Full Length Research Paper

ABSTRACT
The sweet potato weevil, Cylas formicarius (Fabricius) (Coleoptera: Brentidae), is a destructive insect pest that damages sweet potatoes both in the field and during storage. To identify new environmentally friendly insecticides to control this insect pest, three assays (olfactory test, anti-feeding assay, and toxicity assay) were conducted to evaluate the efficacy and mode of action of 10 botanical insecticides against C. formicarius adults in 2015 and 2016. Of these 10 botanical insecticides, tea saponin, pyrethrins, and veratrine showed significant repellency in olfactory tests. Eight botanical insecticides showed anti-feeding effects in the feeding choice test. Five botanical insecticides had high toxicity. Among them, the lethal concentrations of rotenone were lowest followed by pyrethrins. The lethal time values of rotenone were shortest followed by nicotine. In conclusion, rotenone, pyrethrins, nicotine, and toosendanin have the potential to control C. formicarius adults. Of these, pyrethrins and toosendanin are more environmentally friendly than rotenone and nicotine and were identified as better insecticides to control C. formicarius.
Key words: Toxicity, repellency, anti-feeding, Coleoptera, Brentidae, environmentally friendly insecticides.
INTRODUCTION
The sweet potato weevil, Cylas formicarius (Fabricius) (Coleoptera: Brentidae), is an important insect pest of the sweet potato, Ipomoea batatas (L.) Poir.. C. formicarius attacks sweet potatoes both in the field and during storage, thus causing significant damage and yield losses between 3 and 80% (Kandori et al., 2006). Adult C. formicarius damage the vines, crowns, petioles, and storage roots of sweet potatoes, and the females lay their eggs under the epidermis of older portions of vines and storage roots. The larvae then dug tunnels into the vines and storage roots, into which they excrete feces (Reddy et al., 2014). Feeding injuries disrupt the translocation of water and nutrients in plants and induce storage roots to produce terpenoids and phenolic compounds, which causes storage roots to develop a bitter taste, and unpleasant smell, ultimately making them inedible (Akazawa and Uritani, 1960).
Traditionally, chemical insecticides are employed to control C. formicarius (Zhang et al., 2013; Wang et al., 2018). However, the concealed feeding habits of the weevil decreased the effectiveness of chemical insecticides. The improper use of chemical pesticides also causes “3R” problems (Resistance, resurgence, and residue). Although at present, resistance to a specific insecticide by weevils has not been reported, many other pests, including Euschistus heros, Laodelphax striatellus, and Frankliniella occidentalis, have reportedly developed resistance to insecticides used to control weevils (Castellanos et al., 2019; Zhang et al., 2019; Wang et al., 2012). The difficulties in controlling C. formicarius by chemical insecticides may be partly because insecticides tend to induce insect resistance. Sexual attractants, such as (Z)-3-dodecen-1-YL (E)-2-butenoate (Dilipkumar et al., 2019; Reddy et al., 2012; Smit et al., 2001) and microbial pesticides, such as Beauveria bassiana and Metarhizium brunneum (Reddy et al., 2014), and nematodes (Mannion and Jansson, 1992), also exert a controlling effect on C. formicarius populations and have been evaluated as alternatives for chemical insecticides (Yu et al., 2010). However, they work slowly and cannot prevent damage in time (Dotaona et al., 2017).
Botanical insecticides are typically extracted from poisonous plants. Their active ingredients are the secondary metabolites produced by the plant secondary metabolic pathway. These can effectively control a wide range of insect pests, including Spodoptera littoralis (Brem et al., 2002), Xanthogaleruca luteola, Ostrinia nubilalis (Jaoko et al., 2020), and Coptotermes formosanus (Mao and Henderson, 2007). Botanical insecticides quickly affect pests, degrade rapidly, are environmentally friendly and target-speciï¬c (Singh and Kaur, 2018; Kamaraj et al., 2018). They are considered as better alternatives to chemical insecticides.
Although a number of secondary metabolites that were named botanical pesticides have shown a controlling effect on Cylas spp. (Leng and Reddy, 2012; McNeil et al., 2012; Junor et al., 2008; Williams et al., 2003; Nta et al., 2018; Pan et al., 2019), their efficacy and mode of action have not been compared. Therefore the botanical insecticide best for controlling C. formicarius cannot be determined.
