described by Bataglia et al. (1983).
The nitrogen, phosphorus and potassium contents in the leaves were calculated by multiplying the leaf dry matter amounts by the respective nutrient concentrations. These values were used to calculate the potassium use efficiency (PUE) in the leaves, according to the equation proposed by Siddiqi and Glass (1981). PUE = (leaves total dry matter)2/potassium concentration (g2 g-1) in the leaves.
Physiological processes such as stomata conductance, transpiration, and photosynthesis at 135, 142, 152, and 180 days after germination were evaluated. These evaluations were performed between 8 and 11 a.m. in totally expanded leaves in the median part of the third whorl of the composite leaf of a branch of the third verticil with the help of an IRGA (Infra Red Gas Analyzer) model LCpro-SD, a product of ADC BioScientific. Room temperature and relative humidity were measured; CO2 concentration was fixed at 380 ± 10 µmo m-2 s-1. FFFA (photosynthetically active photon flux) of 800 µmol m-2 s-1, corresponding to the maximum photosynthesis value found in a preliminary test with 6 plants, was used.
Each variable was represented by the mean value of the four performed evaluations.
Based on the mean photosynthesis and stomata conductance results, water use efficiency (WUE) was calculated: WUE = photosynthesis/stomata conductance.
At 180 days of germination, the chlorophyll index was evaluated with a portable chlorophyllimeter (CCM 200, Opti Science) and evaluations of survival, height, diameter, leaf area, root, stem and leaves dry matter mass, root/aerial part dry mass ratio were made.
Plant aerial part height was considered to be the length between the stem basis in contact with the soil and the apex of the youngest leaf pair. Diameter was measured 5 cm above soil surface.
Leaf area was measured with equipment named Li-Cor 3100. The leaves, the stem, branches and roots were separately placed inside bags and put to drying in a forced ventilation oven at 65 to 70°C untill a constant weight was reached.
Statistical analysis
The data were statistically analyzed with the AgroEstat program (Barbosa and Maldonado, 2011). Analysis of variance and polynomial regression analysis were made for the effects of potassium on measured variables.
African mahogany plants survival was of 100% in all treatments. Higher concentrations of nitrogen (41.67%) and potassium (21.17%) and lower concentration of phosphorus (6.7%) were detected in leaves when soil moisture content was of 30% in comparison with 70% (Table 1), this is probably due to the dilution effect since under the latter condition, the physiological characteristics of stomata conductance, transpirations and photosynthesis were higher (Table 1) with a consequent increase in biometric characteristics (Table 1).
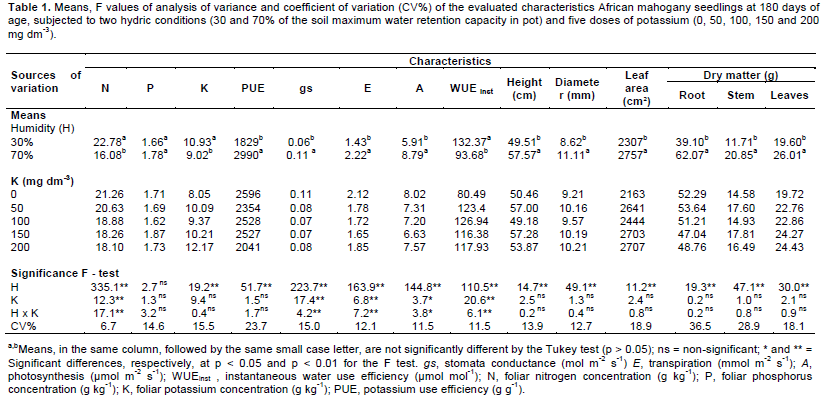
When potassium dose was increased at soil moisture of 70%, a tendency to decrease nitrogen concentration in the leaves was observed, and that variation could be adjusted by a third degree equation (Ŷ = 20.25 - 0.0149x - 0.0006x2 + 0.000003x3; R² = 1; p = 0.0352). This result confirms data found in a study of the interaction between nitrogen, phosphorus and potassium in other species when the highest doses of potassium resulted in the highest production and the highest concentrations of nitrogen were found in the plants that received the lowest doses of potassium (Macleod, 1969).
