Full Length Research Paper

ABSTRACT
The genus Capsicum presents a wide genetic variability. The most common way to determine this variability has been based upon morphological descriptors. We studied the genetic divergence among populations of ornamental pepper (Capsicum annuum L.) using two different multivariate techniques: Cluster analysis and canonical discriminant variables. The analyses enabled us to determine the morphological descriptors that contributed most to the genetic divergence. The study was carried out in a greenhouse in the Northeastern Brazil, in two years: 2013 and 2014. The experimental design was the completely randomized design, considering two crossed factors: Population and years. Thirteen populations of ornamental pepper were evaluated based on sixteen plant descriptors, six flower descriptors and ten fruit descriptors; eight F3 populations, resulting from crossing the accessions 134 (P-9) and 77.1 (P-10), and five additional control populations: P-9, P-10, P-11, P-12 and P-13. There was an agreement between the two multivariate techniques in terms of distance between populations. Fruit descriptors contributed most to the genetic divergence, separating the populations used as control (P-11, P-12 and P-13) from the others. This separation is due to the uniformity of these populations in terms of fruit size and weight.
Key words: Canonical variables, morphoagronomic descriptors, genetic resources, genetic variability.
Abbreviation: PH, plant height; CD, canopy diameter; FFH, first fork height; SD, stem diameter; LL, leaf length; LW, leaf width; FL, fruit length; FLD, fruit largest diameter; FSD, fruit smallest diameter; PL, peduncle length; PT, pericarp thickness; PCL, placenta length; FL/FLD, the ratio between fruit length and fruit largest diameter; FW, fruit weight; DMC, dry matter content; NSF, the number of seeds per fruit; MANOVA, multivariate analysis of variance; Can1, first canonical variable; Can2, Second canonical variable.INTRODUCTION
Pepper belongs to the genus Capsicum and features wide morphological variability present in plants, flowers and fruits. This variability in fruit is portrayed by the differences in colors, shapes, sizes and flavors. The variability present in plant architecture gives the pepper high potential for use as ornamental plants because this sector of the market prefers short plants with colorful and erect fruits, as well as plants resistant to diseases, pests and abiotic stress (Carvalho and Bianchetti, 2007; Rêgo et al., 2009).
Despite this potential, the Capsicum improvement in Brazil is still not compatible with the relevance of pepper in the production chain (Reifschneider, 2000). However, this scenario is changing, considering the concern of breeders to develop cultivars with main focus on the fruit characteristics such as size, shape, capsaicin content, color, firmness, vitamin content and uniformity (Luz, 2007).
Thus, the study of genetic diversity is critical to the understanding of the genetic variability in populations or genotypes kept in Active Germplasm Banks (AGB). As it is known, in general, the morphologically far are the parents to be used in breeding programs the greater the heterotic effect (Gonçalves et al,, 2008; Rêgo et al., 2011b; Sudré et al., 2005). However, some factors, such as lack of documentation, description and evaluation of collections of genetic materials, may hinder the use of AGB, which limits the action of breeders (Gepts, 2006).
The use of morphological descriptors has been a common way to qualitatively and quantitatively characterize the variability in AGBs. According to Sudré et al. (2010), characterization of domesticated Capsicum species is of great interest, particularly for the AGB, as the wide variability in species is not yet fully known and exploited. Gonçalves et al. (2008) emphasize the importance of characterization of AGBs, as it makes the variability between populations or accession available to researchers, which allows the selection of superior genotypes and enables the increase of frequency of favorable alleles. These genotypes can also be used in hybrid combinations of high heterosis for future use in selecting segregating generations (Rêgo et al., 2011a, 2012b).
To determine the genetic distance between individuals, a group of individuals or populations, biometric models are used. In general, these models are based on multivariate techniques which allow a combining multiple information from a set of characteristics. Several multivariate methods can be used on studies of genetic diversity, such as cluster analysis, principal components and canonical discriminant variables. Considerations of using these methods include what is the most appropriate to get a desired accuracy, interpretation of results and how the data were obtained (Cruz and Carneiro, 2006).
