Full Length Research Paper

ABSTRACT
The aim of this research was to evaluate the capacity of symbiotic efficiency of native rhizobia from soils of the state of Rio Grande do Sul (RS), Brazil in Phaseolus lunatus L. plants. Soil and nodule samples from predominant legume plants such as Desmodium species were collected in seven locations of the state of RS. For the isolation of the rhizobia from the soil samples, lima bean “olho de cabra preto” variety and “leucena” (Leucena leucocephala) and plants were used as baits. Subsequently, the symbiotic characterization of the isolates was performed by inoculation on lima bean plants in vitro conditions. The selected bacterial isolates were evaluated for biological nitrogen fixation efficiency in a greenhouse experiment, being determined after: shoot dry mass, root dry mass, nodule dry mass, nitrogen (N) accumulated shoot and calculated the N fixation relative efficiency index. Among the 28 rhizobia isolates tested, 11 induced nodule formation in lima bean. The rhizobia Plu03 and Plu14 stimulated a greater increase of plant dry mass, nodule dry mass and nitrogen accumulation in the shoot, reflecting in a higher relative efficiency index. These results suggest that the isolates Plu03 and Plu14 are more efficient in promoting growth of lima bean, which could be recommended for future agronomic efficiency studies.
Key words: Rhizobia, lima bean, biological nitrogen fixation.
INTRODUCTION
Lima bean (Phaseolus lunatus L.), also known in Brazil as “feijão-fava” or “espadinho” beans in Brazil are considered the second most important species of the genera Phaseolus, after common beans (Phaseolus vulgaris L.). Due to its characteristics as drought and excess moisture resistance and protein potential, representing a source of food and income for small farmers (Fofana et al., 1997; Santos et al., 2009; Vieira, 1992).
In Brazil, it is cultivated in all Brazilian regions, mainly in the northeast region; especially the states of Paraíba, Ceará, Pernambuco, Rio Grande do Norte and Piauí (Vieira, 1992; Santos, 2008; Antunes et al., 2011; Araujo et al., 2015; De Araujo et al., 2016). In the state of Rio Grande do Sul, the species has a diversity of genotypes, the variety “olho de cabra preto”, “olho de cabra vermelho” and “Branco”. However, this bean is cultivated by small producers and its production is limited, due to the tradition of consuming common beans and the lack of cultivars recommended for the subtropical climatic conditions of the region (Cavalheiro, 2012).
This crop has the ability to establish symbiosis with rhizobia, where biological nitrogen fixation (BNF) occurs that considered one forms of sustainable increase in legume productivity and allows mineral nitrogen fertilizers replacement (Franco et al., 2002). The study of the diversity and symbiotic association of rhizobia with P. lunatus was limited to countries with research centers such as Peru and Mexico, where it has been reported that P. lunatus forms symbiosis mainly with Bradyrhizobium species (Ormeño-Orrillo et al., 2006; López-López et al., 2013; Durán et al., 2014). Two new species of Bradyrhizobium (Bradyrhizobium paxllaeri and Bradyrhizobium icense) were reported in Peru (Durán et al., 2014). In addition, it was reported that strains of the genera Rhizobium and Sinorhizobium also induced the formation of lima bean nodules (Ormeño-Orrillo et al., 2007).
However, in Brazil, studies with lima bean symbiotic rhizobia are limited because P. lunatus is not a native legume, but a great morphological and physiological diversity has been found among native nodulant rhizobia of P. lunatus in the northeast of Brazil, being reported the genera Bradyrhizobium, Sinorhizobium, Mesorhizobium and Rhizobium (Araujo et al., 2015; Santos et al., 2011). The studies demonstrated that although some rhizobia have a restricted host range in P. lunatus, low host specificity is observed and symbiosis with rhizobia strains of more than four genera is established (Santamaría et al., 2014).
The selection of rhizobia isolates efficient in BNF in P. lunatus are scarce in Brazil, with only a few studies performed in the northeast region of Brazil. Among these studies, the efficiency BNF of rhizobia isolates in P. lunatus genotype UFPI-468 “fava-miuda” variety compared to strain SEMIA 4077 - CIAT 899 (strains recommended for common bean - P. vulgaris inoculant production) and SEMIA 6357 (NGR 234) symbiont of diversity legumes has been evaluated. The authors observed that eight of the rhizobia isolates contributed to the increase in shoot dry mass, higher N accumulation, reflecting a higher efficiency of these isolates in BNF, compared to the reference strains (Antunes et al., 2011). Similarly, in another study, four isolates of the genera Bradyrhizobium and Rhizobium were evaluated for their symbiotic performance in “boca de moça” and “branca” varieties. The authors reported that plants inoculated with isolates of the genera Bradyrhizobium showed higher growth, nodulation and nitrogen content, reflecting higher photosynthetic efficiency compared to plants inoculated with Rhizobium isolates (Da Costa Neto et al., 2017).