The data showed that phosphorus foliar concentration was influenced by potassium only at soil moisture of 30% and that the largest dose of potassium resulted in a linear increment in the concentration of phosphorus in the leaves (Ŷ = 1.46 + 0.002x; R² = 0.8778; p = 0.007).
As to the foliar concentration of potassium, it was linearly increased with potassium doses when soil moisture was of 30% (Ŷ = 9.236 + 0.017x; R² = 0.7505; p = 0.0004) and at 70% (Ŷ = 7.376 + 0.0164x; R² = 0.7409; p = 0.0005).
When soil moisture was 70%, potassium was used with efficiency 38.8% higher than that observed when soil moisture was of 30%. Applying potassium to the soil decreased soil efficiency of potassium dose at that soil moisture, the variation being adjusted by a third degree curve (Ŷ = 3425.2 - 27.409x + 0.3811x2 -0.0014x3; R² = 0.9996; p = 0.0153). This result confirms data found for Corymbia citriodora (Hook.) K.D. Hill and L.A.S. Johnson also subjected to different potassium doses and soil moisture levels, when a decreasing linear effect in the efficiency of potassium use by the leave was observed. This fact was supposed to be due to the conservation of the leaves dry matter as the concentration of potassium increased (Sá et al., 2014).
When soil moisture was of 30%, stomata conductance (45.5%), transpiration (35.6%) and photosynthesis rate (32.8%) were lower than at 70% (Figure 1). Eucalyptus clones under four irrigation regimes also showed reduction in stomata conductance and photosynthesis rate when water availability was reduced, probably as a consequence of the partial shutting of the stomata soon after the first morning hours (Tatagiba et al., 2007). When under conditions of low water availability, plants activate several mechanisms to avoid dehydration such as the reduction in transpiration. This response is achieved by the shutting of the stomata, which limits water vapor loss by reducing stomata pore although the providing of CO2 to the chloroplasts is also part of the mechanism (Taiz and Zeiger, 2013). The reduction in stomata conductance is, nonetheless, higher than that verified for carbon assimilation and this brings about an increment in the efficiency of water use by the plants. If the low availability of water persists for a long period, photosynthesis rate decreases and, as a consequence, plant growth is reduced (Lima et al., 2003).
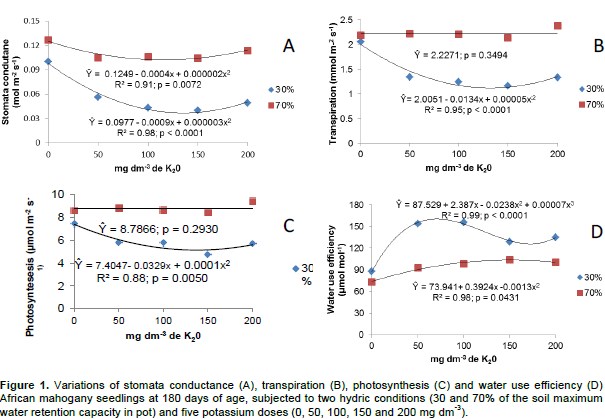
Increasing doses of potassium, when soil moisture content was of 30%, caused reductions in stomata conductance, transpiration and photosynthesis rate whereas when soil moisture content was of 70%, only reduction in stomata conductance was observed; these variations being explained by second degree equations (Figure 1), probably due to the participation of potassium in the regulation of the opening and shutting of the stomata, thus reducing the excessive loss of water by the plant under hydric stress (Taiz and Zeiger, 2013). This favorable effect of potassium was observed in Eucalyptus grandis W.Hill plantlets under moderate hydric stress promoting reduction of water loss by the plant (Silva et al., 2004).
