ABSTRACT
Common bean is grown by smallholder farmers in Malawi who produce low yield due to drought constrains. Two experiments comprising of two different bean growth habits (IIa and IVa) were conducted at Chitedze Research Station in Malawi, to assess eleven common bean genotypes for yield and yield components under drought conditions. Drought reduced significantly number of pods plant-1, number of seeds pod-1 and consequently seed yield in both growth habits. Within growth habit IIa, the most productive genotype was BCB 2, that gave highest yield in both irrigated and drought stress conditions, also presented a higher yield percentage reduction (51.13%) after Sugar 131 (53.11%) implying highest drought susceptibility. On the other hand, VTTT 923/10-3 (growth habit IIa) had the highest drought tolerance (32.40%). Under growth habit IVa, genotype MAC 109 was found to be the most drought tolerant (31.43%) under drought and 12D/2 highest drought susceptibility (50.92%). The results suggest that selection for genotypes with higher number of pods plant-1, number of seeds pod-1 and low yield percentage reduction might improve grain yield under drought stressed condition.
Key words: Phaseolus vulgaris L., drought, growth habits, yield components.
Common bean (Phaseolus vulgaris L.) is an important crop with several uses; increasing food and reducing poverty, providing health and nutritional security, stability and provides a stable and lucrative source of income for many rural households (FAO, 2014) and also enhancing ecosystem resilience (Beebe, 2001). It is estimated that two-thirds of common bean production in the world occur under drought conditions (Beebe, 2008). Drought is the second most important factor in yield reduction after diseases (Rao, 2014). Malawi depends on agriculture where in every four years; farmers are experiencing crop losses due to drought (Kambewa, 2003). Beebe et al. (2011) reported that occasional severe droughts affecting Malawi are often associated with El Niño weather events. Farmers relying on rain fed agriculture are no more producing the required quantities from their fields even providing all the necessary crop inputs. While are challenges with production and productivity, the demand for food continues to increase year by year. There is need to produce more due to the continued population increase and the demand for food.
According to World Bank (2014), the Malawian population was at around 17 million and the World population was pegged at around 7.2 billion in the same year from around 6.1 billion in the year 2000, which means that the growth is significant year by year.
Climate change has resulted in change in food production patterns due to higher temperatures, increase carbon dioxide in the atmosphere, change of precipitation patterns, and increased vulnerability of the landless and the poor. A number of indirect techniques have been used for the evaluation of drought tolerance; however, seed yield is the most consistent indicator because it represents the harvestable product White et al. (1992) reported that the understanding of the relationship between yield and its components is important for making the best use of these relationships in breeding and selection.
The need for climate change adaptation measures has become more necessary to ensure farmers continue to produce for their own food security and that of the growing population (Chirwa, 2007). One such adaptation measure is to produce tolerant genotypes which can survive, yield better and adapt in different environments and to develop strategies to cope with cost-effective drought management techniques particularly for the poor smallholdes farmers who cannot afford to irrigate (Cavalieri, 2011).
Thus, a selected number of bean genotypes which were developed at International Center of Tropical Agriculture (CIAT) and at Lilongwe University of Agriculture and Natural Resources (LUANAR), with different growth habits were studied in a translucent plastic screening house at Chitedze Research Station. The main objective was to evaluate yield and yield components performance under drought condition.
The study was conducted at Chitedze Research Station which lies 1100 masl, latitude 13°85′S and longitude 33°38′E. Chitedze has a mean annual temperature of 200C. Maximum temperatures are more than 24°C in November and lowest below 16°C in July. The station receives a mean annual rainfall of 892 mm, 85% of which falls between November and March. A pot experiment was used because it is easy to have uniform management of treatments in terms of soil fertility and distribution of water.
Germplasm description
Eleven common beans genotypes with two different growth habits (types IIa and IVa) were used in the experiment. Types IIa have determinate growth with stem and branches ending in a reproductive guide. Types IVa are indeterminate with excessively long stems and branches making them weak and ending in vegetative long stems (Singh, 1995).
Experimental design and treatments
The experimental design was split-splot, laid out on completely randomized design (CRD). Two levels of water conditions (irrigated and drought) were the main plots and 11 genotypes namely : BCB 2 (Bunda 2), DRK 57, Sugar 131, VTTT 923/10-3, VTTT 924/4-4, VTTT 925/9-1-2, 12D/2, CIM-Climb 01-03-40, DC 86-263, Kanzama and MAC 109 were the sub-plots, making a total of 22 treatments. Each treatment (experimental unit) was composed of four pots, making a total of 88 pots per replicate.