Currently, in the state of Rio Grande do Sul, Brazil, there are no native isolates or strains released like inoculants for P. lunatus. Therefore, there is a need to isolate and select native rhizobia symbionts of P. lunatus, efficient in BNF that contribute to increase in sustainable productivity. In this sense, the objective of this research was to evaluate the capacity of symbiotic efficiency of native rhizobia from soils of the state of Rio Grande do Sul in P. lunatus plants.
MATERIALS AND METHODS
Study site
The study was conducted at the Soil Microbiology Laboratory and greenhouse of the Soil Department of the Federal University of Rio Grande do Sul (UFRGS). Soil samples were collected from different locations in rural area of Rio Grande do Sul (Table 1). The soil samples were collected at a depth of 0-20 cm, considering the presence of native or introduced leguminous plants. Root nodules were also collected from Desmodium species plants at the UFRGS Agronomic Experimental Station (EEA), in Eldorado do Sul – RS, Brazil.
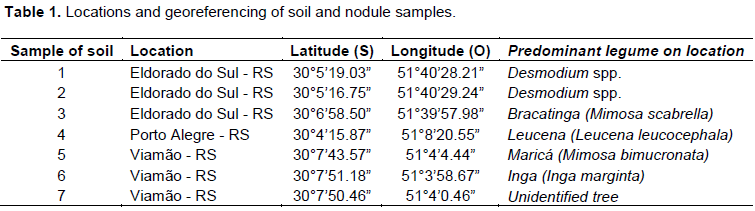
Isolation and characterization of rhizobia colonies from soil samples and nodules collected in the field
The soil samples were taken to the Soil Microbiology Laboratory where a suspension of 10 g of soil in 90 mL of sterile saline solution (NaCl 0.85%) was performed on horizontal shaker for 3 h. In the installation of the experiment, lima bean seeds “olho de cabra preto” variety provided by the producer and “leucena” (Leucaena leucocephala) collected at the EEA-UFRGS as bait plants to obtain nodules were used. The seeds were disinfected by successive immersion in alcohol (70%) for 30 s, sodium hypochlorite (2.5%) for 30 s and immediately received six consecutive washes with sterile distilled water (Vincent, 1970). Then, seeds of leucena and lima bean were seeded separately in plastic pots with a mixture of vermiculite and sand (2:1) and inoculated immediately with addition of 2 mL of the suspensions of the soil samples in each pot, with 5 replications for each soil sample. The plants of leucena was irrigated with Sarruge (1975) nutrient solution and lima beans with Hoagland and Arnon (1950) nutrient solution modified by Silveira et al. (1998) during the experiment and cultivated under laboratory conditions.
After 45 days from inoculation, the root of the leucena and lima bean plants were removed and nodules with alcohol (70%) for 30 s, followed by sodium hypochlorite solution (1%) for 30 s and 7 washes with sterile distilled water. Then, in laminar flow chamber, each nodule were placed in sterile test tubes and macerated with sterile glass rod. The suspension of the macerated nodules was inoculated on plates with Yeast Mannitol Agar with Congo Red (YMA+CR) (Vincent, 1970) in laminar flow chamber using either the drop method (Miles et al., 1938) or streak scattering method separately (Buck and Cleverdon, 1960). After the plates were incubated in an incubator at 28 ± 2°C for 10 days, the nodules of Desmodium spp. collected in the field were disinfected and processed in the same way as the leucena and lima bean nodules described earlier.
All bacterial isolates were transferred to other plates with YMA+CR to obtain colonies with persistent morphology, and daily evaluation of bacterial colony growth in order to analyze the colony morphology. The colony diameter and color, border type, opacity and consistency, and growth time were also observed. In addition, the pH changes of the YMA with bromothymol blue (YMA+BB) was evaluated. All isolates obtained were maintained in LMA medium test tubes (Vincent, 1970) in refrigerator.
Evaluation of rhizobia isolates symbiotic capacity in lima bean plants
Rhizobia isolates obtained from nodules of lima bean and Desmodium spp., in addition to the rhizobia isolates from leucena nodules were evaluated for their ability to induce nodulation in lima bean plants growing on laboratory conditions. For this, each isolate was inoculated into falcon tubes with 30 mL of yeast mannitol (YM) broth (Vincent, 1970) and kept in an orbital incubator at 28°C±2 with agitation of 120 rpm to achieve a concentration of 108 cells.mL-1.
The seeds previously disinfected were placed in 700 mL plastic pots containing a mixture of vermiculite and sand (2:1) and then inoculated with 1 mL of broth from each bacterial isolate with three replications. During the experiment the plants were irrigated with Hoagland and Arnon (1950) nutrient solution modified by Silveira et al. (1998) diluted to 50% without nitrogen. Also, an uninoculated control treatment was conducted to ensure there was no cross contamination. After a period of 45 days the experiment was finished, after observing the root nodules formed and the presence of red color, indicating symbiotic nitrogen fixation.