When soil moisture content was of 30%, the plants showed higher efficiency in the use of water compared to 70% (Table 1). This result may be due to the reduction in stomata conductance being higher than that verified in carbon assimilation when soil moisture was of 30% which increases the efficiency of the instantaneous use of water (Pita et al., 2005). These results are concordant with those reported for six Eucalyptus species which were subjected to hydric stress (20% of the soil water holding capacity) for 10 weeks and this resulted in more severe reduction in stomata conductance than in photosynthesis rate (Merchant et al., 2007). Plants of Eucalyptus genotypes growing in soils treated with potassium made a more efficient use of water than plants growing in conditions of no restriction of water (Mendes et al., 2013).
Water use efficiency increased with the application of potassium both at soil moisture contents of 70 and 30%. The variation verified when soil moisture was of 70% is explained by a second degree equation and the maximum efficiency is estimated with the application of151 mg dm-3 of K. When soil moisture content was of30%, the variation is explained by a third degree equation and the highest efficiency was estimated by the application of 70.7 mg dm-3 of K (Figure 1).
Providing the plant with the adequate dose of potassium is of fundamental importance for the efficiency of water use since this nutrient is associated with characteristics that interfere in the efficiency of water use such as turgor regulation, opening and shutting of the stomata and transpiration (Taiz and Zeiger, 2013). Similar results were reported for Corymbia citriodora and Eucalyptus camaldulensis Dehnh. plants growing under water stress and fertilized with different doses of potassium. The highest doses resulted in increased water use efficiency and in the leaf relative water content (Teixeira et al., 1995).
Increasing potassium dose, when soil moisture content was of 70%, did not affect significantly plant height, diameter, leaf area and root, stem, and leaves dry matter of the plants (Table 1).
Potassium concentration in plant leaves which did not receive potassium fertilization was of 8.05 g kg-1, this being a value close to 9.0 g kg-1, which is considered adequate for African mahogany (Bevege et al., 2006). This suggests that the content of 1.0 mmolc dm-3 of K present in the soil before the experiment was started was sufficient for the initial phase of plant growth.
S. macrophylla (Brazilian mahogany) plantlets growth was not affected by potassium fertilization either, thus suggesting that this species is a K low-demanding one or that it converts efficiently potassium in biomass (Tucci et al., 2011). Omitting K from sowing fertilization of Copaifera langsdorffii (Rashed Tree or Salam Tree) and Hymenea courbaril (courbaril) did not interfere in plant height, diameter and aerial part dry matter. The Rashed Tree plants showed a tendency to increased leaf biomass when they did not receive potassium (Duboc, 1994).
This kind of behavior is likely to be common in species of tropical forests since Rashed Tree, Platycianus regnellii (‘pau-pereira’, in Brazil), and courbaril did not respond to fertilization with potassium, probably because they are slow-growing plants compared to fast-growing species, which do respond to potassium (Silva et al., 1997). So, at a soil moisture content of 70%, the absence of significant responses to potassium by the plants seems to be related to their low demand of that nutrient at their initial growth stages.
When growing in a soil moisture content of 70%, the plants were taller, had a larger diameter, a larger leaf area and plant components dry matter than those growing at 30% (Table 1). Differences in percentage between plant components the ones of plants growing at 70 and 30% were of 43.8% (stem dry matter), 37% (root dry matter), 24.6% (leaf dry matter), 22.4% (diameter), 16.3% (leaf area), and 14% (height). Similar results were reported for eucalyptus genotypes plants which, under various irrigation regimes, grew more than the non-irrigated ones (Mendes, 2011).
Under conditions of hydric stress, plants undergo important modifications in physiological and morphological processes which affect their capacity to withstand environmental adversities. Under such conditions, cell volume is reduced and, consequently, turgor pressure. Thus, activities depending on turgor pressure and cell expansion such as root growth in length and cell expansion are extremely sensitive to hydric stress (Taiz and Zeiger, 2013). Water deficiency may lead to plant growth paralysis or just to a reduction in plant growth (Vellini, 2007). On the other hand, leaf area reduction may be seen as a plant strategy to withstand drought since leaf area reduction leads to a transpiration reduction (Taiz and Zeiger, 2013).
