ABSTRACT
Nitrogen-15 isotopic dilution method was used to estimate biological nitrogen fixation (BNF) and nitrogen (N) budget of fourteen (14) soybean varieties using maize as reference crop. The experiment was carried out at Sekou in Southern Benin. The amount of N derived from air (Ndfa kg N ha-1) ranged from 51 for variety BRS 261 to 148 for variety Canarana. In a scenario where the soybean shoot dry matter and grains were removed from the field after harvest and only the fallen leaves were incorporated into the soil, the N budget ranged between -91 (Canarana) and -17 kg N ha-1 (BRS 260). When only soybean grain was exported from the fields and fallen leaves and shoot dry matter are incorporated into the soil, the N budget varied from 7 (BRS 261) to 74 kg N ha-1 (BRS Garantia). The study showed that Canarana, TGX 1448 2E and BRS Paraiso soybean varieties fixed the highest amount of N among the 14 varieties. The inclusion of those soybean varieties in cereal-based cropping systems would help reduce N inputs and improve soil and crop productivity in farming systems in Benin.
Key words: 15N enrichment technique, N2 fixation, N balance, soil fertility, Benin.
In tropical regions, nitrogen (N) deficiency is frequently one of the major factors limiting crop yields and N fertilization is increasingly applied to increase the crop yields (Reinhold-Hurek and Hurek, 2003). Managing nitrogen inputs in crop production system to achieve economic and environmental sustainability is a major challenge facing agriculture (Haque and Sattar, 2010). In this context, biological fixation (BNF) has become one of the most attractive strategies for the development of sustainable agricultural systems (Hayat et al., 2008). However, farmers in West Africa are often reticent to adopt legume cover crops such as Mucuna species that are not used for human consumption or without a direct economic profit, in spite of the positive impact on restoring soil fertility (Mayong et al., 1999). Several authors proved interests to use dual-purpose grain legumes in farming systems (Sanginga et al., 2003; Houngnandan et al., 2009). The integration of grain legumes such as dual-purpose soybean [Glycine max (L.) Merril] into rice or maize-based systems has been reported to greatly enhance rice and maize productivity and the sustainability of the production systems in the West African savannas (Houngnandan et al., 2009). Indeed, soybean is the most important cropped legume in the world due to its high protein content, its lower susceptibility to pests and diseases, better grain storage quality, huge leaf biomass yield, high ability to fix nitrogen in association with Bradyrhizobium in the root nodules and it N contribution to subsequent crops (Mafongoya et al., 2009). Since, interest in growing soybean has increased in Benin, the exploitation of soybean could be an attractive strategy for sustainable agricultural production in highly degraded soils in the country. But to obtain such a beneï¬cial residual effect after legumes compared to non-legumes, it is expected that the amount of ï¬xed N returned by the legumes to the soil must be greater than the amount of soil N in the harvested grain (Sanginga et al., 2002; Schipanski et al., 2010). It is widely recognized that soybean crops often require more N (or export more N in grain) than they fix and this often results in a net negative contribution to the N balance in the cropping systems (Jaynes and Karlen, 2008; Salvagiotti et al., 2008). The role of BNF, especially in legumes, is well established and documented but it has been reported that various varieties or cultivars of grain legumes show significant differences regarding their ability to support BNF (Hayat et al., 2008; Singh and Shivakumar, 2010).
It has been reported that in various cultivars of grain legume, the estimation of N2 fixation depends largely on the methods used (Hardarson et al., 1993). A number of methods have been developed to quantify biological nitrogen fixation, but each method has its own merits and limitations (Herridge et al., 2008; Unkovich et al., 2008). The choice of a particular method depends on the type and site of the experiment, the available resources and the species and system in question (Gathumbi et al., 2002; Schweiger et al., 2014). The most commonly methods used to determine N fixation in legumes are: N difference, ureides, acetylene reduction assay, 15N natural abundance and 15N enrichment methods (Forrester et al., 2006; People et al., 2009). Among these methods, 15N enrichment and 15N natural abundance are considered to be time integrated methods to determine N2 fixation under natural conditions (Boddey et al., 2000; Unkovich and Pate, 2000). But it is generally believed that isotope dilution utilizing enriched 15N gives the most accurate quantification of nitrogen turnover in the main processes of the nitrogen cycle (Chalk, 1985).
This study aims to estimate the biological N2 fixation of fourteen Brazilian soybean varieties introduced in the Southern Benin by the 15N isotopic dilution method and also estimate the total net N inputs in soil system.
The study was carried out at the Application and Production Farm of the Faculty of Agronomic Sciences of the University of Abomey-Calavi located at Sekou (2°14' - 2°26 E and 6°37' - 6°40' N) in Benin from July to October, 2009. The climate is warm and subequatorial humid with a bimodal rainfall distribution. According to the weather station ASECNA (from year 1988 to year 2008), the annual mean temperature was between 26 and 29°C and the annual mean rainfall was between 1000 and 1400 mm. However, during the experiment, mean precipitation and temperature near the site were 708 mm and 27°C, respectively. The experimental farm has been established on a typical “terre de barre” soil, classified by Food and Agriculture Organization-United Nations Educational, Scientific and Cultural Organisation as Rhodic Ferralsol. Rhodic Ferralsol is a highly degraded soil due to high demographic pressure; intensive cropping with less or no financial capacity for farmers to apply chemical fertilizers, while fallow in the “Terre de Barre” area disappeared. The experimental farm soil had a sandy-clay texture, and its main chemical characteristics are presented in Table 1.