Evaluation of symbiotic efficiency in nitrogen fixation of rhizobia isolates in lima bean plants
This experiment was conducted under greenhouse conditions. The rhizobia tested, 11 isolates from lima bean (Plu01, Plu02, Plu03, Plu05, Plu06, Plu08, Plu08, Plu09, Plu11, Plu13 and Plu14) and the strains SEMIA 4077 (Rhizobium tropici CIAT 899) and SEMIA 6357 (NGR 234) were evaluated. The strain SEMIA 4077 (R. tropici CIAT 899) is released by the Ministry of Agriculture, Livestock and Food Supply (MAPA) from Brazil for the production of commercial inoculants to common bean (P. vulgaris) and SEMIA 6357 has been included in the experiment due to BNF efficiency in cowpea (genotype “UFPI-468” – “Fava-miúda” variety) (Antunes et al., 2011). These strains were obtained from the rhizobia collection SEMIA of the Department of Diagnostic and Agricultural Research (DDPA) from Brazil. The inoculum from each of the isolates and strains were incubated to a concentration of 108 cells.mL-1 in YM broth, the same way as previously described in the evaluation of the symbiotic capacity of the isolates.
In this experiment, seeds of lima bean were used, which were previously disinfected by the method described earlier. After disinfection, the seeds were planted in 1.5 L plastic pots, containing a mixture of vermiculite and sand (2:1) and then inoculated with 2 mL of broth from each isolate studied. One week later roughing was done leaving one plant per pot and put a sterile iron stand to help the plant curl up and stand up.
The experimental design was randomized blocks with 14 inoculated treatments and two controls non-inoculated, one without nitrogen (Control-N) and the second with addition of nitrogen (Control+N) at a dose of N equivalent to 100 kg.ha-1 receiving NH4NO3, being divided into 5 applications of 10 mL of a solution of (4.28 g.L-1) NH4NO3 during the experiment. After the 45 days of cultivation period, the plants were collected by separating the shoot from the root system. Shoot dry mass (SDM), root dry mass (RDM), and nodule dry mass (NDM) were evaluated, and then the nitrogen accumulated N (Nac) in the shoot. For this, the shoot was packed in paper bags and dried in oven with forced air circulation at 65°C for three days and then ground for the quantification of nitrogen by the method described by Tedesco et al. (1995). The roots were washed to remove adhered substrate particles and the nodules were removed and dried separately from the bag roots under the same conditions as earlier and weighed in the balance. The relative efficiency index (REI) was determined according to Brockwell et al. (1966) of BNF of rhizobia isolates and strains.
The REI was calculated using the formula:
REI = ((NT - NT-N) / (NT + N - NT-N)) × 100
where NT = total nitrogen of plant of inoculated treatment; NT-N = total nitrogen of control uninoculated and without nitrogen; NT + N = total nitrogen of control uninoculated and receiving nitrogen supplementation (Brockwell et al., 1966).
Statistical analysis
The data obtained were submitted to the analysis of variance, being the comparison of averages performed by the test of Tukey (p<0.05) using the statistical program SPSS 15.
RESULTS
Rhizobia isolates obtained