Significant effects of potassium doses on leaf area were observed only when soil moisture content was of 30%. The variations in leaf area were verified to occur according to the linear equation (Ŷ = 1976.2 + 3.309x; R2 = 0.5889; p = 0.0184). The increment in leaf dry matter determined by increasing potassium doses, when soil moisture was of 30%, is described by the linear equation Ŷ = 16.72 + 0.0288x; R2 = 0.675; p = 0.0187.
Similar results were reported for eucalyptus clones growing under two hydric conditions and potassium doses in which only leaf area and leaf dry matter were significantly affected by potassium doses (Mendes et al., 2013). On the other hand, studies on the effects of water management and potassium fertilization on the behavior of eucalyptus plants at the rustification phase, showed no effect of potassium on the aerial part of the plant; only root dry matter was significantly influenced. Therefore, it is suggested more studies.
K. senegalensis plants, at their initial stage of development, demand low amounts of potassium as long the other nutritional elements are provided. Under restricted water conditions, potassium increases the efficiency of water use and leaf biomass.
The authors have not declared any conflict of interests
The study was sponsored by the Brazilian National Concil for Research (CPNPq).
REFERENCES
|
Albuquerque MPF, Moraes FKC, Santos RIN, Castro GLS de, Ramos EMLS, Pinheiro HA (2013). Ecofisiologia de plantas jovens de mogno-africano submetidas a déficit hídrico e reidratação. Pesqui. Agropecu. Bras. 48(1):9-16.
Crossref
|
|
|
|
Barbosa JC, Maldonado JW (2011). AgroEstat: sistema para análises estatísticas de ensaios agronômicos, versão 1.1.0.626. Jaboticabal: FCAV, Departamento de Ciências Exatas.
|
|
|
|
|
Bataglia OC, Furlani ANC, Teixeira JPF, Furlani PR, Gallo JR (1983). Métodos de análise química de plantas. Campinas: IAC 48 p.
|
|
|
|
|
Bevege DI, Nikles DG, Dickinson G, Skelton DJ (2006). Where to from here with R&D to Underpin Plantations of High-value Timber Species in the 'Seasonally-dry' Tropics of Northern Australia? In: WORKSHOP, 2006, Townsville. Kairi: Private Forestry North Queensland Association, 1 CD-ROM.
|
|
|
|
|
Cordeiro YEM, Pinheiro HA, Santos Filho BG dos, Corrêa SS, Dias Filho MB (2009). Physiological and morphological responses of Young mahogany (Swietenia macrophylla King) plants to drought. For. Ecol. Manage 258:1449-1455.
Crossref
|
|
|
|
|
Couteron P, Kokou K (1997). Woody vegetation spatial petterns in a semi-arid savanna of Burkina Faso, West Africa. Plant Ecol. 132:211-227.
Crossref
|
|
|
|
|
Duboc E (1994). Requerimentos nutricionais de espécies florestais nativas: Hymenaea courbaril L. var. silibocarpa (Haynee) Lee et Lang (Jatobá), Copaifera tangsdorffii Desf., (Óleo copaíba) e Peltopliorum dublum (Spreng) Taub. (Canafístula) Dissertação (Mestrado)- Escola Superior de Agricultura de Lavras, Lavras 68 p.
|
|
|
|
|
Lamprecht H (1990). Silvicultura nos trópicos: ecossistemas florestais e respectivas espécies arbóreas - possibilidades e métodos de aproveitamento sustentado. Rossdorf: Deutsche Gesellschaft für Technische Zusammenarbeit 343 p.
|
|
|
|
|
Lima WP, Jarvis P, Rhizopoulou S (2003). Stomatal responses of Eucalyptus species to elevated CO2 concentration and drought stress. Sci. Agric. 60(2):231-238.
Crossref
|
|
|
|
|
Macleod LB (1969). Effects of N, P, and K and their interactions on the yield and kernel weight of barley on hydroponic culture. Agron. J. 61:26-29.
Crossref
|
|
|
|
|
Mendes HSJ (2011). Comportamento de clones de eucalipto em resposta a disponibilidade hídrica e adubação potássica. Dissertação (Mestrado) – Faculdade de Ciências Agrárias e Veterinárias – Universidade Estadual Paulista. Jaboticabal. 43 p.
|
|
|
|
|
Mendes HSJ, Paula NF de, Scarpinatti EA, Paula, RC de (2013). Respostas fisiológicas de genótipos de Eucalyptus grandis x E. urophylla à disponibilidade hídrica e adubação potássica. Cerne 19(4):603-611.