We aimed to study the genetic divergence among F3 populations of ornamental pepper through multivariate methods, as well as to determine the morphoagronomic descriptors that contributed most to the genetic divergence.
MATERIALS AND METHOD
The study was conducted in a greenhouse in the Northeastern Brazil. Thirteen populations of ornamental pepper belonging to the Active Germplasm Bank (AGB) of the Laboratory of Plant Biotechnology of the Federal University of Paraíba were used. These populations consisted of eight F3 populations: F21 (P-1), F24 (P-2), F25 (P-3), F27 (P-4), F29 (P-5), F210 (P-6), F211 (P-7) and F231 (P-8), originated from crossing the accessions 134 (P-9) and 77.2 (P-10), and five populations used as additional control, accessions 134 (P-9), 77.2 (P-10), 10.1 (P-11), 10.2 (P-12) and 10.3 (P-13).
The seeds were sown in polystyrene trays with 200 cells filled in with commercial substrate Plantmax HT®. Thirty-five days after sowing, when the seedlings had three true leaves they were transplanted to plastic pots with volume capacity of 800 mL containing the same commercial substrate. The plants were watered daily, on alternate days with nutrient solution consisting of the following composition (g/1000 L): 1000 g of calcium nitrate; 1250 g of potassium nitrate; 250 g of MKP; 500 g of magnesium sulfate; 1.5 g of boric acid; 25 g of quelatec AZ; 25 g of ultraferro; 110 g of potassium chloride and 150 g of potassium sulphate. Phytosanitary treatment was carried out when necessary, throughout the growing cycle, in order to minimize the damage caused by pests and diseases.
The experimental design was completely randomized design. The experimental unit consisted of one plant per pot. Fifty plants of each of the eight F3 populations were assessed, as well as ten plants of the additional controls.
The morphoagronomic characterization was performed in accordance with the recommendations of the Capsicum descriptors proposed by IPGRI (1995). Sixteen morphological descriptors (6 of vegetatives and 10 of reprodutives) were used: PH (plant height), CD (canopy diameter), FFH (first fork height), SD (stem diameter), LL (leaf length), LW (leaf width), FL (fruit length), FLD (fruit largest diameter), FSD (fruit smallest diameter), PL (peduncle length), PT (pericarp thickness), PCL (placenta length), the ratio FL/FLD, FW (fruit weight), DMC (dry matter content), and NSF (the number of seeds per fruit), in two years: 2013 and 2014.
The data were subjected to multivariate analysis of variance according to a two two-way MANOVA model with the factors years and populations. The effect of the interaction between these factors was analyzed. To quantify the relative contribution of the descriptors to the genetic divergence we used the criterion of Singh (1981). The canonical discriminant variables were then constructed, whose average scores for each combination of factors were presented in two-dimensional plane through the biplot technique (Gabriel, 1971). From the loadings of the canonical variables, we evaluated the importance of each characteristic related to plant and fruit on the genetic divergence among populations. Furthermore, a cluster analysis was also performed via Ward algorithm, based on the squared generalized Mahalanobis distance. All analyzes were performed using R version 3.2.1 software (R Core Team, 2015).
RESULTS AND DISCUSSION
The result of the multivariate analysis of variance shows that there is interaction between the two factors (p < 0.01), indicating that the populations of ornamental pepper used in this experiment responded differently in each year.
Based on Singh (1981) criterion (Table 1), it can be seen that the descriptor that contributed most to the genetic divergence was FW, with 44.7 and 38.8% for the first and second year, respectively. This slight decrease in contribution is probably due to the different accu-mulations of fresh fruit weight. The higher contribution in the first year was also high for the FL (15.1%) and low (7.8%) in the second year. The fact that FL is a descriptor closely related to the production (FW) or is a production component explains the changing in the contribution of FL to discriminate the populations, and will affect the contribution of the FW (Table 1). This confirms the results obtained from the scores associated with the canonical variables, in which the contribution of the descriptors of fruit was decisive for determining the distance among populations. Sudré et al. (2006) reported that the most important descriptors were the length and the diameter of the fruit, and this is probably related to the number of species studied, since it is expected more differences in fruit morphology as the number of species increases.