With the sampling and isolation studies, 28 bacterial isolates were obtained from soil samples and nodules: 14 isolates were obtained from soil samples (6 and 7) using lima bean as bait; 12 were isolated of nodules of Desmodium spp. collected in the field at location Eldorado do Sul; and 2 isolates were obtained from soil sample 4 using leucena as bait (Table 2).
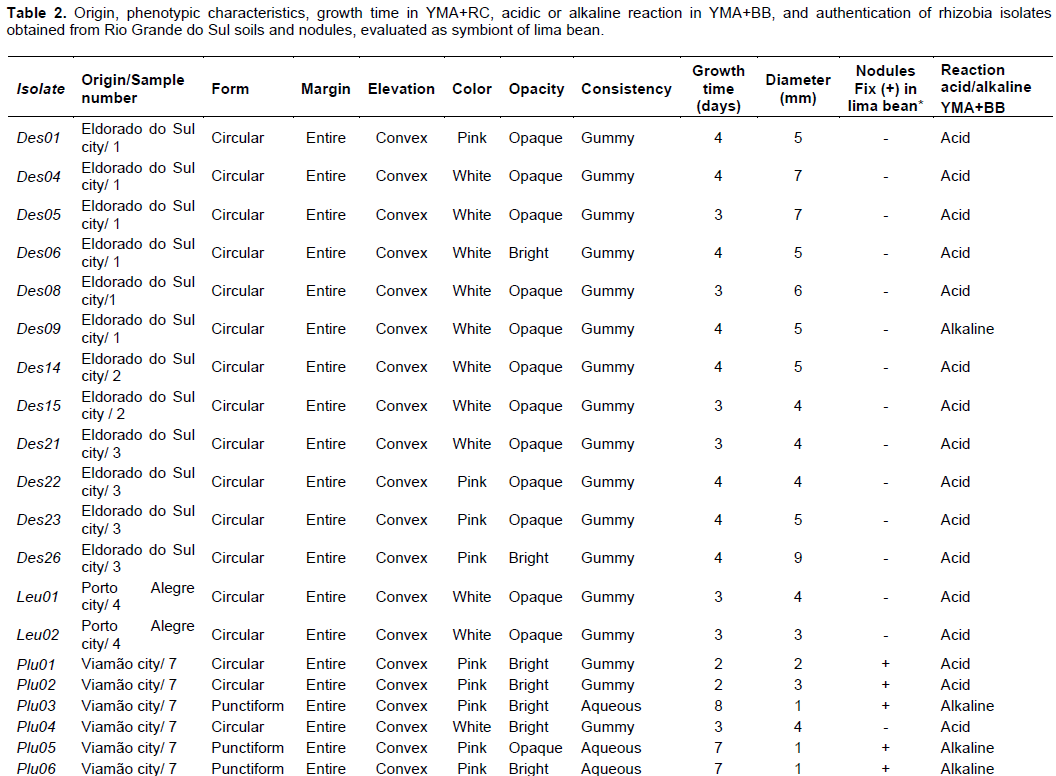

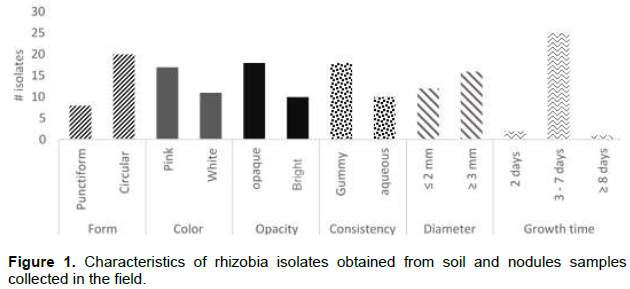
All isolates showed colony morphology with convex elevation and entire border, in addition to the characteristics such as: surface form (circular or punctiform), color (white or pink), opacity (opaque or translucent), consistency (gummy or watery), time of first colony onset (between 2 - 8 days) and diameter (≤ 1mm or more) could be presumptive of rhizobia (Figure 1).
For the production of acid alkali related to the reaction in YMA+BB medium, 14 isolates of lima bean were observed, six produced acid reaction (Plu01, Plu02, Plu04, Plu07, Plu09, Plu13) and eight isolates produced alkaline reaction (Plu03, Plu05, Plu06, Plu08, Plu11, Plu12, Plu14). Some bacterial isolates such as Plu03, Plu05, Plu06, besides having growth 7-8 days, generated alkali production in YMA+BB medium, characteristics that could be presumptive of Bradyrhizobium genera (Somasegaran and Hoben, 1994). Among the rhizobia isolate of nodule of Desmodium spp., 11 produced acid reaction (Des01, Des04, Des05, Des06, Des08, Des14, Des15, Des21, Des22, Des22, Des26) and one (Des09) produced alkaline reaction. The leucena rhizobia isolates Leu01 and Leu02 produced acid reaction.
For the production of acid alkali related to the reaction in YMA+BB medium, 14 isolates of lima bean were observed, six produced acid reaction (Plu01, Plu02, Plu04, Plu07, Plu09, Plu13) and eight isolates produced alkaline reaction (Plu03, Plu05, Plu06, Plu08, Plu11, Plu12, Plu14). Some bacterial isolates such as Plu03, Plu05, Plu06, besides having growth 7-8 days, generated alkali production in YMA+BB medium, characteristics that could be presumptive of Bradyrhizobium genera (Somasegaran and Hoben, 1994). Among the rhizobia isolate of nodule of Desmodium spp., 11 produced acid reaction (Des01, Des04, Des05, Des06, Des08, Des14, Des15, Des21, Des22, Des22, Des26) and one (Des09) produced alkaline reaction. The leucena rhizobia isolates Leu01 and Leu02 produced acid reaction. In the rhizobia authentication, the isolates Plu01, Plu02, Plu03, Plu06, Plu07, Plu08, Plu09, Plu11, Plu13 and Plu14 induced nodules and produce leghemoglobin activity in plants of lima bean, being selected to be evaluated in a greenhouse experiment to evaluate the BNF efficiency (Table 2).
Symbiotic efficiency of rhizobia isolates in plants of lima bean
The values of production of shoot dry mass (SDM), root dry mass (RDM), dry nodule mass (DNM), accumulated nitrogen (Nac) in shoot and Relative Efficiency Index (REI) (Brockwell et al., 1966) obtained from lima bean cultivate in a greenhouse, are shown in Table 3. A higher shoot dry mass in treatments inoculated with isolate Plu14 was observed, followed by Plu03 and SEMIA 4077 compared to treatment control with addition of nitrogen (Control+N). Plants inoculated with SEMIA 4077 strain produced similar SDM to Control+N while those inoculated with strain SEMIA 6357 was lower. The other treatments obtained values below the Control+N.
