Full Length Research Paper

ABSTRACT
Stripe (yellow) rust caused by Puccinia striiformis f.sp. tritici, is one of the major diseases of wheat in the world. Development and use of resistant wheat cultivars is the most economical and environmentally friendly solution in combating wheat stripe rust. Field experiments were carried out at two sites in Ethiopia (Kulumsa and Meraro) and seedling tests were conducted at KARC greenhouse during the 2015 cropping season to evaluate the response 192 elite spring bread wheat genotypes and eight checks to the prevailing races of stripe rust at adult plant stage and seedling stage. About 72.5 and 42.5% of the lines exhibited resistance to stripe rust during the field screening at Kulumsa and Meraro, respectively. Disease was more severe at the cooler site Meraro than Kulumsa. Eighteen genotypes at Kulumsa and 16 genotypes at Merao were almost immune to the disease (severity and AUDPC of zero). Seventy-two genotypes (36%) showed resistant reaction at both locations in field condition for adult plant stage (CI < 20). For seedling, 47% for mixed isolates and 31% for kubsa isolates showed resistance reaction responses to stripe rust disease based on coefficient of infection (CI).
Key words: Puccinia striiformis, resistance, susceptible, Triticum aestivum, genotypes.
INTRODUCTION
Wheat is one of the world’s leading and widely grown cereal crop serving as a staple food for more than 1/3 of the global population. With the rising global population and decreasing arable land, wheat production and yield improvement became crucial. Therefore, to fulfill the food demands of ever-growing population, the food produced in developing countries has to be enhanced by 70% till 2050 (Semenov et al., 2014).
Ethiopia is the largest wheat producer in Sub-Saharan Africa (FAOSTAT, 2018) and wheat is central to achieving food and nutrition security. About 5 million Ethiopian farmers produce 5.3 million tons of wheat across 1.8 million hectares of land under rain-fed conditions (CSA, 2020). Its popularity comes from the versatility of its use in the production of a wide range of food products, such as injera, breads, cakes, pastas, etc. Wheat ranks third in area coverage and total production after teff and maize. Although the productivity of wheat has increased in the last few years in Ethiopia, it is still low as compared to other wheat producing countries. The national average productivity is estimated to be 2.97 t ha-1 (CSA, 2020), which is by far below experimental yields of over 5 tons ha-1 (Mengistu et al., 2018). So, for national productivity reduction, there are different constraints from those with plant diseases among the major factors affecting the yield of wheat.
Among these plant diseases in wheat that significantly reduce wheat production are those caused by the rusts (yellow, stem and leaf) (Khan et al., 2013). Stripe (yellow) rust disease caused by Puccinia striiformis f.sp. tritici is one of the major diseases of wheat in the cool environments including in Ethiopia (Singh et al., 2000). Infection can occur anytime from the one-leaf stage to plant maturity, provided plants are still green (Chen, 2005). Damage of stripe rust depends on susceptibility of the variety, how early epidemic begins, the amount of stripe rust that develops and temperature during grain filling (Uauy et al., 2005). So, breeding for resistant cultivars is considered the most effective, economic and environmentally friendly approach to control stripe rust. Race-specific resistance with single resistance gene is often short lived. Gene pyramiding strategies deploying race-specific and non-race specific resistance genes is required to achieve durable resistance. However, detailed knowledge of resistance genes present in wheat cultivars is a prerequisite in resistance breeding (Reema et al., 2019).
In Ethiopia, major stripe rust epidemics occurred in 1970’s, 1980, 1988, 2000 and 2010 (Teklay et al., 2013). In 2010, more than 400,000 ha of wheat were affected which led to serious yield losses, though difficult to quantify. Most popular commercial bread wheat cultivars; Kubsa and Dashen were susceptible to stripe rust (Nazari, 2011) and it causes yield loss of 70-100% in Ethiopia. This has been clear in the breakdown of stripe rust resistance genes Yr9 in cultivars derived from “Veery” in 1980’s and Yr27 in 2000 and 2010 in widely grown cultivars derived from “Attila” cross such as PBW343 (India), Inquilab-91 (Pakistan), Kubsa (Ethiopia) and others in almost all CWANA (Central and West Asia and North African) countries (Solh et al., 2012). Hence, continuous search for new sources of resistance ahead of changing pathogen and pyramiding of more resistance genes in single cultivars is important to control stripe rust and to avoid the ‘boom and bust cycle’ of cultivar performance.