Fourteen soybean varieties were introduced from three different countries.
(1) Ten Brazilian varieties: BRS 260, BRS 261, BRS 262, BRS 268, BRS Ipameri, BRS Santacruz, BRS Luziania, BRS Paraiso, BRS Garantia, BRS Vencedora;
(2) Two varieties introduced from Ghana (Anidazo and Jenguma) and one from Côte d’Ivoire (Canarana). They were all originally from Brazil.
(3) One variety of International Institute of Tropical Agriculture (IITA): TGX 1448 2E which was used as control because it was already in the extension systems few years ago in Benin.
The microbiological material was the inoculum prepared from IRAT FA3 strain of specific Bradyrhizobial bacteria fixed on the peat. IRAT FA3 strain was selected by the Nitrogen Fixation in Tropical Agricultural Legumes/Microbial Resource Centre Rhizobium project and used to inoculate strictly nodulating soybean varieties adapted to tropical agricultural zones in Africa and Latino America (Houngnandan et al., 2008).
Experimental design
The field experiments were a randomized completed block design with three replicates or blocks. Each block contained fourteen plots. Each plot contained four planting lines. The space between rows was 50 cm and the inter-plant spacing was 20 cm. On the planting row, soybean was seeded at a 5 cm within-row space. Four seeds were sown per hole and later thinned to three 14 days after. Soybean seeds were first inoculated with IRAT FA3 strain of B. japonicum containing an approximate density of 108 viable rhizobia per seed before sowing. One maize plot was sown aside each soybean main plot and used as a reference plant for soybeans varieties. The space between rows was 75 cm and the inter-plant spacing was 40 cm. The isotopic dilution method was applied using micro plots installed in all plots (1 m width × 1 m length). The micro plots of soybean and the reference micro plots were enriched with 20 kg N/ha of 5.73% 15N atom excess labeled urea dissolved in water when the remaining plots received 20 kg N/ha of unlabelled urea solution. All plots received 100 kg ha-1 of P2O5 as triple superphosphate.
Plants sampling
Soybean and maize shoots were collected at the first sampling (at the flowering time 10 weeks after planting). In each plot, eight plants and two plants were randomly sampled, respectively for soybeans and maize. Shoots and roots of soybeans and maize were separated. Roots were washed to remove adhering soil. Nodules were removed and counted and dry weight was taken. Soybean shoots and roots and the reference plants were dried at 65°C for 72 h and ground. Nitrogen content of both plant (soybean and maize) was analyzed by the Kjeldahl method. The labeled soybean and maize sampled were ball milled, well packed and sent to the Seiberdorf laboratory (International Atomic Energy Agency) for 15N atom% excess analysis using an isotope ratio mass spectrometer.
At the second sampling (at the harvest), straws and grains were sampled on an area of 6 m² (3 m × 2 m) per plot. Total fresh weights of straws and grains harvested were taken. Then, a sub-sample of straw and grain was taken for yield calculation. Total N straw and grain was analyzed by the Kjeldahl method.
METHODOLOGY AND CALCULATIONS
Statistical analysis
Statistical analyses were carried out using SAS software version 9.2. One-way analysis of variance (ANOVA) was performed to determine the statistical differences among the soybean varieties. When significant differences (p < 0.05) were noticed, a Student-Newman-Keuls test was used to compare the means.
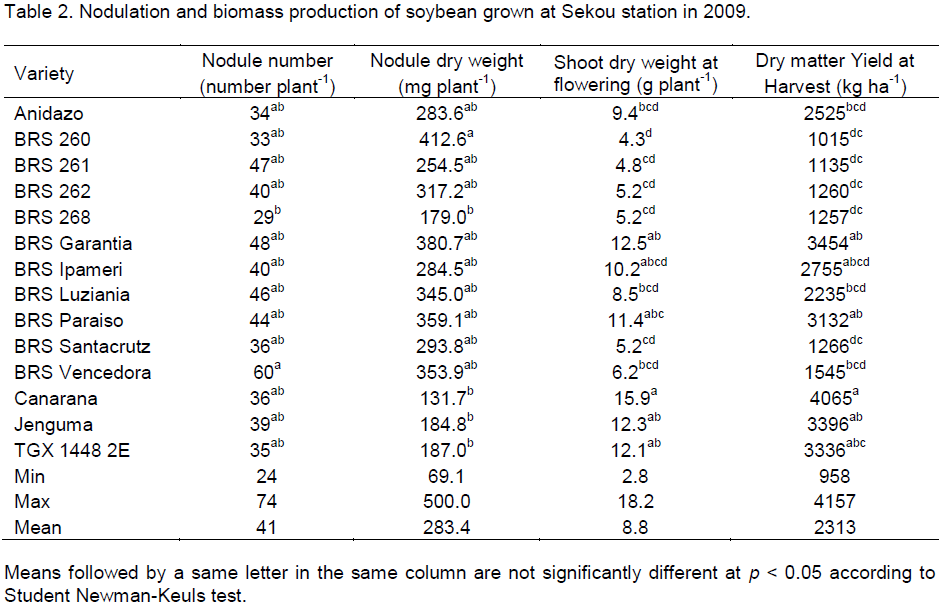
Nodulation and biomass production
Nodulation was observed on all soybean cultivars (Table 2) and differences among cultivars in nodule number and nodule dry weight were significant (p <0.05). The lowest nodule number (29) was recorded by BRS 268, but Canarana had the lowest nodule dry weight (131.7 mg plant-1). The highest nodule number and nodule dry weight were observed, respectively on BRS Vencedora (60) and BRS 260 (412.6 mg plant-1). But the control (TGX 1448 2E) had 35 nodules per plant which weighed 187 mg.