Experimental management
Plants were grown from March to July in 5 litters plastic pots (17.7cm high × 21.2 cm diameter) carefully packed with 4.8 kg of sand clay loam soil. The whole trial was irrigated to field capacity one day before sowing. Four uniform seeds were sown 5cm deep in each pot and then thinned to three plants at two leaf stage. The pots were watered at two-day intervals to replace the water lost by evapotranspiration, keeping the pot soil moisture at 80% of field capacity. Watermark model 200SS-15 soil moisture sensor with cable was used for soil moisture measurement. A compound fertilizer (23% N: 21% P2O5: 0% K2O +4% S) was applied at a rate of 30 kgha-1 during the trifoliate leaf of the crop growth stage. Each pot was supplied with 0.64 g for fertilizer as a basal dressing, aiming to improve plant vigor. Hand weeding was done when necessary. Drought was imposed when 50% of the genotypes in each growth habit had flowered.
Data collection
Parameters such as: soil data, days to 50% flowering, days to 90% physiological maturity, number of pods plant -1, number of seed plant -1, pod length and seed yield were recorded. Percent reduction (change) in parameters measured under water-stress was derived from the difference in parameters’ values between non- stress and stress conditions (Basal et al. 2006) as follows:
Soil data
Soil was analyzed for physical and chemical properties in a soil laboratory at Lilongwe University and Natural Resources (LUANAR) for nitrogen, phosphorus, pH, soil texture and organic matter, and at Agriculture extension Research (ARET) soil laboratory for Potassium, Calcium, iron and zinc. The methods used to analyze the soil were: Dispersal and hydrometric readings, Anderson and Ingram (1993), pH Extractable method Maclean (1982) and total N with Micro-Kjedahl digestion distillation Bremner and Mulvanay (1982). Potassium and phosphorus were determined using Mehlich
three extraction methods (Chilimba et al., 2011). The soil had the following characteristics: pH, 5.35; OM%, 3.8; N%, 0.14; P (ppm), 55.4; K (meq100 g-1), 0.35; Mg (meq100 g-1), 0.017; Fe (ppm), 8.2; Cu (ppm), 2.0, Zn (ppm), 4.0; and Mn (ppm), 8.47.
Statistical data analysis
The statistic model used in the experiment was:
Yijk = m + Gi + Wj + (GW)i + (GW)j +Eij
Where: Yijk = the response of ith genotype within jth water regime in the kth replicate (k= 1, 2), m = the overall mean, Gi = the fixed effect of ith genotype (i = 1, 2, 3….11), Wj= the fixed effect of jth water regime (j= 1, 2), Gwij = fixed effect of interaction of genotype x water regime, Eij = random deviation of the kth replicate from the average of genotype x water regime.
Analysis of variance (ANOVA) was conducted using General statistics (GenStat 17th edition) to test for differences among genotypes, water conditions and interactions between genotypes and water conditions effects (Montgomery, 2001). Treatment means were separated using Fisher’s protect Least Significant Difference (LSD) test at P<0.05.
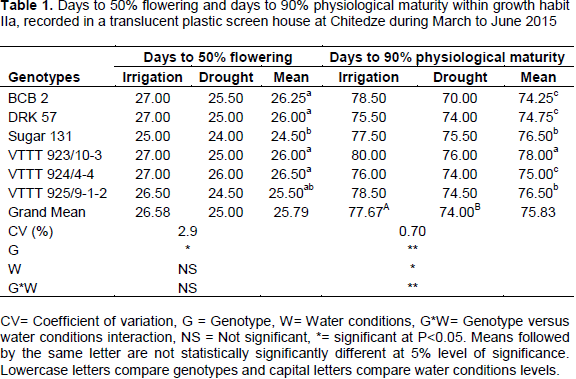
Phenological data within growth habit IIa
There was no significant difference (P>0.05) on days to 50% flowering due to water stress but there was significant differences among genotypes (Table 1). The earliest flowering genotype was noted in Sugar 131 (25.00 days) followed by VTTT 925/9-1-2 (26.00 days) and delayed flowering (27.00) was found in the genotype BCB2, DRK 57, VTTT 923/10-3, and VTTT 924/4-4. According to Schmalenbach et al. (2014) early flowering genotype is a common drought escape strategy that ensures plant survival under severe water deficit; still, early flowering shortens the time available for carbon assimilation during vegetative development and, thus, possibly results in yield reduction. The effect of water condition and genotypes interaction at 50% flowering were not significant (P>0.05) among the genotypes. The results from growth habit IIa showed significant differences between number of days to 90% physiological maturity in different bean genotypes. BCB 2, DRK 57 and VTTT 924/4-4 were the earliest maturing bean genotypes in 70.0, 74.0 and 74.0 days, respectively. Genotypes which have traits of shorter maturity normally escape terminal drought compared to longer days to maturing.