Field and seedling evaluation of the level of resistance of various genotypes and multi-locational disease testing of germplasm is used to obtain data to support breeding strategies aimed at broadening the genetic base (Khan et al., 2013). Stripe rust like the other rusts have complex life cycle that involve alternate hosts and several spores’ stages. New races continually surfaced-out due to rust ability to mutate and sexually recombine. Understanding wheat rust severity and coefficient of infection, along with identifying of effective genes and characterizing them would help to design future breeding schemes. Hence, the objective of this study was to screen and identify elite spring bread wheat genotypes for resistance to stripe rust for adult plant and seedling resistance.
Although these Yr genes have been identified in diverse wheat accessions, the race specificity of seedling resistance genes limits their efficacy against pathotypes (Kankwatsa et al., 2017). In contrast, APR is generally considered to be durable, but APR genes represent a minority of known resistance genes (Kankwatsa et al., 2017; Yuan et al., 2018). Therefore, enhancing the resistance of adult plants to cope with evolving races of Pst is the preferred strategy to breeding for resistance. The objectives of this study were to evaluate 192 ICARDA, wheat genotypes with 8 checks for stripe rust resistance at adult plant and seedling stage conditions and therefore, the resistance genotypes can be used for further manipulation in the wheat breeding program by incorporation into adapted cultivars to assess the variability to yellow rust resistance.
MATERIALS AND METHODS
Description of experimental locations
The field experiment was conducted at two locations in Arsi zone, Ethiopia; namely, Kulumsa and Meraro. These locations are hot spot for wheat stripe rust in Ethiopia. Kulumsa represents highland areas with an altitude of 2200 m a.s.l., mean annual rainfall of 820 mm with maximum temperature of 22.8°C and minimum temperature of 10.5°C. Meraro represents extreme highland and cold area and is located at an altitude of 2990 m.a.s.l. The mean annual rainfall of the site is 1196 mm, maximum temperature is 18.1°C while the minimum temperature is 5.7°C.
Experimental materials
A total of 192 elite spring bread wheat genotypes and 8 checks Pastor-2, Qimma-12, Attila-7, Kabowsh-1, SIDS-1, Debira, Goumria-3 and Hiddab were tested using alpha lattice design with two replications (Appendix Table 1). The checks were used to compare the resistance of these genotypes to stripe rust. The stripe rust spore was harvested and maintained from the field during the previous growing season (September-October 2014) and multiplied in the greenhouse using universal susceptible wheat cultivars (Morocco and Kubsa) during (July-September, 2014/15) and used for inoculating 192 elite spring bread wheat genotypes and the eight checks.
Seedling test
Four to five seeds of each genotype were planted in 7 cm x 7 cm x 7 cm plastic pots. Each pot was filled with a potting mix which consists of soil, sand and compost at a ratio of 2:1:1 (v/v/v). After one week of planting, when the first leaves were fully expanded, the seedlings were inoculated by spraying the most virulent and dominant varieties Kubsa/Attila and mixed isolates urediospores suspended in mineral oil using an atomizer. Inoculated plants were allowed to dry for 5 min, were fine-misted with water and placed in a wet plastic cage with a small amount of water at the bottom. The inoculated seedlings were incubated at 10°C for 24 h in a dew chamber with relative humidity close to 100%. Seedlings were transferred to a greenhouse with mean temperature of about 18°C at the Ethiopian Institute of Agricultural Research, Kulumsa Agricultural Research Center (KARC), Greenhouse Lab. Disease assessment was carried out on the 15th day after inoculation using 0–4 scale (McIntosh et al., 1995) based on the infection types. Low infection types (LITs = 0–2) were considered resistant, and infection type = 2+ as intermediate while high infection types (HITs = 3–4) were rated susceptible.