There were highly significant difference (P<0.001) among soybean varieties for biomass production (Table 2). At flowering and at maturity, Canarana showed the highest biomass production (15.9 mg plant-1 and 4065 kg ha-1) and BRS 260 had the lowest (4.3 mg plant-1 and 1015 kg ha-1). The control (variety TGX 1448 2E) produced 12.1 g of biomass per plant at flowering and 3336 kg ha-1 at harvest. The three best varieties in terms of biomass production were Canarana, BRS Garantia and Jenguma.
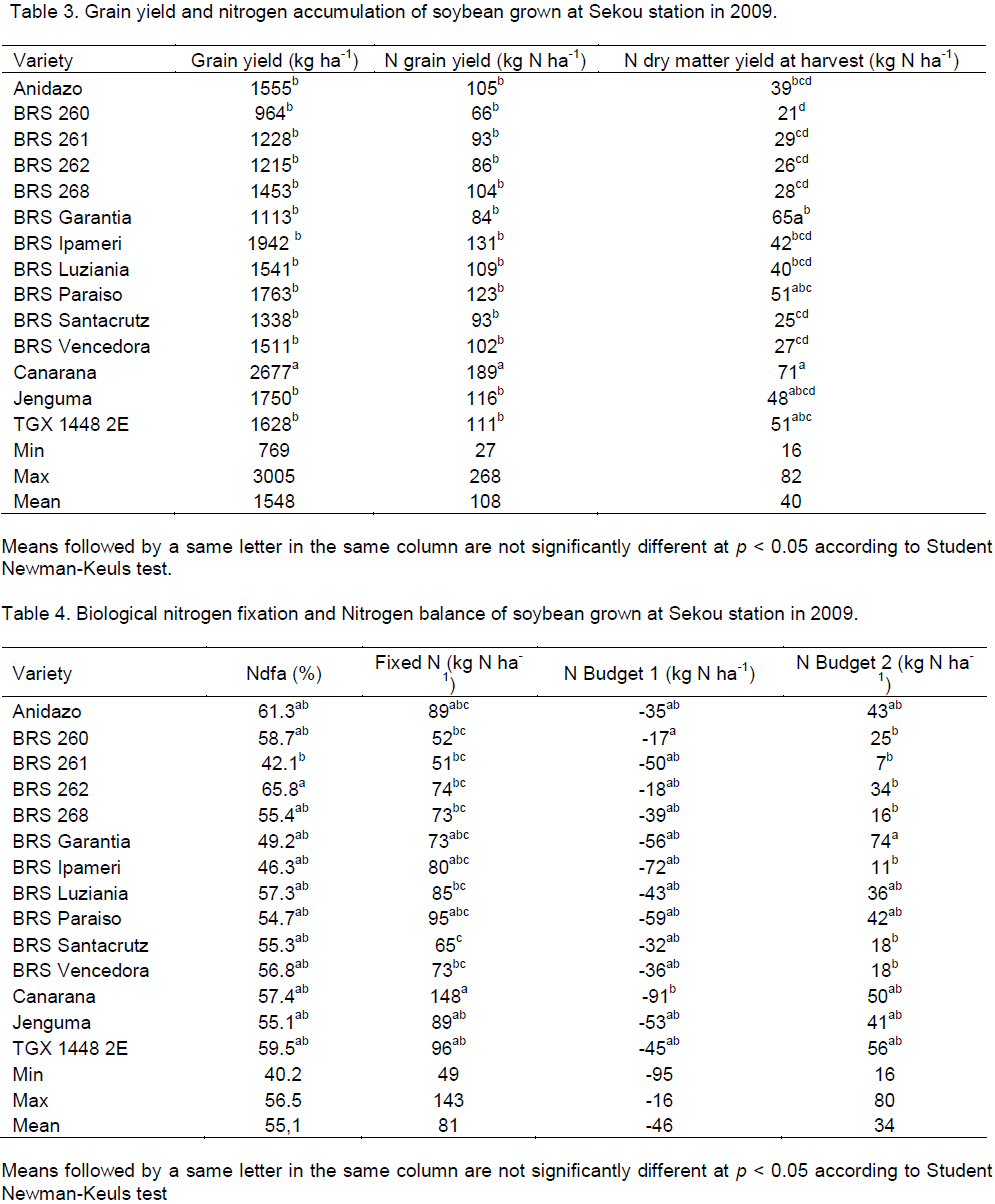
Grain yield and nitrogen accumulation
A significant difference (p<0.05) was observed on soybean varieties for grain yield and N grain yield (Table 3). The grain yield ranged between 964 (BRS 260) and 2677 kg DM ha-1 (Canarana). The N grain yield varied from 66 (BRS 260) to 189 kg N ha-1 (Canarana).