In this study, the efficacy and mode of action of 10 botanical insecticides with high insecticidal effect and wide insecticidal range against C. formicarius adults were evaluated. These insecticides were 25% pyrethrins, 50% rotenone, 6% celangulin, 1% stemonine, 98% matrine, 99% nicotine, 70% cnidium lactone, 1% veratrine, 98% tea saponin, and 2% toosendanin. The findings of this study provide basic data for the development of a powerful botanical insecticide to control C. formicarius.
MATERIALS AND METHODS
Insects, host plant material and botanical insecticides
Damaged sweet potato storage roots, containing C. formicarius larvae and pupae, were collected from Guangxi University Farm (Nanning, China) in December 2014 and were placed in netting bags (30 cm × 60 cm; mesh diameter 0.25 mm). Then, they were transferred to an insect rearing room at the College of Agriculture and were raised at 24 ± 2°C, 70-80% relative humidity, and a photoperiod of 14:10 h (L:D). When adult weevils appeared, additional sweet potatoes were added to the netting bags to enable reproduction and subsequent larval development. The second-generation adult weevils (5-8 days after emergence) were collected for experiments. The sweet potato variety ‘Jidanhuang’ was used in the experiment. The storage roots of sweet potatoes were washed with tap water to feed weevils. The washed storage roots were cut into strips (2 cm × 3 cm × 7 cm) for bioassay.
The following ten botanical insecticides were purchased from Shaanxi Angsheng Biomedical Technology Co. (Xi’an, China): pyrethrins (25%, from Tanacetum cinerariifolium), rotenone (50%, from Derris trifoliata), celangulin (6%, from Celastrus angulatus), stemonine (1%, from Stemona japonica), matrine (98%, from Sophora flavescens), nicotine (99%, from Nicotiana tabacum), cnidium lactone (70%, from Cnidium monnier), veratrine (1%, from Veratrum nigrum), toosendanin (2%, from Melia azedarach), and tea saponin (98%, from Camellia sinensis). High-performance liquid chromatography was used to determine the content of active ingredients of these botanical insecticides.
Olfactory test
A Y-tube olfactometer was used for olfactory tests to determine the repellency effect of different botanical insecticides on C. formicarius adults. The two arms of the Y-tube olfactometer (two arms and one stem, 15 cm in length, each at a 120° angle, and with an inner diameter of 2.0 cm) were connected to odor bottles. One of the odor bottles contained the botanical insecticide. An air pump (VLC6503-24V with a speed governor TAA02-24V, Chengdu Qihai Electromechanical Manufacturing Co. LTD, Chengdu, China) was connected to the stem of the Y-tube, and was used to draw charcoal-purified clean air from the odor bottle to the Y-tube. Airflow through each of the Y-tube arms was set to 200 ml·min-1 (combined flow 400 ml·min-1) (Yan and Wang, 2006). All olfactory tests were undertaken in complete darkness (the olfactometer was covered with a black cloth).
One of the botanical insecticides (0.1 g) was added into one of the odor bottles. The other odor bottle was left empty and was used as control. Groups of 10 adult weevils that were starved for 3-4 h were introduced into the Y-tube via the entrance of the stem. The air pump was activated and continued to pump for 5 min. After 5 min, the number of adult weevils in each arm was recorded. Weevils that did not enter any arm were not included in the analysis. Each weevil was tested only once and then discarded. When each botanical insecticide had been tested, the olfactometer was cleaned with neutral soap and 99% acetone and dried in an oven (60°C) for at least 2 h. Six replicates were applied for each insecticide.
Antifeedant assay
The two-choice test was used to determine the antifeedant activity of each botanical insecticide. The active ingredients of the botanical insecticides were prepared into four concentrations (0.25, 0.5, 1, and 2 g·L-1) with deionized water + Tween 80 (0.5%). Sweet potato strips that had been used for the bioassay were completely immersed in one of the prepared dilutions of the botanical insecticides for 5 to 8 s as treatments or were immersed in deionized water + Tween 80 (0.5%) for 5-8 s as control. All treatments and control strips were removed and air-dried for 30 min with a fan at 24 ± 2°C. Then, one treatment strip and one control strip were transferred into both ends of a horizontal transparent plastic cylinder (60 cm in length and 18 cm in diameter). Ten adult weevils, starved for 3-4 h, were introduced into the middle of the cylinder, and the number of feeding holes on both treatment and control sweet potato strips were recorded after 24 h. Five replicates were applied for each treatment.