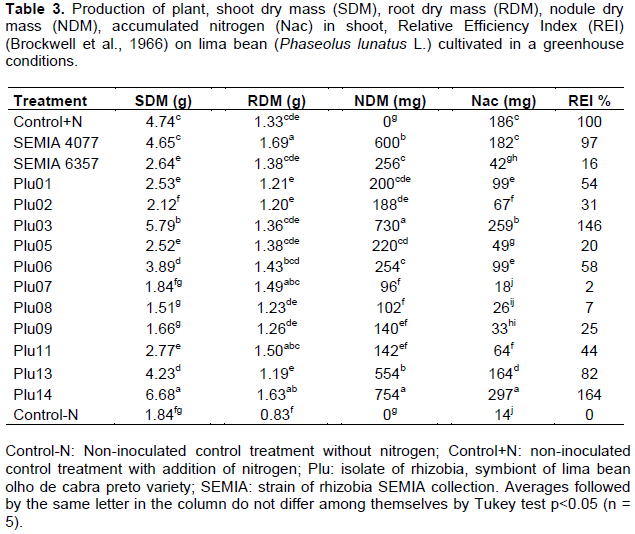
In the variable, root dry mass (RDM) was found that the plants inoculated with Plu14 and SEMIA 4077 showed higher RDM compared with Control+N. The other treatments were similar to the Control+N with exception of control non-inoculated without N (Control-N).
The nodules dry mass (NDM) varied between inoculated treatments. The highest values in plant inoculated with isolates Plu14, Plu03, Plu13 and the strain SEMIA 4077 were obtained, being the isolates Plu14 and Plu03 superior to strain SEMIA 4077 (p<0.05). The plants inoculated with strain SEMIA 6357, followed by inoculated treatments Plu01, Plu02, Plu05, Plu06, Plu07, Plu08, Plu09, Plu11 and Plu13 have low NDM values found, compared to strain SEMIA 4077 recommended by MAPA for common bean (Table 3). Related to the nitrogen fixation capacity, the isolates Plu14 and Plu03 contributed to a greater accumulation of N in the shoot of plants compared to the plants non-inoculated with addition of N (Control +N) and strain SEMIA 4077 with significant differences (p<0.05), being the isolate Plu14 superior to all treatments (Table 2). In addition, treatment with strain SEMIA 4077 was observed to be similar to non-inoculated control treatment Control +N, while treatment with SEMIA 6357 had a low N accumulation compared to Control + N. The other isolates and Control-N were inferior to the non-inoculated treatment (Control + N).
In this study, the Relative Efficiency Index (REI) according to Brockwell et al. (1966), the isolates Plu14 and Plu03 stood out as the most efficient in biological nitrogen fixation, obtaining an REI of 164 and 146 respectively, exceeding 100% of control treatment non-inoculated with addition of N (control+N). The treatment with strain SEMIA 4077 obtained an REI of 97%, followed by treatment with Plu13 isolate of 82%, values close to 100% obtained by Control+N. However, a low REI was observed in the treatment with strain SEMIA 6357 equivalent to 16% and 2-58% in the other isolates treatments.
DISCUSSION
Isolation, selection and diversity studies of rhizobia with ability to perform symbiosis with lima bean are still few in Brazil, possibly due to the fact that lima bean (P. lunatus L.) is not a native legume in the country (Araujo et al., 2015). Some studies developed in Peru, where it is considered native, Mexico and in the northeast of Brazil, have shown that lima beans can be considered promiscuous because they have low host specificity and form symbiosis with isolates of four rhizobia genera: Bradyrhizobium, Mesorhizobium, Rhizobium and Sinorhizobium (Ormeño-Orrillo et al., 2006; Ormeño-Orrillo et al., 2007; Santos et al., 2011; Antunes et al., 2011; López-López et al., 2013; Santamaria et al., 2014; Duran et al., 2014; Araujo et al., 2015; Servín-Garcidueñas et al., 2014; Ormeño-Orrillo et al., 2017).
In this study, it was possible to obtain several bacterial isolates on YMA+RC medium of soil samples collected from different location of the state of Rio Grande do Sul and from root nodules of Desmodium spp., being the highest number of rhizobia symbiont of lima bean from soil samples using the same plants as bait. Furthermore, the results showed that the rhizobia obtained from Desmodium spp. and L. leucocephala nodules as bait in soil sample, are not able to form nodules in lima bean olho de cabra preto variety in the symbiotic evaluation test. This is possible because the symbiotic association and nodule formation are dependent on both plant genotypes as inoculated isolate and environmental conditions (Moreira and Siqueira, 2006).
Among the rhizobia isolates obtained as lima bean symbionts in this study, some have characteristics that distinguish the genera Bradyrhizobium. Similar results were observed in isolates obtained from nodules of lima bean crops in Peru soils, where the genera Bradyrhizobium was the predominant symbiont (Ormeño-Orrillo et al., 2006). However, other rhizobia obtained in this study showed rapid growth and acid production, similar to characteristics found in isolates identified as Rhizobium spp. obtained from lima bean variety “fava-miúda” (Santos et al., 2011), but the identification of the genera and species of rhizobia of these isolates obtained in this study requires molecular identification by 16S.