There were highly significant differences (P<0.001) among soybean varieties for total N yield (Table 3). Canarana showed the highest N yield (71 kg N ha-1) and BRS 260 had the lowest (21 kg N ha-1).
The control, TGX 1448 2E produced 1628 kg DM ha-1 grain yield. It accumulated in grains 111 and 51 kg N ha-1 in biomass. It was ranked third in terms of total N accumulation after Canarana and BRS Garantia at harvest and ranked fifth after Canarana, BRS Ipameri, BRS Paraiso and Jenguma among all varieties in terms of N grain yield.
Biological nitrogen fixation and nitrogen balance
All soybeans varieties derived nitrogen from atmosphere (Table 4) and differences among varieties in % of N derived from atmosphere (Ndfa) and the total N fixed (kg N ha-1) were significant (p<0.001). BRS 261 had the lowest %Ndfa (42.1%) and BRS 262 had the highest (65.8%). The control TGX 1448 2E showed 59.5% Ndfa. The amount of N fixed ranged between 51 (BRS 261) and 148 kg N ha-1 (Canarana). Canarana, TGX 1448 2E and BRS Paraiso showed the highest amount of N fixed (Table 4).
The N balance with all shoot dry matter removed (N Budget 1) and with all shoot dry matter incorporated (N Budget 2), indicated significant differences (p<0.01) between soybeans varieties (Table 4). N budget 1 ranged between -91 (Canarana) and -17 kg N ha-1 (BRS 260). N budget 2 varied from 7 (BRS 261) to 74 kg N ha-1 (BRS Garantia). But for the variety TGX 1448 2E grown as control, N budget 1 and N budget 2 were, respectively - 45 and 56 kg N/ha.
Correlation between amount of nitrogen fixed and selected yield parameters
Correlation between amount of nitrogen fixed and some yield parameters is shown in Table 5. N fixed was significantly and positively correlated with N grain yield, N biomass production and N budget 2, but negatively with N budget 1. No correlation was observed between N fixed and nodule number. N budget 1 and N budget 2 were, respectively negatively and positively correlated with N grain yield, N biomass production and N fixed.
Biological nitrogen fixation of the soybean varieties
These results showed that 15N isotopic dilution method can be used to evaluate the percentage and amount of N derived from atmosphere by the 14 soybean varieties. The precision and accuracy of nitrogen fixation estimated with this method depend on the choice of reference plant. A prerequisite for using the method is that the legume and reference plant should not differ in the ratio of N assimilated from added 15N to endogenous unlabelled N taken up from the soil (Stahl et al., 2002). The criteria for the selection of the appropriate reference crop are: having no ability to fix nitrogen, having the same ability to extract nitrogen and also the same relative nitrogen uptake profile as fixing plant (Unkovich et al., 2008). The best non-fixing reference crop usually is non-nodulating lines of the test legume (Okito et al., 2004). But when a non-nodulating line is not available, non-fixing reference mono or dicotyledonous crops (Reiter et al., 2002) or non-legume weeds (Schwenke et al., 1998) could be used. Since a non-nodulating line of soybean was not available during our study, maize was used as the reference crop. The use of maize to estimate N fixation with the 15N isotopic dilution method, was previously reported (Adu-Gyamfi et al., 2007). But it may be better to choose several reference crops to estimate nitrogen fixed using 15N isotopic dilution method because of variable response of the reference crops in different conditions.
In this study, percentage of Ndfa ranged between 42 and 66% and the N fixed varied from 51 to 148 kg N ha-1. Similar results were obtained by several authors for inoculated soybean cultivar. For example, using the 15N isotope dilution method on five promiscuous IITA soybean lines over two seasons at Mokwa, in the southern Guinea savannah of Nigeria, Sanginga (2002) reported mean value of 91 kg N ha−1. In other countries, soybeans have been reported to fix 85 to 154 kg N ha-1 in Brazil, 26 to 57 kg N ha-1 in Thailand, 78 kg N ha-1 in Australia (Peoples and Crasswell, 1992; Salvagiotti et al., 2008).