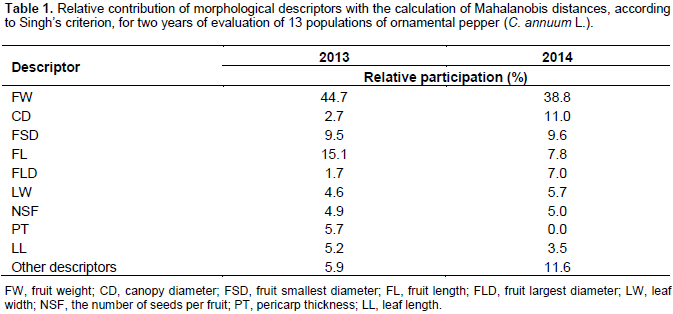
One can also highlight the participation of the variables FL (15.1%, year 1) and (7.8%, year 2) and FSD (9.5%, year 1) and (9.6% year 2). This decrease on FL is due to differences in the growth rate and greater uniformity of fruits observed in the second year, as well as the fact that populations used as control (P-11, P-12 and P-13) have heavier fruits, thus contributing to a smaller distance among these populations and the greater distance from them to other populations. This could be explained by the fact that the diversity contributed by cannopy diameter (CD) was low (2.7) in the first year and high (11.0) in the second year. The lower the use of assimilate for the vegetative growth (CD) might contribute to the higher the use for reproductive growth (FL and FW) and vise versa (Table 1).
The CD showed no effective contribution to the divergence among the populations, in both years. However, in the second year, it reached 11%. Probably, this major contribution is due to the fact that some populations naturally present different rates of develop-ment of plant canopy. According to Barbosa et al. (2002), Stommel and Bosland (2006), Rêgo et al. (2009) and Barroso et al. (2012), plants with compact size, lower height and smaller canopy diameter are of interest for selection of pepper plants with ornamental use purpose.
In Figures 1A and B, through the scores associated with the first two canonical variables (Can1 and Can2), the populations seem dispersed, although some form coherent clusters. The variability retained at the first canonical variable (Can1) was 58.4%, in the first year. The variables that contributed most to this canonical variable, in descending order of importance, were: FW, FLD, FL, FSD, PCL, LL and LW.
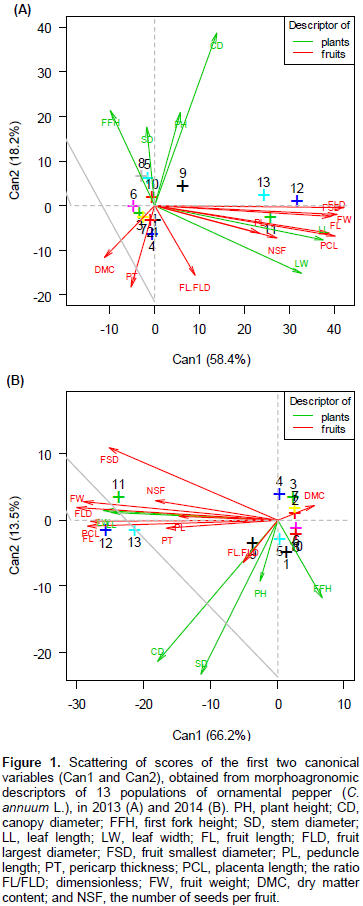
The canonical discriminant analysis showed that, in general, the variables associated with fruit (FW, FLD, FSD, FL and PCL) contributed most to the distance between populations, both in the first (Figure 1A) and in second (Figure 1B) year. These results coincide with those obtained by Hand et al. (2011), who reported that the two variables that contributed most to the divergence were variables of fruit.