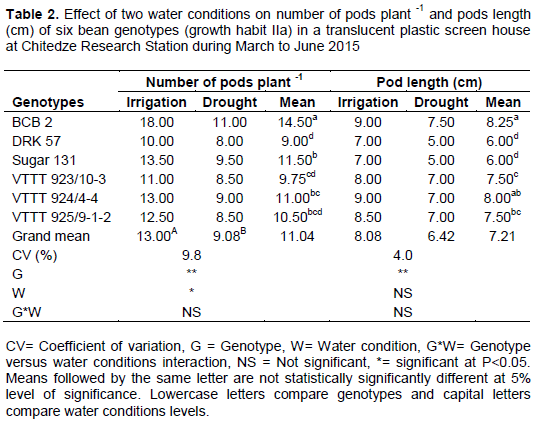
Effect of drought on yield components of common bean genotypes within growth habit IIa
The effect of drought on number of pods plant-1 and pod length was significant (P<0.01) different for genotypes and water conditions effect (Table 2). Results indicated that production of pods was less under drought condition as compared to irrigated condition in all genotypes. However, BCB2 had higher number of pods (18.00) followed by Sugar 131 (13.50) and the lowest number of pods were observed on DRK 57 (8.00). A similar result was also reported by Beebe et al. (2012) in common bean genotypes. This finding is also in agreement with Nielsen and Nelson (1998) who observed a reduction in the number of pods in plants subjected to drought; that effect could have been due to ovule or pollen abortion (Kokubun et al. 2001) and also due to increase of ethylene (ABA) production when plants are subjected to drought causing flowers abortion. The mean number of pods plant-1 was reduced by 30% and number of seeds plant-1 by 21%, these suggest that reduction of seed yield in drought conditions is mainly due to number of pods plant-1. The reduction of number of pods under drought in this study was probably due to limited assimilate supply under drought condition, as reported by Leport et al. (2006) in legumes, including common bean varieties.
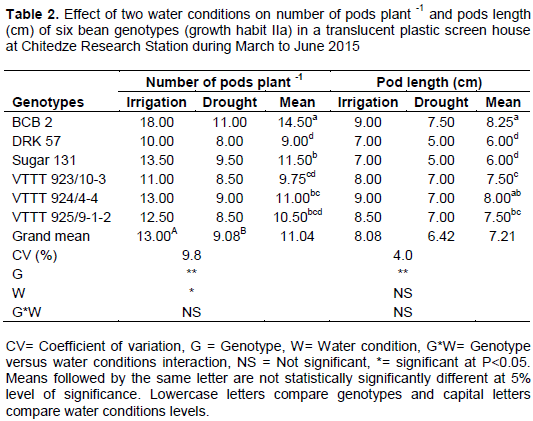
The overall means of pod length under drought was (6.42 cm) compared with (8.08 cm) under irrigated condition. Higher pod length was observed in BCB 2 (9.00 cm), followed by VTTT 925/9-1-2 (8.50 cm). Sugar 131 and DRK 57 had lowest pod length (7.00 cm). Acquaah (2007), report that when the genetic materials are different, there is also huge variability within them. Hence, the results in pod length on this study are in accordance to (Acquaah 2007).