Peoples et al. (2009) reported a mean fixation rate for North America of 144 kg N ha-1. Nodulation was observed on all soybean varieties. These results confirm those of Sanginga et al., (2000) and Houngnandan et al., (2008). But the number of nodules was not correlated with amount of N fixed. There were varieties which had a lot of nodule number, but fixed a small amount of N contrary to varieties which had little nodule number with a high amount of N derived from atmosphere. For example, Canarana had 36 nodules and fixed 148 kg N ha-1 while BRS Vencedora having 60 nodules, fixed only 73 kg N ha-1. Thuita et al., (2012) reported that increase in nodulation was not accompanied by an increase in %Ndfa. Similar results were found by De Bruin et al., (2010) who suggested that it is pertinent to know prior to the introduction of a soybean variety if inoculation can better promote nodulation and N2 fixation.
Nitrogen balance
Nitrogen balance values were negative when shoot dry matter and grain were removed from field after harvest. These values were improved when only shoot dry matter was left in the field. Biomass production, N yield and N fixed were negatively correlated with N budget 1, but positively with N budget 2. The N export from harvested grains was for all varieties greater than the N input through symbiotic N2 fixation, resulting in a negative N balance. This means that irrespective of the cropping system, soybean resulted in a net removal of N from the soil in spite of symbiotic N2 fixation (Jaynes and Karlen, 2008; Salvagiotti et al., 2008). Larger amounts of N derived from air were accumulated by soybean; therefore, a large proportion of N fixed by soybean is exported from the field at grain harvest. Previous studies showed that soybean cultivation results in a net N loss, if all residues are removed from the field.
If residues are retained, soybean N budgets range from -35 to 50 kg N ha-1 depending on the cultivar and environmental conditions (Sanginga et al., 2002). Amanuel et al., (2000) reported that negative N balance was due to the fact that nitrogen input (fertilizer and nitrogen fixation) was not enough to meet crop demand. They also demonstrated that N balance after legume harvest is positive when crop residues are returned to soil and only seed or grain is removed. The same results were reported (Adu-Gyamfi et al., 2007) for intercrops of maize-pigeon pea. They showed that, in situations where the aboveground biomass of both the legume and the cereal were removed from the field, high negative values were observed, but if only the grains were exported, all budgets were improved. However, Schipanski et al. (2010) found that a positive N balance resulted when the percentage of N from fixation was greater than 60%. Laberge et al., (2009) found the same result on soybean, but they showed that errors can arise when important processes are ignored and the results of their study suggested that N rhizodeposition from soybean and grain legumes was such a neglected process. Rhizodeposits include roots exudates, fine roots, and root necrosis products accrued in the soil during plant growth (Hertenberger and Wanek, 2004). In reality, most plant biomass, and its N content, left in the field from the previous cropping season is lost over the dry season due to strong winds, free-roaming ruminants, bush fires and termites (Schulz et al., 2001). Ghosh et al., (2007) showed that besides the direct addition of N through the above-ground biomass, the legumes may enhance the soil available N pool for following crops via root exudates or inefficiencies in recovering soil mineral N during the legume phase, and subsequent decomposition of root and nodule residues. Birouste et al., (2012) and Arcand et al., (2013) have estimated that root-derived N comprises as much as 80% of the total ground N. These indirect additions of N contribute to substantial saving of N (20 to 30 kg N/ha) and enrichment of soil fertility even when all the legume residues are removed (Sharma, 2005).
In West Africa, the use of dual-purpose grain soybean varieties with potential for good nitrogen fixation and nitrogen balance is a promising technology that has multiple benefits such as improving household nutrition, source of cash income, and supply of N inputs, which can contribute to improve soil fertility and to the sustainability of the cropping systems since farmers are often reluctant to adopt legume cover crops that are not useful for human consumption or without a direct economic benefit (Mayong et al., 1999; Vanlauwe et al., 2001). Thus, soybean with a positive N balance can be used to replenish soil nutrients and contribute to reduce land degradation occurring in Benin and consequently improve subsequent cereals crops. Indeed, the benefits of including legumes as green manures in rotations with cereal crops as a source of nitrogen (N) is well documented (Mason and Spaner, 2005; Kirkegaard et al.,
2008; Peoples et al., 2009a). Legumes have the ability to supply a renewable source of N to agricultural soils through biological N-fixation, providing an economically and ecologically attractive means of delivering N to non-leguminous crops and reducing off-farm N inputs (Kirkegaard et al., 2008; Peoples et al., 2009; Thiessen Martens et al., 2001). However, smallholder farmers invariably carry the whole shoot of soybean from the fields for threshing. Or, if the legume stover is removed, there is often no observable benefit to the next crop and there is usually a net removal of N from the cropping system in the legume grain (Giller, 2001) and this is well reported in our results indicating negative N budget ranging from -91 to -17 kg N ha-1.
This study aimed to assess biological nitrogen fixation of fourteen soybean varieties using isotopic dilution method with maize as reference plant. With this method, it appeared that all soybean varieties tested fixed nitrogen derived from air. But a particular problem of this method is that the enrichment the 15N enrichment of the soil N would need to be relatively constant over time and space, or the time course and depth of soil N uptake by the reference and N2-fixing plants the same. The result would be more reliable if a non-fixing soybean cultivar instead of maize was used, because soybean and maize do not have similar root characteristics.