Toxicity assay
The no-choice test was used to determine the toxicity of each botanical insecticide. During the first round of tests, the active ingredient of the botanical insecticides was prepared into four concentrations (0.25, 0.5, 1, and 2 g·L-1) with deionized water + Tween 80 (0.5%). Sweet potato strips were treated with the same method as for the antifeedant assays. The treated sweet potato strips were transferred into treated glass bottles (12 cm in height and 12 cm in diameter). The inner wall of these glass bottles was treated with the same diluted botanical insecticides and then air-dried. Each bottle contained one sweet potato strip, and the bottles were covered with a piece of netting cloth (16 cm × 16 cm). Control sweet potato strips were transferred into control glass bottles (the inner walls of which were treated with deionized water + Tween 80 (0.5%) and air-dried). Twenty adult weevils, which were starved for 3-4 h, were introduced into each bottle. The number of dead weevils in each bottle was recorded after 24 and 72 h. Three replicates were applied for each concentration of each botanical insecticide.
According to the results of the first round of tests, insecticides with high toxicity were chosen. Different concentrations of active ingredients were prepared to identify the concentration range that induced a mortality rate between zero and 100%. These included 25% pyrethrins, 50% rotenone, 2% toosendanin, 99% nicotine, and 6% celangulin. The used experimental method was the same as that of the first round of tests. After 24 and 72 h of treatment, the number of surviving weevils in each bottle was recorded. Three replicates were applied for each concentration of each botanical insecticide. Then, five insecticides were prepared at a single concentration, and the number of surviving weevils was recorded every 2 days after treatment. This identified the time range that induced a mortality rate between zero and 100%. The method was the same as the first round of tests. Three replicates were applied for each botanical insecticide. All bioassays were performed in a bioassay laboratory at 24 ± 2°C, 70-80% relative humidity, and a photoperiod of 14:10 h (L:D).
Statistical analysis
The repellency rate was calculated using the following equation:
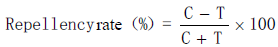
where T represents the number of C. formicarius adults in the Y-tube treatment arm, and C represents the number of C. formicarius adults in the control arm. Independent sample t-test was used to analyze the significance between the number of C. formicarius adults in the treatment arm and that in the control arm.
The antifeedant rate was calculated in the same way as the repellency rate. However, here, T represents the number of feeding holes of C. formicarius adults on the treated sweet potato strip, and C represents the number of feeding holes on the control sweet potato strip. Independent sample t-test was used to analyze the differences between the feeding holes of C. formicarius adults on the treatment strip and the feeding holes on the control strip. Tukey’s multiple comparison test was used to assess the differences in antifeedant rates of different botanical insecticides and different concentrations. Results with p < 0.05 were considered significant. The adjusted mortality for the toxicity assay was calculated using Abbott's formula (Fleming and Retnakaran, 1985) as follows:

where T represents the number of surviving adults in treatment bottles and C represents the number of surviving adults in control bottles. Tukey’s multiple comparison test was used to evaluate differences in the adjusted mortality rate between different botanical insecticides. The adjusted mortalities were arcsine-transformed prior to ANOVA.
Concentration–mortality data and time–mortality data were subjected to probit analysis to identify the lethal concentrations that cause 50 and 90% mortality (LC50 and LC90), the time required to cause 50 and 90% mortality (LT50 and LT90), their 95% fiducial limits, and chi-square values. The data were analyzed using SPSS version 22.0 (IBM Corp., Armonk, NY, USA) for Windows 10.
RESULTS AND DISCUSSION
According to the olfactory test, tea saponin (T = 5.477, df = 10, p < 0.0001), pyrethrins (T = 4.635, df = 10, p = 0.001), and veratrine (T = 2.997, df = 10, p = 0.013) exhibited a significant repellency effect against weevils. Cnidium lactone (T = 2.236, df = 10, p = 0.089) and celangulin (T = 0.649, df = 10, p = 0.552) had a repellency effect, but they did not exhibit a significant difference compared with control. Toosendanin, matrine, stemonine, nicotine, and rotenone had no repellency effect. The repellence effects of different botanical insecticides were significantly different (F = 3.153, df = 9, 50, p = 0.004) (Figure 1). The results of the olfactory test showed that the olfactory organ of the weevil plays a role in insecticide recognition. Moreover, the protective effect of several botanical insecticides on plants may be partly associated with the odor of the insecticides.