It is noteworthy to highlight the behavior of populations P-11, P-12 and P-13 (Figure 1). They presented fruits significantly greater than the others. The aforementioned fruit characteristics were also responsible for the proximity between the populations (P-8 and P-5) (P-2 and P-1) (P-3 and P-7), (P-7 and P -2), (P-4 and P-2), (P-4 and P-1) and (P-6 and P-3), which are thus clustered for presenting, in general, smaller fruits.
In the first year, the variability retained by the second canonical variable (Can2) was 18.2%, mainly due to the contribution of CD, FFH, PH, PT, SD, CD and LW (Figure 1A). Silva Neto et al. (2014), working with C. annuum, reported that the characteristics that contributed most to the diversity were: stem diameter, canopy diameter and the first fork height. It is also observed that the populations P-4, P-1, P-2, P-7, P-3 and P-11 showed lower values ​​for variables related to the size of plants, while populations P-5 and P-8 showed the highest values. According to Rêgo et al. (2011a), variables related to growth habit and harmony between plant canopy and vase size are crucial for the potential of pepper as ornamental plant. Thus, populations presenting lower contribution to that characteristic can be selected for cultivation in vase, whereas when they have higher values, should be discarded.
The first two canonical variables retained together 76.6% of the variability between populations (Figure 1A) and, then, it was possible to identify two clusters of F3 populations: Cluster 1: P-1, P-2, P-3 and P-7; Cluster 2: P-8 and P-5. The distance between these two clusters and the populations P-11, P-12 and P-13 is evident. According to Cruz and Regazzi (2004), the formation of clusters with intracluster homogeneity and heterogeneity intercluter is the starting point for a more thorough assessment thereof, for use in future breeding programs.
In the second year of evaluation (Figure 1B), the variability retained in Can1 increased to 66.2%, and the descriptors that contributed to the distancing of the populations were: FW, FLD, FL, PCL, LW and FSD. In relation to Can2, which retained 13.5% of the total variability, the variables that contributed most were: SD, CD, FFH, PH, FSD and FL/FLD (Figure 1B). It highlights the phenotypic proximity between P-2 and P-7, also found in the first year, and the proximity of P-6, P-8 and P-10. This last cluster was formed probably due to the higher contribution of the characteristics relating to the size of the plant, presented by the populations P-6 and P-10, higher than that observed in plants of P-8, in the second year.
The first two canonical variables retained together 79.7% of the variability among populations (Figure 1B). The relationship between the characteristics studied and the evaluation period (first and second year) decisively influenced the behavior of the populations, because it is observed that in the first year, the P-1 population had one of the lowest values ​​for the size of plant characteristics while in the second year, showed relatively larger plants. Figure 1A can be seen that the P-5 and P-8 populations showed a similar behavior to that seen in the first year of assessment. Nevertheless, in the second year, both had increased relative to the size characteristics. It is worth noting the behavior exhibited by the population P-2, P-3, P-4, P-7, P-9, P-12 and P-13, which did not change when evaluated in the first and second years. Probably this fact is related with genetic factor, since there were no drastic changes in environmental issues.
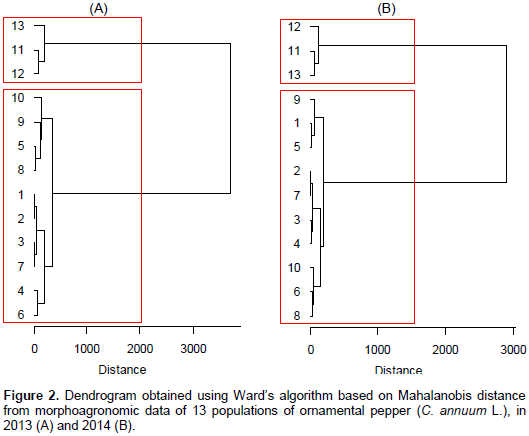
In Figure 2A and B we present the dendrograms obtained with Ward's algorithm, for the first and second year, respectively. In both years, only two clusters of populations were identified: (i) Populations from P-1 to P-10, and (ii) P-11, P-12 and P-13. These clustering were also identified by the canonical variables. Therefore, the canonical discriminant variables were more effective than the cluster analysis via Ward algorithm based on Mahalanobis distance. For instance, it can be seen through the canonical scores that P-6 diverges from P-9 (Figure 1A). This result was not verified using the cluster analysis (Figure 2A).