The study showed that Canarana, TGX 1448 2E and BRS Paraiso soybean varieties were identified as the highest amount of N fixed capacity among the 14 varieties. From the results, it could be concluded that the involvement of Canarana, TGX 1448 2E and BRS Paraiso soybean varieties in cereal-based cropping systems would help reduce N inputs and improve soil and crop productivity in farming systems in Benin. The incor-poration of legume residues into the soil immediately after harvest might enhance positive nitrogen balance in cropping systems.
The authors have not declared any conflict of interests.
The authors are grateful to the International Atomic Energy Agency which provided the financial support under Technical Cooperation Project BEN 5005 "Improving maize and yam based cropping systems and soil fertility" by offering training in "the use of 15N in Biological Nitrogen Fixation" and by analyzing plant samples.
REFERENCES
|
Adu-Gyamfi JJ, Myaka FA, Sakala WD, Odgaard R, Vesterager JM, Høgh-Jensen H (2007). Biological nitrogen fixation and nitrogen and phosphorus budgets in farmer-managed intercrops of maize-pigeonpea in semi arid southern and eastern Africa. Plant Soil. 295:127-136.
Crossref
|
|
|
|
Amanuel G, Kühne RF, Tanner DG, Vlek, PLG (2000). Biological nitrogen fixation in faba bean (Vicia faba L.) in the Ethiopian highlands as affected by P fertilization and inoculation. Biol. Fertil. SoilsBiol. Fert. Soils. 32: 353–-359.
Crossref
|
|
|
|
|
Arcand MM, Knight JD, Farrell RE (2013). Estimating belowground nitrogen inputs of pea and canola and their contribution to soil inorganic N pools using 15N labeling. Plant. Soil 371: 67-80.
Crossref
|
|
|
|
|
Bationo A, Koala S, Ayuk E (1998). Fertilité des sols pour la production céréalière en zone sahélo-soudanienne et valorisation des phosphates naturels. Cahiers Agricultures 7 (5): 365-371.
|
|
|
|
|
Birouste M, Kazakou E, Blanchard A, Roumet C (2012). Plant traits and decomposition: are the relationships for roots comparable to those for leaves? Ann. Bot. 109:463-472.
Crossref
|
|
|
|
|
Boddey RM, Peoples MB, Palmer B, Dart PJ (2000). Use of the 15N natural abundance technique to quantify biological nitrogen fixation by woody perennials. Nutri. Cycle. Agroecosyst. 57:235-270.
Crossref
|
|
|
|
|
Chalk PM, (1985). Estimation of N2 fixation by isotope dilution: an appraisal of techniques involving l5N enrichment and their application. Soil Biol. Biochem. 17:, 389-410.
Crossref
|
|
|
|
|
Chalk PM, Smith CJ, Hopmans P, Hamilton, SD (1996). A yield independent, 15N-isotope dilution method to estimate legume symbiotic dependence without a non-N2-fixing reference plant. Biol. Fertil. Soils 23:196-199.
Crossref
|
|
|
|
|
Danso SKA, Bowen GD, Sanginga N (1992). Biological nitrogen fixation in trees in agroecosystems. Plant. Soil. 141:177-196.
Crossref
|
|
|
|
|
De Bruin JL, Pedersen P, Conley SP, Gaska JM, Naeve SL, Kurle JE, Elmore RW, Giesler LJ, Abendroth LJ (2010). Probability of yield response to inoculants in fields with a history of soybean. Crop. Sci. 50: 265-272.
Crossref
|
|
|
|
|
LJ (2010). Probability of Yield Response to Inoculants in Fields with a History of Soybean. Crop. Sci. 50:265â€272.
Crossref
|
|
|
|
|
Forrester DI, Bauhus J, Cowie AL, Vanclay JK (JK (2006). Mixed-species plantations of Eucalyptus with nitrogen-fixing trees: A review. For. Ecol. and Manage. 233:211-230.
Crossref
|
|
|
|
|
Fried M, Middelboe Middleboe V (1977). Measurement of amount of nitrogen fixed by a legume crop. Plant and Soil. 47: 713-715.
Crossref
|
|
|
|
|
Gathumbi SM, Cadisch G, Giller KE (2002). 15N natural as a tool for assessing N2-fixation of herbaceous, shrub and tree legumes in improved fallows. Soil. Biol. Biochem. 34:1059-1071.
Crossref
|
|
|
|
|
Ghosh, PK, Bandyopadhyay, KK, Wanjari, R., Manna MC, Misra , AK, Mohanty M, Rao , AS (2007). Legume effect for enhancing productivity and nitrogen-use efficiency in major cropping systems-an Indian perspective: a review. J. Sustain. Agric. 30 (1): 59-86.
Crossref
|
|
|
|
|
Giller KE (2001). Nitrogen fixation in tropical cropping systems. CAB International, Wallingford. pPp. 166-167.