In the antifeedant assay of the 10 tested botanical insecticides, pyrethrins exhibited the strongest antifeedant activity, which was followed by toosendanin. Veratrine, stemonine, matrine, rotenone, tea saponin, and nicotine also exhibited antifeedant activity, but only at high concentrations. Cnidium lactone and celangulin had no antifeedant activity (Table 1). These results showed that antifeedant activity was very common in the tested botanical insecticides

In the toxicity assay of these 10 botanical insecticides, five botanical insecticides showed high toxicity and rotenone exhibited the highest toxicity. At 72 h after treatment, the adjusted mortality of rotenone at concentrations of 1 and 2 g·L-1 reached 100%. Pyrethrins showed the second-highest toxicity with an adjusted mortality of 78.33% at a concentration of 2 g·L-1. Toosendanin and celangulin showed moderate toxicity. Cnidium lactone, matrine, tea saponin, and stemonine showed weak toxicity. The adjusted mortality was only 3.33-8.33% at 2 g·L-1 after 72 h (Figure 2). Different lowercased letters above the bar indicate signiï¬cant differences in different botanical insecticides according to one-way ANOVA (p < 0.05).

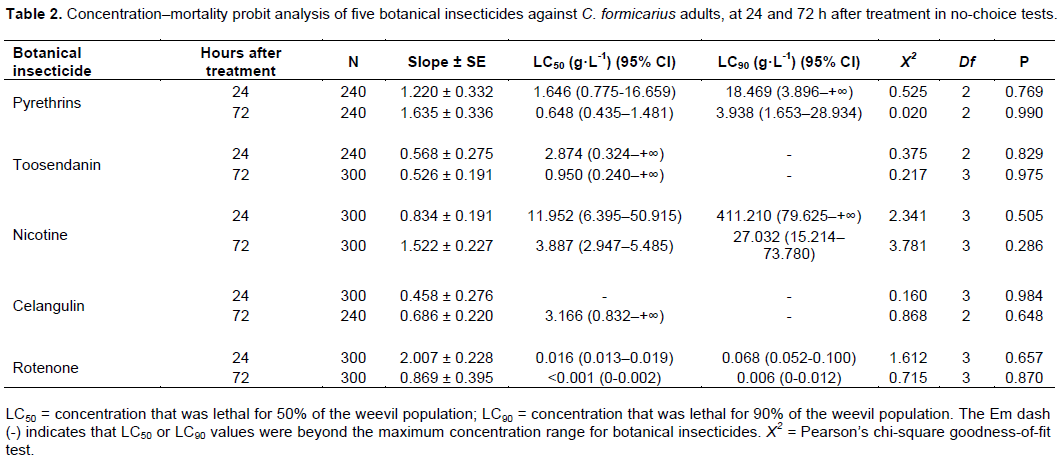
Concentration–mortality probit analysis was performed on five botanical insecticides with high mortality. The results showed that rotenone had the highest toxicity. The LC50 values of rotenone were 0.016 and <0.001 g·L-1 at 24 and 72 h after treatment, respectively, which was followed by pyrethrins and toosendanin. The LC50 value of celangulin exceeded the concentration of the raw insecticide 24 h after treatment. The LC90 values of rotenone were 0.068 and 0.006 g·L-1 after 24 and 72 h, respectively, which was followed by pyrethrins and nicotine. The LC90 values of azadirachtin and celangulin exceeded the concentration of their raw pesticides (Table 2). Time-mortality probit analyses showed that the LT50 value of rotenone was shortest, followed by that of nicotine, while that of celangulin was the longest. The LT90 value of rotenone was the shortest, followed by nicotine. The LT90 value of celangulin exceeded 50 days (Table 3).

According to the results, rotenone showed no repelling activity, a certain degree of antifeedant activity, but the strongest toxicity. At the same time, a preliminary experiment showed its moderate fumigation activity (the fumigation mortality rate of 0.1 g of raw insecticide after 24 h of treatment was 66.67%, unpublished data). Several studies demonstrated the high toxicity of rotenone for pests, mites, and nematodes (Li et al., 2017; Wen et al., 2016) as well as moderate antifeedant activity (Hu et al., 2005). However, the results of others suggest that rotenone can induce Parkinson’s disease in humans (Bu et al., 2019; Bandookwala et al., 2019). Nicotine also had no repellence activity, but exhibited moderate antifeedant activity (Rimal and Lee, 2019), high insecticidal activity (Wang et al., 2016), and moderate fumigation activity (the fumigation mortality rate of 0.1 g of raw insecticide after 24 h was 63.89%, unpublished data). However, this botanical insecticide is quickly absorbed into the mammalian bloodstream. From the bloodstream, it is easily transferred to breast milk, causing emphysema, heart rate variability, and histopathological changes in the lung and liver of suckling infants (Kobayashi et al., 2020). Consequently, the application of rotenone and nicotine as insecticides for the control of C. formicarius has potential ecological risks (Walia et al., 2017).