Field test
One hundred and ninety-two genotypes and 8 checks in this study were planted using an alpha lattice design in two replications in a plot size of 1 m length, planted in two rows with 0.2 m spacing between rows at Kulumsa and Meraro. The eight bread wheat cultivars that were used as checks were planted within intervals of twenty-four entries. Field managements and agronomic practices were carried out as recommended for each location. Spreader rows were planted as mixtures of universal susceptible bread wheat cultivars and the dominant varieties (Morocco and Kubsa) adjacent to the 192 elite genotypes and 8 checks on both sides of each block, bordering the trials to ensure production of sufficient inoculum to provide uniform stripe rust infection. The inoculation of spreader row was carried out during tillering stage by spraying method and during stem elongation stage by injection methods at 50 cm interval. Spraying of stripe rust on spreader row during tillering stage was done by mixing fresh stripe rust spore with water and then sprayed to spreader row using Knapsack sprayer. Stripe rust injection to spreader row was conducted by mixing stripe rust spore with pure water and was applied to the spreader row by injecting stem at stem elongation stage using injection syringe.
Disease severity was assessed according to the modified Cobb’s scale (Peterson et al., 1948). The genotype’s reaction response to the infection in the field was scored four times at 12 days interval starting from mid-September when disease symptom commenced up to the time when disease development progress ceased as ‘‘R’’ or resistant (small uredinia surrounded by chlorosis or necrosis); ‘‘MR’’ or moderately resistant (medium sized uredinia surrounded by chlorosis or necrosis); ‘‘MS’’ or moderately susceptible (medium large compatible uredinia without chlorosis and necrosis); and ‘‘S’’ or susceptible (large, compatible uredinia without chlorosis and necrosis) while the disease severity was scored in the 0 to 100-percent scale (Roelf et al., 1992). Thus, Yellow rust scores 10 MRMS means 10% severity of moderate resistant-to-moderately susceptible response while the 20MSS score indicates 20% severity of moderately susceptible-to-susceptible response and yellow rust score 60S suggests, 60% severity of susceptible type response. Finally, after the last disease score when the disease progress ceased, according to Stubbs (1986), the field severity data was converted to Coefficient of Infection (CI) by multiplying with constant values of response. After the last disease score when the disease progress ceased, according to Stubbs (1986), the disease severity data and host reaction response were combined to calculate the coefficient of infection (CI) following Pathan and Park (2006), by multiplying severity value with constant values of 0, 0.2, 0.4, 0.6, 0.8, or 1.0 for host response ratings of immune (I), resistant (R), moderately resistant (MR), intermediate (M), moderately susceptible (MS), or susceptible (S), respectively. Genotypes with coefficient of infections ranging from 0 to 20 were considered as resistant while 20 to 30, 30 to 40, 40 to 60 and 60 to 100 were moderately resistance, moderately susceptible, moderately susceptible to susceptible and susceptible, respectively based on the reaction of check cultivars.
Area under Disease Progress Curve (AUDPC) was calculated in order to compare the genotypes’ susceptibility and resistance. The AUDPC was calculated using the midpoint rule method (Campbell and Madden, 1990). The formula is:
AUDPC = Si=1n-1 [(ti+1 – ti)(yi + yi+1)/2],
Where “t” is time in days of each reading, “y” is the percentage of affected foliage at each reading and “n” is the number of readings. AUDPC was calculated by considering each disease severity score and the coefficient of infection that was taken four times.
RESULTS
Response of genotypes in field condition
Phenotypic variation for stripe rust was observed at both environments for infection types and level of severity for the 192 ICARDA elite spring bread wheat genotypes and eight susceptible checks. Terminal score ranged from 0 (immune) to 100 S (highly susceptible). Reaction response to stripe rust for these genotypes at Kulumsa and Meraro locations are summarized in Figures 1 and 2 and Appendix Table 1. More disease severity/pressure was observed at Meraro than at Kulumsa. The checks showed variable reaction responses from moderately resistant to susceptible and a severity level ranging from 10 to 100%. Some of the checks such as Attila-7 (Kubsa), Sids-1, Goumaria-3 and Hiddab exhibited high terminal severity (>50) at both sites while Pastor-2, Qimma-12, Kabowsh-1 and Debira showed lower terminal severity at Kulumsa and high terminal severity at Meraro. Five of the eight checks showed higher stripe rust severity level at Meraro than at Kulumsa (Appendix Table 2). The frequency of these elite spring bread wheat genotypes and the checks under different severity classes at Kulumsa and Meraro is presented on Figures 1 and 2, respectively, according to the coefficient of infection (CI) score. At Kulumsa, of the total 192 elite spring bread wheat genotypes and eight checks evaluated, 145 (72.5%) including 2 checks (Pastor-2 and Kabowsh-1) exhibited resistance reaction response (CI = 0 to 20); nine genotypes including one check (Debira) (4.5%) were moderately resistant (CI = 20 to 30); sixteen (8%) with one check (Qimma-12) were moderately susceptible (CIs = 30 to 40), twelve (6%) genotypes with Attila-7 were moderately susceptible to susceptible (CI = 40 to 60) and 18 (9%) including the remaining three checks were susceptible (CI= 60 to 100).