Crossref
|
|
|
|
|
Graham PH, Vance, CP (2000). Nitrogen fixation in perspective: an overview of research and extension needs. Field. Crops. Res. 65:93-106.
Crossref
|
|
|
|
|
Haque MEHaque ME, Sattar MA (2010). 15N aided Studies on Legume Nitrogen Fixation in Tista Meander Floodplain Soils of Bangladesh. 19th World Congress of Soil Science, Soil Solutions for a Changing World 1-6 August 2010, Brisbane, Australia.
|
|
|
|
|
Hardarson G, Bliss FA, Cigales-Rivero MR, Henson RA, Kipe-Nolt JA, Longeri L (1993). Genotypic variation in biological nitrogen fixation by common bean. Plant Soil , 152:59-70.
Crossref
|
|
|
|
|
Hayat R, Ali S, Siddique MT, Chatha TH (2008). Biological nitrogen fixation of summer legumes and their residual effects on subsequent rainfed wheat yield. Pak. J. Bot., 40(2): 711-722., 2008.
|
|
|
|
|
Herridge DF, Peoples, MB, Boddey, RM (2008). Global inputs of biological nitrogen fixation in agricultural systems. Plant Soil 311:1-18.
Crossref
|
|
|
|
|
Hertenberger G, Wanek W (2004). Evaluation of methods to measure differential 15N labelling of soil and root N pools for studies of root exudation. Rapid Commun. Mass SpectromRapid. Comm. Mass. Spectrom. 18: 2415-2425.
Crossref
|
|
|
|
|
Houngnandan P, Yemadje RGH, Oikeh SO, Djidohokpin CF, Boeckx P, Van Cleemput O (2008). Improved estimation of biological nitrogen fixation of soybean cultivars (Glycine max L. Merril) using 15N natural abundance technique. Biol. Fertil. Soils 45:175-184.
Crossref
|
|
|
|
|
Houngnandan P, Zapata F, Boeckx P, Van Cleemput O (2009). Effect of Bradyrhizobium inoculation and N fertilization on promiscuous soybean and subsequent maize yield grown in degraded "Terre de barre" in Benin. Annales des sciences agronomiques 12 (2): 99-116.
|
|
|
|
|
IAEA (1990). Use of nuclear techniques in studies of soil-plant relationships. Training course series N°2, International Atomic Energy Agency, Vienna, Austria.
|
|
|
|
|
Jaynes DB, Karlen D (2008). Sustainingn soil resources while managing nutrients. Final report: Gulf Hypoxia and local water quality concern workshop. ASABE, St. Joseph MI.
|
|
|
|
|
Kirkegaard J, Christen O, Krupinsky J, Layzell D (2008). Break crop benefit in temperate wheat production. Field. Crops. Res. 107:185-195.
Crossref
|
|
|
|
|
Laberge G, Franke AC, Ambus P. Høgh-Jensen H (2009). Nitrogen rhizodeposition from soybean (Glycine max) and its impact on nutrient budgets in two contrasting environments of the Guinean savannah zone of Nigeria. Nutr. Cycle. Agroecosyst. 84:49-58.
Crossref
|
|
|
|
|
Mafongoya PL, Mpepereki S, Mudyazhezha S (2009). The importance of biological nitrogen fixation in plants. In Nitrogen fixation in crop production, Emerich DW, Krishnan HB (eds). Am.erican Soc.iety of Agron.omy: Madison, pp. 239-264.
|
|
|
|
|
Mayong MV, Houdekon VA, Sanginga PC, Vissoh P, Honlonkou AN (1999). Mucuna fallow diffusion in Southern Benin. IITA Ibadan, Nageria, P 21 p.
|
|
|
|
|
McNeill AM, Pilbeam CJ, Harris HC, Swift RS (1998). Use of residual fertiliser 15N in soil for isotope dilution estimates of N2 fixation by grain legumes. Aust. J. Agric. Res. 49:821-828.
Crossref
|
|
|
|
|
Okito A, Alves B, Urquiaga S, Boddey RM (2004). Isotopic fractionation during N2 fixation by four tropical legumes. Soil. Biol. Biochem. 36:1179-1190.
Crossref
|
|
|
|
|
Parrotta JA, Baker DD, Fried M (1994). Application of 15N enrichment methodologies to estimate nitrogen fixation in Casuarina equisetifolia. Can. J. For. Res. 24:201-207.
Crossref
|
|
|
|
|
Peoples MB, Crasswell ET (1992). Biological nitrogen fixation: investments, expectations, and actual contributions to agriculture. Plant . Soil 141: 13–-39.