Pyrethrins showed a moderate repellence activity (repellence rate 44.83%), the strongest antifeedant activity (antifeedant rate 97.92% at a concentration of 2 g·L-1), and high toxicity (adjusted mortality rate 78.33% after 72 h of treatment at a concentration of 2 g·L-1). Furthermore, pyrethrins had a high fumigation activity (the mortality rate in response to 0.1 g of raw insecticide was 91.67% after 24 h of treatment, unpublished data). The strong antifeedant activity and high toxicity of pyrethrins have been confirmed before (Prota et al., 2014; Paramesha et al., 2018). Moreover, it has been shown that pyrethrin has no significant negative impact on non-target organisms (Papanikolaou et al., 2018). Toosendanin had no repellence activity, but strong antifeedant activity with an anti-feeding rate of 97.87% at a concentration of 2 g·L-1 (Jaoko et al., 2020) and moderate toxicity (with an adjusted mortality rate of 56.67% after 72 h of treatment at a concentration of 2 g·L-1) (Ma et al., 2013). The preliminary experiment also identified its high fumigation activity (the mortality rate of 0.1 g of raw insecticide was 92.0% after 24 h of treatment, unpublished data). Toosendanin is an environmentally safe insecticide (Zhang et al., 2007). Therefore, pyrethrins and toosendanin were considered the best environmentally friendly bioinsecticides of all tested insecticides for the control of C. formicarius.
Veratrine is an activator of sodium channels in saliva, and induces vomiting (Andrews et al., 1998). The antifeedant activity of veratrine against weevils as identified by this study (antifeedant rate 59.43% at 2 g·L-1) and against Helicoverpa armigera as reported by Tian et al. (2018) may be related to its emesis-inducing property. Veratrine also showed moderate repellence activity (repellence rate of 30.43%) and weak toxicity (adjusted mortality rate of 20% at 2 g·L-1). Moreover, veratrine exhibited strong fumigation activity (the fumigation mortality of 0.1 g of the raw insecticide was 96.77% at 24 h after treatment, unpublished data), indicating good application prospect for the protection of stored roots indoors.
Stemonine (Brem et al., 2002) and matrine (Mao and Henderson, 2007) exert an antifeedant effect and have weak toxicity for weevils, but showed no repellence and fumigation effect. Celangulin did not show repellence and antifeedant effects at all. However, it exhibited a weak fumigation effect and had moderate toxicity, which is lower than that of pyrethrin and azadirachtin, and its control speed is also relatively slow (Zhang et al., 2011).
Several studies have investigated the insecticidal activity of tea saponin (Lin et al., 2018; Rizwan-Ul-Hao et al., 2009; Zhu et al., 2019). In the present study, tea saponin had the strongest repellence and moderate antifeedant activity, but its acute toxicity is insufficient to control weevil in time. At the same time, tea saponin is also an exploitable surfactant (Zhu et al., 2019), which can be added to an insecticide formulation to work as a surfactant, repellent, and antifeedant.
CONCLUSIONS
The results of olfactory tests showed that tea saponin, pyrethrins, and veratrine had significant repellence effects against weevils. Antifeedant assay showed that pyrethrins and toosendanin had strong antifeedant activity. Toxicity assay showed that five botanical insecticides had high toxicity. Rotenone had the highest toxicity with the lowest LC50 values and LC90 values, followed by pyrethrins, toosendanin and nicotine. Time–mortality probit analysis showed that the LT50 value and the LT90 value of rotenone were shortest followed by nicotine.