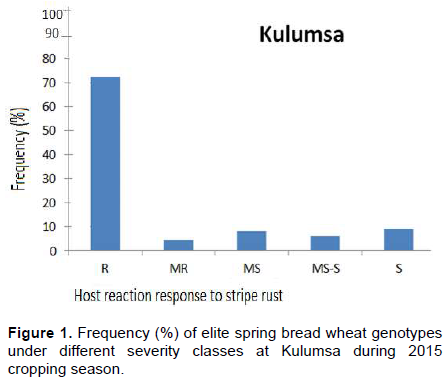
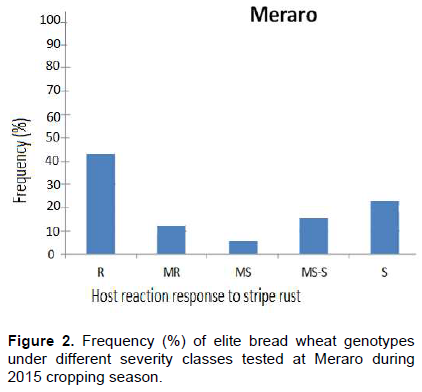
At Meraro, 86 (43%) elite genotypes exhibited resistance reaction response (CI = 0 to 20); 24 (12%) moderately resistance (CI = 20 to 30); 12 (6%) moderately susceptible (CI = 30 to 40); 30 (15%) including the five checks (Pastor-2, Kabowsh-1, Sids-1, Debira and Hiddab) were moderately susceptible to susceptible (CI = 40 to 60) and 48 (24%) were found to be susceptible (CI = 60 to 100).
After the final score, 74 genotypes (37%) out of the 200 spring bread wheat genotypes showed similar reaction response at both environments, they were resistant to stripe rust (CI from 0 to 20); 27 of these genotypes had CI less than 2 at both locations and were almost resistance to the disease. Disease severity development was increased gradually through time from 0 to 100% depending upon differences in stripe rust reaction response of the genotypes. AUDPC computed for each genotype varied from 0 to 2490 and from 0 to 1956 for Kulumsa and Meraro, respectively. The stripe rust disease development intensity through time and AUDPC at both locations are given in Appendix Table 2. Thirteen genotypes (6.5%) were susceptible to stripe rust (CI > 60) at both locations.
Generally, the AUDPC showed that the disease severity development at Meraro was higher than at Kulumsa, which indicated the availability of more virulent races, high disease pressure and/or suitable environment at Meraro than at Kulumsa.
Seedling stage screening in greenhouse
Kubsa and one mixed stripe rust isolates were used for their virulence and avirulence against the 192 elite spring bread wheat genotypes and 8 checks at seedling stage. Among them, mixed stripe rust isolates were the more virulent than Kubsa isolates. Out of the 192 spring bread wheat genotypes and 8 checks tested in the greenhouse, 53% of the genotypes showed susceptible reaction (infection type (IT)=3-4) for the mixed stripe rust isolates and 43% of the genotypes showed susceptible reaction (infection type (IT)=3-4) for Kubsa isolates. Reaction of elite spring bread wheat genotypes and checks against Kubsa and mixed isolated at seedling stage is shown in Appendix Table 2. Nearly 47% of the genotypes exhibited resistance reaction response (IT=0-2), only one genotypes showed intermediate reaction (2+) for mixed stripe rust isolates and 57% showed resistance for Kubsa isolate. Out of 192 bread wheat genotypes tested in the greenhouse, sixty-two (31%) exhibited common resistance reaction response for both (Kubsa and mixed) stripe rust isolates.