In the evaluation of symbiotic efficiency of isolates of rhizobia, the shot dry mass (SDM) of plants besides being considered an indicator of nutritional status variable in the production of plant cultivation, allows the selection of new isolates to be suggested as strains for the production of inoculants (Xavier et al., 2006; Zilli et al., 2006; Souza et al., 2008). In this study, the results showed that Plu03 and Plu14 isolates provided a higher production of SDM compared to non-inoculated control treatments (Control-N and Control+N) and the strain recommended for common bean SEMIA 4077. Similar results were found by Antunes et al. (2011) in northeastern Brazil, in the state of Piauí, where they evaluated the efficiency of some isolates symbiont of UFPI-468 genotype, collected and described by Santos (2008), using as criteria the SDM of plants, SEMIA 4077 and SEMIA 6357 (NGR 234) strains and an absolute non-inoculated nitrogen free control (Control-N) as controls. The authors found that 12 isolates evaluated generated in plants a SDM similar to the two strains SEMIA 4077 and SEMIA 6357, which were efficient and higher than the absolute control (Control-N). However, these results also contrast with those obtained in the study in lima bean (Table 3) with strain SEMIA 6357, where low SDM of plants was found under both strain SEMIA 4077, Plu03 and Plu14 isolates and non-inoculated treatment control (Control-N).
Other similar result was found in one study from the state of Piauí, Brazilian northeast, where native isolates of genera Rhizobium and Bradyrhizobium were evaluated in lima bean plants of “Boca de Moça” and “Branca” varieties in three evaluation periods (37 days after germination, 50 days after pod emission and 63 days in the declining nitrogen fixation period). The authors reported that plants inoculated with Bradyrhizobium (ISOL18) showed a higher continuous increase of SDM of plants during the three periods (Da Costa Neto et al., 2017).
In the variable root dry mass (RDM) of plants, the results showed that the plants inoculated with Plu14 isolated and the SEMIA 4077 strain influenced a higher increase of root growth of lima bean plants, being superior to the non-inoculated control treatments (Control + N and Control N). Similar results were found by Da Costa Neto et al. (2017) in the “Boca de Moça” and “Branca” varieties of lima bean inoculated with Rhizobium and Bradyrhizobium isolates, where they showed that Bradyrhizobium isolate (ISOL18) increased the RDM of plants during the three periods of evaluation. Contrary to the results obtained by Antunes et al. (2011) in lima bean genotype UFPI-468, where rhizobia isolates had no influence on root growth compared to control (Control-N).
Regarding the nodules dry mass (NDM) is a widely used variable as an indication of nodulation, which allows to determine the best performance of symbiotic association (Döbereiner, 1966; Bohrer and Hungria, 1998; Hungria and Bohrer, 2000; Ferreira and Castro, 1995; Araújo et al., 2007), being evidenced in this study. The highest performance in the isolates Plu14, Plu03 and Plu13, and strain SEMIA 4077, standing out is isolates Plu14 and Plu03 as superior to strain SEMIA 4077. Similar results were found by Da Costa Neto et al. (2017) where the isolate ISOL18 (Bradyrhizobium spp.) induced a higher NDM in lima bean plants Boca de Moça and Branca varieties. In the same way, Antunes et al. (2011) reported that seven isolates of lima bean genotype UFPI-468 showed the best nodulation performance, similar to the SEMIA 4077 and SEMIA 6357 strains used as control. Although in our study with lima bean plants inoculated with the SEMIA 6357 strain showed low nodulation, as did the other isolates (Table 3) and these results can be explained by the fact that nodule formation is dependent on plant genotype, isolate or inoculated strains and environmental conditions (Moreira and Siqueira, 2006) varieties compared to Rhizobium spp. which showed a low nodulation.
In relation to the nitrogen accumulated (Nac) in shoot of the plants the results indicate that the treatments with the isolates Plu14 and Plu03 presented higher ability to fix N atmospheric by symbiotic association and supply it to the lima bean plant in conditions of this experiment, being superior to the non-inoculated control treatment without N (Control-N), non-inoculated control with N addition (Control + N) and SEMIA 4077 strain. These results are similar and in agreement with those found by Antunes et al. (2011), which showed that eight of the isolates tested in lima bean genotype UFPI-468 also had higher ability to fix N atmospheric. Likewise, the results agree with that found by Da Costa Neto et al. (2017), where the isolate ISOL18 stood out for increasing Nac in lima bean Boca de Moça and Branca varieties.
In addition, the strain SEMIA 4077 that had not been evaluated on lima bean olho de cabra preto variety plants facilitated a higher nitrogen accumulation in the shoots similar to non-inoculated control treatment with addition of N (Control + N). This opens the possibility of being recommended for this variety of lima bean in addition to the genotype UFPI-468 where this same strain had higher ability to fix nitrogen (Antunes et al., 2011). However, SEMIA 6357 induced a lower accumulation of N in the shoot compared to the control (Control + N), differing from that found by Antunes et al. (2011) in the genotype UFPI- 468, where strain SEMIA 6357 showed higher Nac.