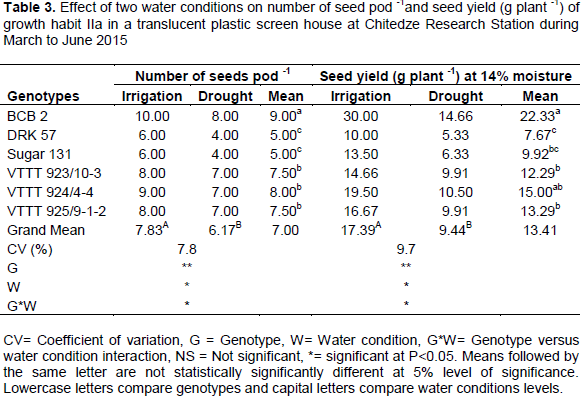
Analysis of variance indicate that number of seeds pod-1 and seed yield (g plant-1) were significantly (P<0.01) affected by the water condition. The interaction between genotypes and water condition on number of seeds pod-1 and seed yield (g plant-1) were also significant (P<0.05), suggesting a great amount of variability for drought tolerance in bean genotypes under study. Results are presented in Table 3.
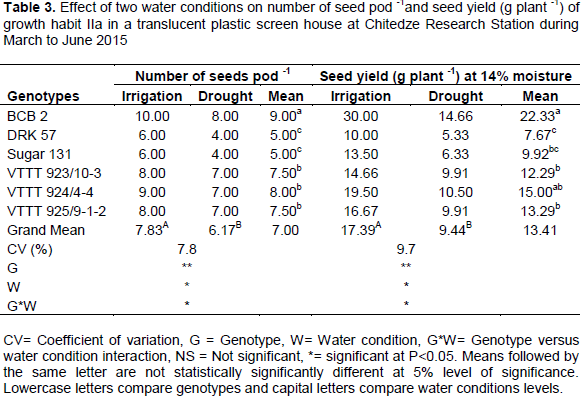
The overall means of number of seed pod-1 under drought was (6.17) compared with (7.83) under irrigated condition resulting in a 21.58% reduction. Average seed yields were 9.44 and 17.39 g plant-1 under drought and irrigated conditions, respectively. The percentage reduction was 45.72%. However, genotype VTTT923/10-3 had the highest drought tolerance (32.40%), followed by VTTT 925/9-1-2 (40.55%), VTTT 924/4-4 (46.15%), DRK 57 (46.70%). Both BCB2 (51.13%) and Sugar 131 (53.11%), had more than 50% reduction. Genotype BCB 2 showed second highest drought susceptibility level, after Sugar 131, though it produced the highest seed yield under drought condition. The highest seed yield of BCB2 may be attributed to high number of pods plant-1 and number of seeds pod-1. Furthermore, Rosales-Serna et al. (2004), also reported that drought resistant genotypes that display high yield under stress are more efficient in photoassimilate remobilization and this difference in seed yield among common bean cultivars under drought can be associated with physiological and
biochemical responses, such as tissue water retention, osmotic adjustment, integrity of membrane system, protease activity and stomata adjustment (Lizana et al.,2006). Genotypes and water conditions interaction had showed significant differences (P<0.05), implying that drought had decreased seed yield in all genotypes.
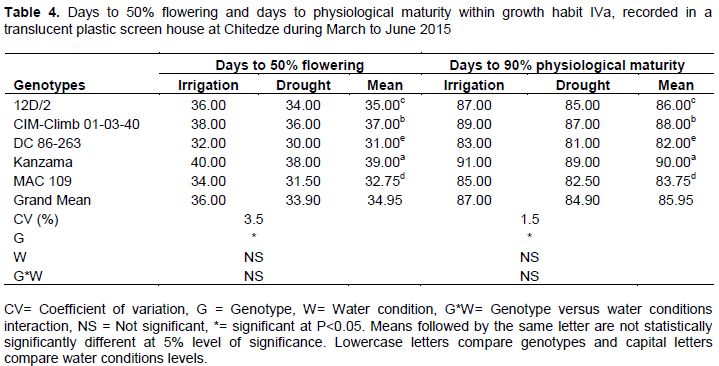
Phenological data within growth habit IVa
There was no significant difference (P>0.05) in days to 50 % flowering and 90% physiological maturity due to water stress condition (Table 4).
The earliest flowering genotype was noted in DC 86-263 (30.00 days) followed by MAC 109 (31.50 days) and delay flowering (38.00 days) was found in the genotype Kanzama. Early flowering indicates short life cycle and is considered a positive character for improvement of genotypes. Results under this study are in agreement with Oladosu et al. (2014) in a similar work on rice. The same scenario was observed on days to physiology maturity (Table 4).