DISCUSSION
Knowledge of the genetic basis of stripe rust resistance is very essential because it will facilitate the incorporation of resistance genes into high yielding and locally adapted bread wheat cultivars and release new stripe rust resistant varieties for large scale production by end users/farmers. According to Chen et al. (2002), considerable numbers of virulent races of the stripe rust have appeared through somatic recombination or mutation.
Somatic recombination plays a major role in variation of stripe rust populations and formation of new races with combinations of previously existing virulence. Ayele et al. (1990) also reported that stripe rust isolates with virulence factors on Yr8 and Yr9 were detected in Ethiopia. In Ethiopia, stripe rust often causes substantial yield loss in higher elevation (>2400 masl), however, in 2010, the disease was widespread reaching even to the lower elevations as a result of virulence to Yr27 present in the most widely grown cultivar, ‘Kubsa’. The country previously experienced yellow rust epidemics resulting in significant yield losses to farmers (Ayele, 2002). Chen (2005) reported that high humidity with cool environment and low temperature promotes stripe rust disease by favoring spore germination.
The AUDPC result also confirmed the availability of more disease severity/pressure and suitable environment for stripe rust development at Meraro than at Kulumsa (mean AUDPC of 567.9 vs 371.4). This may be attributed to variation of environmental conditions that favor the incidence, level of disease expressions and presence of more stripe rust races and greater rust pressure at Meraro. In fact, Meraro’s environment is very cool with high humidity that is suitable for stripe rust spore germination and multiplication. Several sources of durable stripe rust resistance have been reported in wheat lines from Europe, Northwest USA, and China and in cultivars released from CIMMYT. Wang et al. (2002) indicated that field resistance in the CIMMYT wheat genotypes like Pavon-76 which has been grown in Ethiopia for the last many decades remained effective under high stripe rust pressure because Pavon-76 is adult plant resistant (APR) varieties which contain three to four genes. So, developing durable/adult plant resistance varieties like Pavon-76 is key and primary goal for breeders and is the most economically effective option to manage stripe rust in developing countries. According to Tadesse et al. (2014), most of the spring bread wheat genotypes introduced to Ethiopia from CIMMYT and ICARDA possess adult plant resistance to stripe and leaf rust based on several genes with minor effects, there is significant diversity for genes that have minor to intermediate additive effects on stripe rust resistance. In the case of seedling stage test, sixty-two (31%) of the tested genotypes were resistant for both isolates (Kubsa and Mixed) (Appendix Table 2). There were more susceptible genotypes in the mixed isolate than Kubsa isolate, mostly since the mixed races would attack more genotypes than one single race; and also due to more genes being attacked by more race than single race.
CONCLUSION
In search of resistance to wheat stripe rust, 192 elite spring bread wheat genotypes along with eight checks were tested at two locations in Ethiopia to identify those with resistance to the local pathotypes of stripe rust races and in greenhouse for seedling stag test. The identified resistant genotypes can be released to end users after testing for other traits in multi-environment trials or used as parental lines for crosses with potential and adapted wheat cultivars to develop resistant varieties.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Ayele B, Stubbs RW, Van Ginkel M, Getinet G (1990). Identification of resistance genes to Puccinia striiformis in seedlings of Ethiopian and CIMMYT bread wheat varieties and lines. Netherlands Journal of Plant Pathology 96(4):199-210. |
|
|