The relative efficiency index – REI (Brockwell et al., 1966) indicates the percentage efficiency of bacteria on nitrogen fixation in symbiotic association with legumes. The results in this study showed that Plu03 and Plu14 isolates were the most efficient, due to the REI values exceeded 100%, above the minimum efficiency of 70% expected from rhizobia potentially efficient (Miranda, 1995). These isolates were the same ones that also presented higher SDM, RDM, DNM and Nac. Likewise, in a study with lima bean genotype UFPI-468, the authors found eight isolates efficient in biological nitrogen fixation (Antunes et al., 2011) using the efficiency index according to Faria and Franco (2002).
In case of SEMIA 4077 and SEMIA 6357 strains, the results showed that only SEMIA 4077 strain showed a REI above 70% minimum, differing from that found by Antunes et al. (2011) in the lima bean UFPI-468 genotype where SEMIA 6357 was efficient.
CONCLUSION
The rhizobia isolates Plu03 and Plu14 are efficient in biological nitrogen fixation in lima bean plants, as well as the strain SEMIA 4077 and might be recommended for future agronomic efficiency studies within different soils, both in the greenhouse and in the field for lima bean olho de cabra preto variety.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
The authors thank the program of Alliances for Education and Training (PAEC) between the Organization of American States (OAS) and the Coimbra Group of Brazilian Universities (GCUB), together with the academic excellence program of the Coordination for the Improvement of Higher Education Personnel (CAPES-PROEX) for granting a doctoral scholarship and graduate program in Soil Science at the Federal University of Rio Grande do Sul, Brazil for the support.
REFERENCES
|
Antunes L, Gomes F, Lopes A, Araújo F, Lyra P, Figueiredo B (2011). Eficiência simbiótica de isolados de rizóbio noduladores de feijão-fava (Phaseolus luntaus L.). Revista Brasileira de Ciência do Solo 35:751-757. |
|
|
Araujo F, Lopes A, Gomes F, Junior B, Antunes L, Lyra P, Do Vale M (2015). Diversity of native rhizobia-nodulating Phaseolus lunatus in Brazil. Legume Research - An International Journal 38(5):653-657. |
|
|
Brockwell J, Hely W, Neal-Smith A (1966). Some symbiotic characteristics of rhizobia responsible for spontaneous, effective field nodulation of Lotus hispidus. Australian Journal of Experimental Agriculture and Animal Husbandry 6(23):365-370. |
|
|
Bohrer J, Hungria M (1998). Avaliação de cultivares de soja quanto à fixação biológica do nitrogênio. Pesquisa Agropecuária Brasileira 33:937-952. |
|
|
Buck D, Cleverdon C (1960). The spread plate as a method for the enumeration of marine bacteria. Limnology and Oceanography Waco 5:78-80. |
|
|
Cavalheiro V (2012). Caracterização de genótipos de feijão-lima (Phaseolus lunatus l.) na região de pelotas - Rio Grande do Sul. 2012. Dissertação (Mestrado em Ciência e Tecnologia de Sementes) - Universidade Federal de Pelotas, Pelotas. |
|
|
Da Costa Neto P, Mendes S, de Araújo F, de Alcântara Neto F, Bonifacio A, Rodrigues C (2017). Symbiotic performance, nitrogen flux and growth of lima bean (Phaseolus lunatus L.) varieties inoculated with different indigenous strains of rhizobia. Symbiosis 73(2):117-124. |
|
|
De Araujo F, de Almeida Lopes C, Teran M, Palkovic A, Gepts P (2016). Nodulation ability in different genotypes of Phaseolus lunatus by rhizobia from California agricultural soils. Symbiosis 73(1):7-14. |
|
|
Döbereiner J (1966). Evaluation of nitrogen fixation in legumes by the regression of total plant nitrogen with nodule weight. Nature 210:850-852. |
|
|
Durán D, Rey L, Mayo J, Zúñiga-Dávila D, Imperial J, Ruiz-Argüeso T, Martínez-Romero E, Ormeño-Orrillo E (2014). Bradyrhizobium paxllaeri sp. nov. and Bradyrhizobium icense sp. nov., nitrogen-fixing rhizobial symbionts of Lima bean (Phaseolus lunatus L.) in Peru. International Journal of Systematic and Evolutionary Microbiology 64:2072-2078. |
|
|
Faria M, Franco A (2002). Identificação de bactérias eficientes na fixação biológica de nitrogênio para espécies leguminosas arbóreas. Seropédica, Embrapa Agrobiologia 16 p. (Documentos, 158). |
|
|
Fofana B, Vekemans X, Jardin P, Baudoin P (1997). Genetic diversity in lima (Phaseolus lunatus L.) as revealed by RAPD markers. Euphytica 95:157-165. |
|
|
Franco C, Cassini A, Oliveira R, Vieira C, Tsai M (2002). Nodulação em cultivares de feijão dos conjuntos gênicos andino e meso-americano. Pesquisa Agropecuária Brasileira 37:1145-1150. |