Crossref
|
|
|
|
|
Merchant A, Callister A, Arndt S, Tausz M, Adams M (2007). Constrasting physiological responses of six Eucalyptus species to water déficit. Ann. Bot. 100(7):1507-1515.
Crossref
|
|
|
|
|
Misson L, Limousin JM, Rodriguez R, Letts LG (2010). Leaf physiological responses to extreme droughts in Mediterranean Quercus ilex Forest. Plant Cell Environ. 33(11):1898-1910.
Crossref
|
|
|
|
|
Orwa C, Mutual A, Kindt R, Jamnadass R, Antony S (2009). Agroforestree Database: a tree reference and selection guide version 4.0.
|
|
|
|
|
Pinheiro AL, Couto L, Pinheiro, DT, Brunetta JMFC (2011). Ecologia, silvicultura e tecnologia de utilização dos mognos-africanos (Khaya spp.). Viçosa: Sociedade Brasileira de Agrossilvicultura 102 p.
|
|
|
|
|
Pita P, Ca-as I, Soria F, Ruiz F, Toval G (2005). Use of physiological traits in tree breeding for improved yield in drought-prone environments. The case of Eucalyptus globulus. For. Syst. 14(3):383-393.
Crossref
|
|
|
|
|
Prado RM (2008). Nutrição de plantas. São Paulo: Editora UNESP. P. 407.
|
|
|
|
|
Raij B, Andrade JC, Cantarella H, Quaggio JA (2001). Análises químicas para avaliação da fertilidade de solos tropicais. Campinas: Instituto Agronômico 285 p.
|
|
|
|
|
Sá AFL, Valeri SV, Cruz MCP, Barbosa JC, Rezende GM, Teixeira MP (2014). Efeitos da aplicação de potássio e umidades do solo no crescimento de plantas de Corymbia citriodora. Cerne 20(4):645-651.
Crossref
|
|
|
|
|
Siddiqi MY, Glass ADM (1981). Utilization index: a modified approach to estimation and comparison of nutrient utilization efficiency in plants. J. Plant Nutr. 4:289-302.
Crossref
|
|
|
|
|
Silva IR, Furtini Neto AE, Curi N, Vale FR (1997). Crescimento inicial de quatorze espécies florestais nativas em resposta à adubação potássica. Pesqui. Agropecu. Bras. 32(2):205-212.
|
|
|
|
|
Silva MR, Klar AE, Passos JR (2004). Efeitos do manejo hídrico e da aplicação de potássio nas características morfofisiológicas de mudas de Eucalyptus grandis (Hill ex. Maiden). Irriga 9(1):31-40.
|
|
|
|
|
Taiz L, Zeiger E (2013). Fisiologia vegetal. 5. ed. Piracicaba, Artmed. 820 p.
|
|
|
|
|
Tatagiba SD, Pezzopane JEM, Reis EF (2007). Avaliação do crescimento de clones de Eucalyptus submetidos a diferentes manejos de irrigação. Cerne 1(13):1-9.
|
|
|
|
|
Teixeira PC, Leal PGL, Barros NF, Novais RF (1995). Nutrición potásica y relaciones em plantas de Eucalyptus spp. Bosque 16(2):61-68.
Crossref
|
|
|
|
|
Tucci CAF, Santos JZL, Silva Júnior CH, Souza, PA, Batista, IMP. Venturin N (2011). Desenvolvimento de mudas de Swietenia macrophylla em resposta a nitrogênio, fósforo e potássio. Floresta 41(3):471-490.
Crossref
|
|
|
|
|
Vellini ALTT (2007). Desempenho e divergência genética entre clones de Eucalyptus spp. em diferentes regimes de irrigação em casa de vegetação. Tese (Doutorado em Genética e Melhoramento de Plantas) - Universidade Estadual Paulista. Jaboticabal 95 p.
|
|