Effect of drought on yield components of common beans genotypes within growth habit IVa
Analyses of variance had shown that number of pods plant-1 and pod length (cm) were significantly affected water condition. Genotypes and water condition interaction did not affect pod length (Table 5). However, interaction between genotypes and watering regimes was significant (P<0.05). Genotype 12D/2 had more number of pods plant-1 (11.00), followed by CIM-Climb 01-03-40 (10.50), Kanzama and DC 86-263 (10.00), respectively. MAC 109 genotype had less number of pods plant-1 (9.00).
Graham and Ranalli (1997) found that drought results in many phenotypic changes in the plant development including reducing number of pods-1, number of seed plant-1 and pod lenght. Under drought conditions, Kanzama, 12D/2 and CIM-Climb 01-03-40 had higher pod length with means of 8.50, 8.00 and 7.50 cm, respectively. DC 86-263 had lowest pod length (7.00 cm).
There were significant differences (P<0.05) for number of seeds pod-1 and seed yield (g plant-1) Results are presented in Table 6, suggesting that indirect selection for this trait is possible. Number of seeds pod-1 were different under irrigation condition, but not under drought, meaning that drought did not affect the number of seeds pod-1. However, genotypes 12D/2 (9.50), CIM-Climb 01-03-40 (9.00) had more seeds. Kanzama had least seeds pod-1 (5.00).
Drought had reduced yield by 39.29% (Table 7) under growth habit IVa. However, genotype MAC 109 had the highest drought tolerance (31.43%), DC 86-263 (32.78%), Kanzama (35.07%) and CIM-Climb 01-03-40 (35.50%). The genotype 12D/2 has been found to be most drought susceptible (50.93%).
Comparison between group habit IIa and IVa
In general, growth habit IIa had produced less yield and yield components compared with IVa. Yield and yield components are effective traits used in breeding programs. From this study, the lowest number of pods plant-1 was DRK 57 (10.00 and 8.00) under irrigated and drought conditions, respectively. DRK 57 and Sugar 131 had least number of seeds pod-1 (6.00 and 4.00) under both conditions, respectively. The same genotypes had least pod length (7.00 and 5.00 cm). In terms of seed yield (g plant -1) DRK 57 had least yield (10.00 and 5.33g plant -1) under irrigated and drought condition, respectively. The highest on this group was BCB2 genotype with the follow values: number of pods (18.00 and 11.00) under irrigated and drought condition; number of seeds pod -1 (10.00 and 8.00); pod length (9.00 and 7.50 cm) and seed yield of (30.00 and 14.00 g plant -1) under irrigated and drought conditions, respectively. On other hand, the lowest number of pods plant -1 in growth habit IVa was MAC 109 (10.50 and 9.00) under irrigated and drought conditions, respectively. Lowest number of seeds pod -1 was recorded on Kanzama (7.00 and 5.00) under irrigated and drought, respectively. Lowest pod length was from DC 86-263 (8.50 and 7.00 cm), under irrigated and drought conditions. Lowest seed yield was from Kanzama (12.83 and 8.33 g plant-1), under irrigated and drought conditions. Genotype 12D/2 had highest number of pods plant -1 (18.50 and 11.00) and number of seeds pod -1 (11.50 and 9.50) under irrigated and drought conditions. The highest pod length was Kanzama (10.00 and 8.50 cm), under irrigated and drought conditions, respectively. The highest yield was 12D/2 (35.50 and 17.42 g plant-1) under both water conditions. VTTT 923/10-3 (growth habit IIa), MAC 109 and DC 86-263 (growth habit IVa) were more tolerant to drought than the rest. The genotype that was most susceptible to drought was BCB2 under growth habit IIa and 12D/2 under growth habit IVa
Common bean is an important crop for food and reducing poverty for most of Malawian population. Development of drought tolerant genotypes is crucial important to produce crops which can yield better and adapt in different environments in Malawi. Drought has been found to, considerably, reduce the number of pods plant -1, number of seeds pod -1 and final yield. Yield performance under growth habit IVa was greater than in growth habit IIa. The findings indicate that selection for growth habit IVa would be useful to improving yield under drought condition.
The authors have not declared any conflict of interests.
We are grateful to Alliance for Green Revolution in Africa (AGRA) for having granted work and making financial resources available throughout all the study period. We wish to acknowledge the International Center of Tropical Agriculture (CIAT) for providing the bean germplasm used in this study. We also extend gratefulness to Lilongwe University of Agriculture and Natural Resources (LUANAR) for technical assistance and support.