Ayele B (2002). Breeding wheat with multiple Disease Resistance and high yield for the Ethiopian Hilands: Broadening the Genetic Basis of Yellow Rust & Tan Spot Resistance. Cuvllier Verlag Gottingen. |
|
|
Campbell CL, Madden LV (1990). Introduction to Plant Disease Epidemiology. John Wiley & Sons, New York City. |
|
|
Chen X, More M, Milus EA, Long DL, Line RF, Marshall D, Jackson L (2002). Wheat Stripe Rust Epidemics and Races of Puccinia striiformis f. sp . tritici in the United States in 2000. The American Phytapathological Society pp. 28-30. |
|
|
Chen XM (2005). Epidemiology & control of stripe rust (Puccinia striiformis f. sp. Tritici) on wheat. Canadian journal of plant pathology 27(3):314-337. |
|
|
Central Statistical Agency agricultural Sample Survey (CSA) (2020) Volume Ireport On area And Production Of Major Crops (Private Peasant Holdings, Meher Season) Addis Ababa April, 2020587statistical Bulletin, Addis Ababa, Ethiopia. |
|
|
FAOSTAT (2018) Food and agricultural data. Resistance along with grain yield. Annals of Agricultural Sciences 60(1):29-39. |
|
|
Kankwatsa P, Singh D, Thomson PC, Babiker EM, Bonman JM, Newcomb M, Park RF (2017). Characterization and genome-wide association mapping of resistance to leaf rust, stem rust and stripe rust in a geographically diverse collection of spring wheat landraces. Molecular Breeding 37:1-24. |
|
|
Khan MH, Bukhari A, Dar ZA, Rizvi SM (2013). Status & strategies in breeding for rust resistance in wheat. Agricultural Sciences 4(6):292-301. |
|
|
Mengistu M, Netsanet BH, Merkuz A (2018). Characterization of Slow Rusting Resistance Against Stem Rust (Puccinia graminis f. sp. tritici) in Selected Bread Wheat Cultivars of Ethiopia. Advances in Crop Science and Technology 6(5):398. |
|
|
Nazari K (2011). ICARDA Research to Action Strategies to reduce the emerging wheat strip rust disease. International Wheat Stripe Rust Symposium. |
|
|
Peterson RF, Champbell AB, Hannah AE (1948). a diagrammatic scale for estimating rust intensity of leaves and stem of cereals. Canadian Journal of Research 26(5):496-500. |
|
|
Reema R, Rajender S, Neelam R, Yadav NR (2019). Evaluating stripe rust resistance in Indian wheat genotypes and breeding lines using molecular markers. Comptes Rendus Biologies 342(5-6):154-174. |
|
|
Semenov MA, Stratonovitch P, Alghabari F, Gooding MJ (2014). Adapting wheat in Europe for climate change. Journal of Cereal Science 59(3):245-256 |
|
|
Singh RP, Nelson JC, Sorrells ME (2000). Mapping Yr28 and other genes for resistance to stripe rust in wheat. Crop Science 40(4):1148-1155. |
|
|
Solh M, Nazari, Tadesse KW, CR Wellings (2012). Growing threat of stripe rust worldwide. Borlaug Global Rust Initiative (BGRI) conference. Beijing, China. |
|
|
Stubbs RW (1986). Stripe Rust: The Cereal Rusts II: Diseases, Distribution, Epidemiology, & Control. In: A.P. Roelfs & W.R. Bushnell (eds.). Academic Press, Inc.,New York, pp. 61-101. |
|
|
Tadesse W, Ogbonnaya FC, Jighly A, Nazari K, Rajaram S, Baum M (2014). Association Mapping of Resistance to Yellow Rust in Winter Wheat Cultivars and Elite Genotypes. Crop Science 54(2):607-616. |
|
|
Teklay A, Getaneh W, Woubit D, Adhiena M, Yemane N (2013). Distribution and Physiologic Races of Wheat Stem Rust in Tigray, Ethiopia. Journal of Plant Pathology and Microbiology 3(6):2-5. |
|
|
Uauy C, Brevis JC, Chen X, Khan I, Jackson L, Chicaiza O, Distelfeld A, Fahima T, Dubcovsky J (2005). High-temperature adult-plant (HTAP) stripe rust resistance gene Yr36 from Triticum turgidum ssp. dicoccoides is closely linked to the grain protein content locus Gpc-B1. Theoretical and Applied Genetics 112(1):97-105. |
|
|
Wang LF, Ma JX, Zhou RH, Wang XM, Jia JZ (2002). Molecular tagging of the yellow rust resistance gene Yr10 in common wheat, P.I. 178383 (Triticum aestivum L.). Euphytica 124(1):71-73. |
|
|
Yuan FP, Zeng QD, Wu JH, Wang QL, Yang ZJ, Liang BP, Kang ZS, Chen XH, Han DJ (2018). QTL mapping and validation of adult plant resistance to stripe rust in chinese wheat landrace Humai 15. Frontiers in Plant Science 9:968. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