According to Bosland (1993), low-sized plants that produce small fruits, are considered promising for ornamental pepper agribusiness. One can also point out that plants with low-size and small canopy diameter are more harmonics. Furthermore, it is important to consider the relationship between plant architecture and vase size.
The plant height and canopy diameter should be 1.5 to 2 times larger than the size of the vase (Barbosa et al., 2002; Barroso et al., 2012).
CONCLUSIONS
It can be inferred that the populations are divergent, enabling the formation of different clusters, with the multivariate techniques - canonical discriminant variables and cluster analysis - presenting considerable agreement on the composition of the main clusters and on the contribution of the descriptors. Nonetheless, the canonical discriminant variables were more effective than the cluster analysis via Ward algorithm based on Mahalanobis distance. Fruit weight, fruit smallest diameter, fruit length and fruit largest diameter explained most of the variation among the populations, in the two years of evaluation. In breeding programs aimed at obtaining ornamental sized pepper, one should give importance to populations P-2, P-3, P-4 and P-7, with small values ​​for plant and fruit size, that is, it is recommended to carry out selection within these populations, in order to continue the ornamental pepper breeding program.
ABBREVIATION
PH, plant height; CD, canopy diameter; FFH, first fork height; SD, stem diameter; LL, leaf length; LW, leaf width; FL, fruit length; FLD, fruit largest diameter; FSD, fruit smallest diameter; PL, peduncle length; PT, pericarp thickness; PCL, placenta length; FL/FLD, the ratio between fruit length and fruit largest diameter; FW, fruit weight; DMC, dry matter content; NSF, the number of seeds per fruit; MANOVA, multivariate analysis of variance; Can1, first canonical variable; Can2, Second canonical variable.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interest.
ACKNOWLEDGMENTS
The authors are thankful to CNPq (Brazil) and the Instituto Federal Goiano (Brazil) for the financial support.
REFERENCES
|
Barbosa RI, Luz FJF, Nascimento Filho HR, Maduro CB (2002). Capsicum peppers cultivated in Roraima, Brazilian Amazonia. I. Domestic species. Acta Amaz. 32(2):177-177. |
|
|
Barroso PA, Rêgo ER, Rêgo MM, Nascimento KS, Nascimento NFF, Nascimento MF, Soares WS, Ferreira KTC, Otoni WC (2012). Analysis of segregating generation for components of seedling and plant height of pepper (Capsicum annuum L.) for medicinal and ornamental purposes. Acta Hortic. 953:269-275. |
|
|
Bosland PW (1993). Breeding for quality in Capsicum. Capsicum and Egplant Newsletter 12:25-31. |
|
|
Carvalho SIC, Bianchetti LB (2014). Pimenta (Capsicum spp.): botânica. Brasília: Embrapa Hortaliças, 2007. |
|
|
Cruz CD, Carneiro PCS (2006). Modelos biométricos aplicados ao melhoramento genético. Viçosa, MG: Universidade Federal de Viçosa. |
|
|
Cruz CD, Regazzi AJ, Carneiro PCS (2004). Modelos biométricos aplicados ao melhoramento genético. Viçosa: UFV. 480 p. |
|
|
Gabriel KR (1971). The biplot graphical display of matrices with application to principal component analysis. Biometrika 58(3):453-467. |