Rotenone showed no repellence activity, a certain degree of antifeedant activity, and the strongest toxicity activity. Preliminary experiments indicated its moderate fumigation activity. Nicotine showed no repellence activity, but exhibited a moderate antifeedant activity, toxicity activity, and fumigation activity. However, application of rotenone and nicotine as insecticides for the control of C. formicarius induces potential ecological risks. Pyrethrins showed a moderate repellence activity, the strongest antifeedant activity, and high toxicity. Furthermore, its high fumigation activity was high. Toosendanin showed no repellence activity, but strong antifeedant activity and moderate toxicity. A preliminary experiment also identified its high fumigation activity. Neither of the two botanical insecticides had significant negative impact on non-target organisms. Therefore, pyrethrins and toosendanin can be considered the best environmentally friendly bioinsecticides for the control of C. formicarius. Veratrine showed strong fumigation activity and moderate anti-feeding activity, indicating its application prospect for the protection of roots stored indoors. Although tea saponin is less toxic to weevils, it had the strongest repellent ability, moderate anti-feeding activity and good surfactant activity, suggesting its merit as surfactant that can be added to insecticide preparations.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Akazawa T, Uritani I (1960). Isolation of ipomeamarone and two coumarin derivatives from sweet potato roots injured by the weevil, Cylas formicarius elegantulus. Archives of Biochemistryand Biophysics 88:150-156. |
|
|
Andrews PLR, Sims DW, Young JZ (1998). Induction of emesis by the sodium channel activator veratrine in the lesser spotted dogfish, Scyliorhinus canicula (Chondrichthyes: Elasmobranchii). Journal of the Marine Biological Association of the United Kingdom 78:1269-1279. |
|
|
Bandookwala M, Sahu AK, Thakkar D, Sharma M, Khairnar A, Sengupta P (2019). Edaravone-caffeine combination for the effective management of rotenone induced Parkinson's disease in rats: An evidence based affirmative from a comparative analysis of behavior and biomarker expression. Neuroscience letters 711:1-6. |
|
|
Brem B, Seger C, Pacher T, Hofer O, Vajrodaya S, Greger H (2002). Feeding deterrence and contact toxicity of Stemona alkaloids - A source of potent natural insecticides. Journal of Agricultural and Food Chemistry 50(22):6383-6388. |
|
|
Bu J, Qiao X, He Y (2019). Colonic electrical stimulation improves colonic transit in rotenone-induced Parkinson's disease model through affecting enteric neurons. Life Science 231:1-10. |
|
|
Castellanos NL, Haddi K, Carvalho GA, de Paulo PD, Hirose E, Guedes RNC, Smagghe G, Oliveira EE (2019). Imidacloprid resistance in the Neotropical brown stink bug Euschistus heros: selection and fitness costs. Journal of Pest Science 92(2):847-860. |
|
|
Dilipkumar M, Erwan-Shah S, Anuar A, Sivapragasam A (2019). A sex pheromone-baited trapping system for management of sweetpotato weevil, Cylas formicarius (Coleoptera: Brentidae). Journal of Applied Entomology 143(4):408-416. |
|
|
Dotaona R, Wilson BAL, Stevens MM, Holloway J, Ash GJ (2017). Chronic effects and horizontal transmission of Metarhizium anisopliae strain QS155 infection in the sweet potato weevil, Cylas formicarius (Coleoptera: Brentidae). Biolgical Control 114:24-29. |
|
|
Fleming R, Retnakaran A (1985). Evaluating Single Treatment Data Using Abbott's Formula With Reference to Insecticides. Journal of Economic Entomology 78(6):1179. |
|
|
Hu MY, Zhong GH, Sun ZT, Sh G, Liu HM, Liu XQ (2005). Insecticidal activities of secondary metabolites of endophytic Pencillium sp. in Derris elliptica Benth. Journal of Applied Entomology 129(8):413-417. |
|
|
Jaoko V, Nji TTC, Backx S, Mulatya J, Van den Abeele J, Magomere T, Olubayo F, Mangelinckx S, Werbrouck SPO, Smagghe G (2020). The phytochemical composition of Melia volkensii and its potential for insect pest management. Plants (Basel, Switzerland) 9 (2):1-11. |