Crossref
|
|
|
|
|
Peoples MB, Unkovich MJ, Herridge DF (2009). Measuring symbiotic nitrogen fixation by legumes. In: Nitrogen fixation in crop production. Ed. DW Emerich DW, Krishnan, H.B. Am.erican Soc.iety of Agron. omy: United States of America.
|
|
|
|
|
Reinhold-Hurek B, Hurek T (2003). Endophytic association of Azoarcus spp. In Associative and endophytic nitrogen-fixing bacteria and Cyanobacterial associations, Elmerich C, Newton WE (eds). Kluwer Academic Publishers, The Netherlands, pp. 1-21.
|
|
|
|
|
Reiter K, Schmidtke K, Rauber R (2002). The influence of long-term tillage systems on symbiotic N2 fixation of pea (Pisum sativum L.) and red clover (Trifolium pratense L.). Plant . Soil 238:41-55.
Crossref
|
|
|
|
|
Salvagiotti F, Cassman KG, Specht JE, Walters DT, Weiss A, Dobermann A (2008). Nitrogen uptake fixation and response to fertilizer N in soybeans: a review. Field. Crops. Research. 108:1-13.
Crossref
|
|
|
|
|
Sanginga N, Thottappilly G, Dashiell K (2000). Effectiveness of rhizobia nodulating recent promiscuous soybean selections in the most savanna in Nigeria. Soil Biol. Biochem. 32:127-133.
Crossref
|
|
|
|
|
Sanginga N (2002). Role of biological nitrogen fixation in legume based cropping systems; a case study of West Africa farming systems. Plant . Soil 252: 25-39.
Crossref
|
|
|
|
|
Sanginga N, Dashiell KE, Diel J, Vanlauwe B (2003). Sustainable resource management coupled to resilient germplasm to provide new intensive cereal-grain-legume livestock systems in the dry savanna. Agric. Ecosyst. Environ. 100:305-314.
Crossref
|
|
|
|
|
Schulz S, Carsky RJ, Tarawali SA (2001). Herbaceous legumes: the panacea for West African soil fertility problems? In Sustaining soil fertility in West Africa, Tian G, Keatinge JDH, Ishida F (eds). SSSA special publication no 58, Madison pp. 179–-196.
|
|
|
|
|
Sharma SN (2005). Integrated nutrient management in rice-wheat cropping system. Fertil. News. 50(2):53-71.
|
|
|
|
|
Schipanski M, Drinkwater L, Russelle M (2010). Understanding the variability in soybean nitrogen fixation across agroecosystemsagro ecosystems. Plant . Soil 329:379-397.
Crossref
|
|
|
|
|
Schweiger P, Hofera M, Vollmann J, Wanek W (2014). The relationship between N isotopic fractionation within soybean and N2 ï¬xation during soybean development. Physiol. Plant. 152: 546-557.
Crossref
|
|
|
|
|
Schwenke GD, Peoples MB, Turner GL, Herridge DF (1998). Does nitrogen fixation of commercial, dryland chickpea and faba bean crops in north-west New South Wales maintain or enhance soil nitrogen? Aust. J. Exp. Agric. 38:61–-70.
Crossref
|
|
|
|
|
Singh G, Shivakumar BG (2010). The Role of Soybean in Agriculture. In The soybean: botany, production and uses, Guriqbal Singh (ed): CAB International; pp. 24-47.
|
|
|
|
|
Stahl L, Nyberg G, Högberg P, Buresh RJ (2002). Effects of planted tree fallows on soil nitrogen dynamics, above-ground and root biomass, N2-fixation and subsequent maize crop productivity in Kenya. Plant . Soil 243: 103-117.
Crossref
|
|
|
|
|
Thiessen Martens JR, Hoeppner JW, Entz MH (2001). Legume cover crops with winter cerecrops with winter cereals in southern Manitoba: Establishment, productivity and productivity and microclimate effects. Agron. J. 93:1086-1096.
Crossref
|
|
|
|
|
Thuita M, Pypers P, Herrmann L, Okalebo RJ, Othieno C, Muema E, Lesueur D (. 2012). Commercial rhizobial inoculants significantly enhance growth and nitrogen fixation of a promiscuous soybean variety in Kenyan soils. Biol. Fertil. Soils (2012) 48:87 – -96.
|
|
|
|
|
Unkovich MJ, Pate JS (2000). An appraisal of recent field measurements of symbiotic N2 fixation by annual legumes. Field. Crops. Res. 65:211-228.
Crossref
|
|
|
|
|
Unkovich M, Herridge D, Peoples M, Cadisch G, Boddey R, Giller K, Alves B, Chalk P (2008). Measuring plant-associated nitrogen fixation in agricultural systems. ACIAR Monograph 136, 258 PP.
|
|
|
|
|
Vanlauwe , B, Aihou , K, Houngnandan , P, Diels , J, Sanginga , N, Merckx , R (2001). Nitrogen management in adequate 'input' maize-based agriculture in the derived savanna benchmark zone of Benin Republic. Plant Soil 228: 61-71.
Crossref
|
|
|
|
|
Vassiliki B, Salvati L, Di Bartolomei R, Constantinos A (2012). Land Evaluation and Agri-Environmental Indicators: Exploring Spatial Trends of Nitrogen Balance in Greece. Int. J. Latest Trends in Financ. and Econ.omic Sci. Int J. Latest. Trends. Fin. Eco Sc 2 (4): 330-336.
|
|

.png)