|
|
Gepts P (2006). Plant genetic resources conservation and utilization: the accomplishments and future of a societal insurance policy. Crop Sci. 46(5):2278-2296. |
|
|
Gonçalves LS, Rodrigues R, Amaral Junior AT, Karasawa M (2008). Comparison of multivariate statistical algorithms to cluster tomato heirloom accessions. Genet. Mol. Res. 7(4):1289-1297. |
|
|
IPGRI A, CATIE (1995). Descriptors for Capsicum (Capsicum spp.). Descriptores para Capsicum (Capsicum spp.). Rome: Int. Plant Genet. Resour. Institute (100p.)-illus.. ISBN, 700497568. |
|
|
Luz FJF (2007). Caracterizações morfológica e molecular de acessos de pimenta (Capsicum chinense Jacq.). 2007. 70 p. PhD Thesis (Agronomy) – Faculdade de Ciências Agrárias e Veterinária, Universidade Estadual Paulista, Jaboticabal-SP. |
|
|
R Core Team (2015). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available from: http://www.R-project.org/. Accessed on June 18, 2015. |
|
|
Rêgo ER, Finger FL, Nascimento MF, Barbosa LAB (2011b). Pimenteiras Ornamentais. In: Produção, Genética e Melhoramento de Pimentas (Capsicum spp.) (Rêgo ER, Finger FL, Rêgo MM (eds.). Imprima Recife pp. 205-223. |
|
|
Rêgo ER, Rêgo MM, Cruz CD, Finger FL, Casali VWD (2009). A diallel study of yield components and fruit quality in chilli pepper (Capsicum baccatum). Euphytica 168(2):275-287. |
|
|
Rêgo ER, Rêgo MM, Cruz CD, Finger FL, Casali VWD (2011a). Phenotypic diversity, correlation and importance of variables for fruit quality and yield traits in Brazilian peppers (Capsicum baccatum). Genet. Resour. Crop Evol. 58(6):909-918. |
|
|
Rêgo ER, Rêgo MM, Silva DF, Cortez RM, Sapucay MJLC, Silva DR, Silva Junior SJ (2009). Selection for leaf and plant size and longevity of ornamental peppers (Capsicum spp.) grown in greenhouse condition. Acta Hortic. 829:371-375. |
|
|
Reifschneider FJB (2000). Capsicum: pimentas e pimentões no Brasil. Brasília, DF: EMBRAPA Hortaliças. |
|
|
Silva Neto JJ, Rêgo ER, Nascimento MF, Silva Filho VAL, Almeida Neto JX, Rêgo MM (2014). Variabilidade em população base de pimenteiras ornamentais (Capsicum annuum L.). Rev. Ceres 61(1):84-89. |
|
|
Singh D (1981). The relative importance of characters affecting genetic divergence. Indian J. Genet. Plant Breed. 41(2):237-245. |
|
|
Stommel JR, Bosland PW (2006). Ornamental pepper, Capsicum annuum, In: Anderson, NO (ed). Flower breending and genetics: Issues, challenges and opportunities for the 21st Century. Springer, Dordrecht. The Netherlands. pp. 561-599. |
|
|
Sudré CP, Cruz CD, Rodrigues R, Riva EM, Amaral Júnior AT, Silva DJH, Pereira TNS (2006). Variáveis multicategóricas na determinação da divergência genética entre acessos de pimenta e pimentão. Hortic. Bras. 24(1):88-93. |
|
|
Sudré CP, Gonçalves LSA, Rodrigues R, Amaral Júnior AT, Riva-Souza EM, Bento CS (2010). Genetic variability in domesticated Capsicum spp as assessed by morphological and agronomic data in mixed statistical analysis. Genet. Mol. Res. 9(1):283-294. |
|
|
Sudré CP, Rodrigues R, Riva EM, Karasawa M (2005). Divergência genética entre acessos de pimenta e pimentão utilizando técnicas multivariadas. Hortic. Bras. 23(1):22-27. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