|
|
Junor GO, Porter RBR, Yee TH, Williams LAD (2008). Chemical composition and insecticidal activity of the essential oils from Bursera hollickii (Britton) found in Jamaica. Journal of Essential Oil Research 20(6):560-565. |
|
|
Kamaraj C, Gandhi PR, Elango G, Karthi S, Chung I, Rajakumar G (2018). Novel and environmental friendly approach: Impact of neem (Azadirachta indica) gum nano formulation (NGNF) on Helicoverpa armigera (Hub.) and Spodoptera litura (Fab.). International Journal of Biological Macromolecules: A 107:59-69. |
|
|
Kandori I, Kimura T, Tsumuki H, Sugimoto T (2006). Cold tolerance of the sweet potato weevil, Cylas formicarius (Fabricius) (Coleoptera: Brentidae), from the Southwestern Islands of Japan. Applied Entomology and Zoology 41(2):217-226. |
|
|
Kobayashi K, Tsugami Y, Suzuki N, Suzuki T, Nishimura T (2020). Nicotine directly affects milk production in lactating mammary epithelial cells concurrently with inactivation of STAT and glucocorticoid receptor in vitro. Toxicology in vitro 63:1-8. |
|
|
Leng PH, Reddy GVP (2012). Bioactivity of Selected Eco-Friendly Pesticides against Cylas formicarius (Coleoptera: Brentidae). Florida Entomologist 95(4):1040-1047. |
|
|
Li Z, Huang R, Li W, Cheng D, Mao R, Zhang Z (2017). Addition of cinnamon oil improves toxicity of rotenone to Spodoptera litura (Lepidoptera: Noctuidae) larvae.Florida Entomologist 100(3):515-521. |
|
|
Lin S, Chen YX, Bai Y, Cai HJ, Wei H, Tian HJ, Zhao JW, Chen Y, Yang G, Gu XJ, Murugan K (2018). Effect of tea saponin-treated host Plants on activities of antioxidant enzymes in larvae of the diamondback moth Plutella xylostella (Lepidoptera: Plutellidae). Environmental Entomology 47(3):749-754. |
|
|
Ma Z, Gulia-Nuss M, Zhang X, Brown MR (2013). Effects of the botanical insecticide, toosendanin, on blood digestion and egg production by female Aedes aegypti (Diptera: Culicidae): Topical application and ingestion. Journal of Medical Entomology 50(1):112-121. |
|
|
Mannion CM, Jansson RK (1992). Movement and postinfection emergence of entomopathogenic nematodes from sweetpotato weevil, Cylas formicarius (F.) (Coleoptera: Apionidae).Biological Control 2:297-305. |
|
|
Mao L, Henderson G (2007). Antifeedant activity and acute and residual toxicity of alkaloids from Sophora flavescens (Leguminosae) against Formosan subterranean termites (Isoptera: Rhinotermitidae). Journal of Economic Entomology 100(3):866- 870. |
|
|
McNeil MJ, Porter RBR, Williams LAD (2012). Chemical composition and biological activity of the essential oil from Jamaican Cleome serrata. Natural Product communications 7(9):1231-1232. |
|
|
Nta AI, Okweche SI, Umoetok SB (2018). Efficacy of three plant powders in the control of Cylas puncticollis (Boheman) (Coleoptera: Curculionidae) on sweet potato during storage. African Entomology 26(1):141-149. |
|
|
Pan R, Tang X, He L, Ren L (2019). Effects of tea saponin on behavior, growth and development of Cylas formicarius (Fabricius) (Coleoptera: Brentidae). Southwest China Journal of Agricultural Sciences 32(7):1566-1571. |
|
|
Papanikolaou NE, Kalaitzaki A, Karamaouna F, Michaelakis A, Papadimitriou V, Dourtoglou V, Papachristos DP (2018). Nano-formulation enhances insecticidal activity of natural pyrethrins against Aphis gossypii (Hemiptera: Aphididae) and retains their harmless effect to non-target predators. Environmental Science and Pollution Research 25(11):10243-10249. |
|
|
Paramesha M, Manivannan S, Rao SJA, Srikanth KS, Neelwarne B, Shetty NP (2018). Augmentation of pyrethrins content in callus of Chrysanthemum cinerariaefolium and establishing its insecticidal activity by molecular docking of NavMS Sodium Channel Pore receptor. 3 Biotech 8(8):1-10 |
|
|
Prota N, Bouwmeester HJ, Jongsma MA (2014). Comparative antifeedant activities of polygodial and pyrethrins against whiteflies (Bemisia tabaci) and aphids (Myzus persicae). Pest Management Science 70(4):682-688. |
|
|