|
|
Hoagland R, Arnon I (1950). The water culture method of growing plants without soil. Berkeley. University of California 32 p. |
|
|
Hungria M, Bohrer J (2000). Variability of nodulation and dinitrogen fixation capacity among soybean cultivars. Biology and Fertility of Soils 31:45-52. |
|
|
López-López A, Negrete-Yankelevich S, Rogel A, Ormeño-Orrillo E, Martínez J, Martínez-Romero E (2013). Native bradyrhizobia from los Tuxtlas in Mexico are symbionts of Phaseolus lunatus (lima bean). Systematic and Applied Microbiology 36:33-38. |
|
|
Miles A, Misra S, Irwin O (1938). The estimation of the bactericidal power of the blood. Epidemiology and Infection 38(6):732-749. |
|
|
Moreira S, Siqueira O (2006). Fixação Biológica do Nitrogênio. In: |
|
|
Moreira, S., Siqueira, O. Microbiologia e bioquímica do solo. 2.ed. Lavras. Universidade Federal Lavras pp. 501-529. |
|
|
Ormeño-Orrillo E, Vinuesa P, Zúñiga-Dávila D, Martínez-Romero E (2006). Molecular diversity of native bradyrhizobia isolated from Lima bean (Phaseolus lunatus L.) in Peru. Systematic and Applied Microbiology 29:253-262. |
|
|
Ormeño-Orrillo E, Rey L, Durán D, Canchaya A, Rogel A, Zúñiga-Dávila D, Martínez-Romero E (2017). Draft genome sequence of Bradyrhizobium paxllaeri LMTR 21 T isolated from Lima bean (Phaseolus lunatus) in Peru. Genomics Data 13:38-40. |
|
|
Ormeño-Orrillo E, Torres R, Mayo J, Rivas R, Peix A, Velázquez E, Zúniga D (2007). Phaseolus lunatus is nodulated by a phosphate solubilizing strain of Sinorhizobium meliloti in a Peruvian soil. In: Velázquez El, Rodríguez-Barrueco C (eds) Developments in Plant and Soil Sciences. Dordrecht. Springer pp. 243-247. |
|
|
Santamaría I, Bustos P, Sepúlveda-Robles O, Lozano L, Rodríguez C, Fernández L, Juárez S, Kameyama L, Guarneros G, Dávila G, González V (2014). Narrow-host-range bacteriophages that infect Rhizobium etli associate with distinct genomic types. Applied and Environmental Microbiology 80:446-454. |
|
|
Santos O (2008). Divergência genética em feijão-fava (Phaseolus lunatus L.): diversidade genética entre isolados nativos de rizóbios noduladores do feijão-fava (Phaseolus lunatus L.). Dissertação (Mestrado em Agronomia) - Universidade Federal do Piauí, Teresina. |
|
|
Santos O, Araújo F, Gomes F, Lopes A, Figueiredo B (2009). Ontogenia da nodulação em feijão-fava (Phaseolus lunatus). Revista Brasileira de Ciências Agrárias 4(4):426-429. |
|
|
Santos O, Antunes L, Lyra P, Araújo F, Gomes F, Lopes A, and B. (2011). Genetic diversity among isolates of Rhizobia from Phaseolus lunatus. Annals of Microbiology 61(3):437-444 |
|
|
Servín-Garcidueñas E, Zayas-Del Moral A, Ormeño-Orrillo E, Rogel A, Delgado-Salinas A, Sánchez F, Martínez-Romero E (2014). Symbiont shift towards Rhizobium nodulation in a group of phylogenetically related Phaseolus species. Molecular Phylogenetics and Evolution 79:1-11. |
|
|
Silveira G, Contado L, Mazza M, Oliveira A (1998). Phosphoenolpyruvate carboxylase and glutamine synthetase activities in relation to nitrogen fixation in cowpea nodules. Revista Brasileira de Fisiologia Vegetal 10:9-23. |
|
|
Somasegaran P, Hoben J (1994). Handbook for Rhizobia: Methods in legume-rhizobium technology. New York. Springer-Verlag. |
|
|
Souza A, Hungria M, Franchini C, Maciel D, Campo J, Zaia M (2008). Conjunto mínimo de parâmetros para avaliação da microbiota do solo e da fixação biológica do nitrogênio pela soja. Pesquisa Agropecuária Brasileira 43(1):83-91. |
|
|
Tedesco J, Gianello C, Bissani A, Bohnen H, Volkweiss J (1995). Análises de solo, plantas e outros materiais. 2.ed. Porto Alegre, Universidade Federal do Rio Grande do Sul P 174. |
|
|
Vieira F (1992). A cultura do feijão-fava. Informe Agropecuário 16(174):30-37. |
|
|
Vincent M (1970). A manual for the practical study of root nodule bacteria. Oxford. Blackwell Scientific, International Biological Programme Handbook 15, 164 p. |
|
|
Xavier R, Martins V, Ribeiro A, Rumjanek G (2006). Especificidade simbiótica entre rizóbios e acessos de feijão-caupi de diferentes nacionalidades. Caatinga 19:25-33. |
|
|
Zilli E, Valicheski R, Rumjanek G, Simões Araújo L, Freire Filho R, Neves P (2006). Eficiência simbiótica de estirpes de Bradyrhizobium isoladas de solo do Cerrado em caupi. Pesquisa Agropecuária Brasileira 41:811-818. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