Reddy GVP, Gadi N, Taianao AJ (2012). Efficient sex pheromone trapping: Catching the sweetpotato weevil, Cylas formicarius. Journal of Chemical Ecology 38 (7):846-853. |
|
|
Reddy GVP, Zhao Z, Humber RA (2014). Laboratory and ï¬eld efï¬cacy of entomopathogenic fungi for the management of the sweet potato weevil, Cylas formicarius (Coleoptera: Brentidae). Journal of Invertebrate Pathology 122:10-15. |
|
|
Rimal S, Lee Y (2019). Molecular sensor of nicotine in taste of Drosophila melanogaster. Insect Biochemistry and Molecular Biology 111:1-7. |
|
|
Rizwan-Ul-Hao M, Hu QB, Hu MY, Zhong G, Weng Q (2009). Study of destruxin B and tea saponin, their interaction and synergism activities with Bacillus thuringiensis kurstaki against Spodoptera exigua (Hübner) (Lepidoptera: Noctuidae). Applied Entomology and Zoology 44(3):419-428. |
|
|
Singh B, Kaur A (2018). Control of insect pests in crop plants and stored food grains using plant saponins: A review. LWT - LWT - Food Science and Technology 87:93-101. |
|
|
Smit NEJM, Downham MCA, Laboke PO, Hall DR, Odongo B (2001). Mass-trapping male Cylas spp. with sex pheromones: a potential IPM component in sweetpotato production in Uganda. Crop Protection 20(8):643-651. |
|
|
Tian W, Li Q, Yan H, Yang S (2018). Bioactivity of ethanol extracts from Acorus calamus var. angustatus to eggs and larvae of Helicoverpa armigera. Acta Phytophylacica Sinica 45(5):1012-1020. |
|
|
Walia S, Saha S, Tripathi V, Sharma KK (2017). Phytochemical biopesticides: some recent developments. Phytochemistry Reviews 16:989-1007. |
|
|
Wang C, Yin F, Jia X, Chen X, Chen S (2018). Comparative efficacy tests of 6 pesticides to control sweet potato weevil. Pesticide Science and Administration 39(8):45-49. |
|
|
Wang J, Luan Q, Jin R, Zhang Y, Jiang X, Yun L, Wang Q, Zhang T (2016). Toxicity and Field Control Efficacy of 6 Low-toxic Insecticides to Dictyoploca japonica. Journal of Northwest Forestry University 31(3):179-182. |
|
|
Wang SY, Zhou XH, Zhang AS, Li LL, Men XY, Zhang SC, Liu YJ, Yu Y (2012). Resistance mechanisms and cross-resistance of phoxim-resistant Frankliniella occidentalis Pergande population. The Journal of Applied Ecology 23(7):1933-1939. |
|
|
Wen RR, Ma L, Dong WY, Wang BY (2016). Toxic effect of rotenone on Gypsy moth. Journal of Northeast Forestry University (9):22. |
|
|
Williams LAD, Vasques E, Reid W, Porter R, Kraus W (2003). Biological activities of an extract from Cleome viscosa L. (Capparaceae). Naturwissenschaften 90(10):468-472. |
|
|
Yan ZG, Wang CZ (2006). Identification of Mythmna separata-induced maize volatile synomones that attract the parasitoid Campoletis chlorideae. Journal of Applied Entomology 130(4):213-219. |
|
|
Yu HB, Zheng Q, Chen SL (2010). Biological characteristics and integrated control measures of Cylas formicarius. Journal of Heibei Agricultural Science 14:32-35. |
|
|
Zhang H, Wang LP, Fan KJ, Zou XM, Chen SL, Wang RY (2013). Field screening test of sweet potato weevils in the Sanxia Reservoir area. Shaanxi Journal of Agricultural Sciences 59(06):34-36. |
|
|
Zhang JW, Hu Z, Li SK, Wu WJ (2011). Synthesis and insecticidal activities of new ester-derivatives of celangulin-V. International Journal of Molecular Sciences 12(12):9596-9604. |
|
|
Zhang Y, Ma Z, Wang H, Wang Z, Gao B, Feng J, Zhang X (2007). Evaluation of the toxicity of botanical pesticide toosendanin to non-target organisms. Acta Scientiae Circumstantiae 27(12):2038-2045. |
|
|
Zhang Y, Ma X, Han Y, Wang L, Liu Z, Guo H, Fang J (2019). Transcript-level analysis of detoxification gene mutation-mediated chlorpyrifos resistance in Laodelphax striatellus (Hemiptera: Delphacidae). Journal of Economic Entomology 112(3):1285-1291. |
|
|
Zhu Z, Wen Y, Yi J, Cao Y, Liu F, McClements DJ (2019). Comparison of natural and synthetic surfactants at forming and stabilizing nanoemulsions: Tea saponin, Quillaja saponin, and Tween 80. Journal of Colloid and Interface Science 536:80-87. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