REFERENCES
|
Acquaah G (2007). Principles of plant genetics and breeding. Blackwell Publishing, Oxford, UK.
|
|
|
|
Basal H, Demiral MA, Canavar O (2006). Shoot biomass production of converted race stocks of upland cotton Gossypium hirsutum L exposed to salt stress. Asian J. Plant Sci. 5:238-242.
Crossref
|
|
|
|
Beebe S, Rengifo J, Gaitan E, Duque MC, Tohme J (2001). Diversity and origin of Andean landraces of common bean. Crop Sci. 41:854-862.
Crossref
|
|
|
|
Beebe SE, Rao IM, Cajiao C, Grajales M (2008). Selection for drought resistance in common bean also improves yield in phosphorus limited and favorable environments. Crop Sci. 48:582-592.
Crossref
|
|
|
|
Beebe S, Ramirez J, Jarvis A, Rao IM, Mosquera G, Bueno JM, Blair W (2011). Genetic improvement of common beans and the challenges of climate change. In: S.S. Yadav, R.J. Redden, J.L. Hatfield, H. Lotze- Campen and AE Hall (eds.) Crop Adaptation to Climate Change. John Wiley and Sons, Ltd., Published by Blackwell Publishing Ltd. pp. 356-369.
Crossref
|
|
|
|
Beebe SE, Rao IM, Blair MW, Acosta-Gallegos JA (2013) .Phenotyping common beans for adaptation to drought. Front. Plant Physiol. 4(35):1-20.
Crossref
|
|
|
|
Bremner JM, Mulvaney CS (1982): Methods of soil analysis, part 2 chemical and microbiological properties pp. 595-624.
|
|
|
|
Cavalieri A, Merchant A, Van Volkenburgh E (2011). Why not beans? Funct. Plant Biol. 38:iii–vi.
Crossref
|
|
|
|
Chirwa R (2007). Experiences in implementing the Bean Seed Strategy in Malawi. J. Sustain. Agric. 29(27):43-69.
Crossref
|
|
|
|
Chilimba ADC, Young SD, Black CR, Rogerson KB, Ander EL, Watts M, Lammel J, Broadley MR (2011). "Maize grain and soil surveys reveal suboptimal dietary selenium Intake is widespread in Malawi," Sci. Reports 1:72.
Crossref
|
|
|
|
FAO (2014). Estadísticas de fríjol seco. In:
|
|
|
|
Graham PH, Ranalli P (1997). Common bean (Phaseolus vulgaris L.). Field Crops Res. 53:131-146.
Crossref
|
|
|
|
Kambewa P (2005). A Review of Safety Net Programmes for 2003–2004 (Republic of Malawi: Lilongwe, 2005).
|
|
|
|
Kokubun M, Shimada S, Takahashi M (2001). Flower abortion caused by preanthesis water deficit is not attributed to impairment of pollen in soybean. Crop Sci. 41:1517-1521.
Crossref
|
|
|
|
Leport L, Turner NC, Davies SL, Siddique KHM (2006). Variation in pod production and abortion among chickpea cultivars under terminal drought. Eur. J. Agron. 24(3):236-246.
Crossref
|
|
|
|
Lizana C, Wentworth M, Martinez JP, Villegas D, Meneses R, Murchie EH, Pinto M (2006). Differential adaptation of two varieties of common bean to abiotic stress. I. Effects of drought on yield and photosynthesis. J. Exp. Bot. 57:685-697
Crossref
|
|
|
|
Mazuma EDL, Kisyombe CT, Kumwenda S, Maseko L, Kabvala E, Butawo E, Mviha P, Kapapa C (2004). Development of Integrated Management for Major Diseases in Common Beans.
|
|
|
|
Mkandawire ABC, Benati S, Pankomera D, Lupwayi NZ (1997). Drought tolerance in African common bean genotypes. In Adipala E, Tenywa, JS, Ogenga-Latigo, MW. Afr. Crop Sci. 3:239-246.
|
|
|
|
Montgomery DC (2001). Design and analysis of experiments. (5thed.). Hoboken: John Wiley & Sons, Inc.
|
|
|
|
Nielsen DC, Nelson N (1998) Black bean sensitivity to water stress at various growth stages. Crop Sci. 38:422-427.
Crossref
|
|
|
|
Oladosu Yusuff, Rafii MY, Norhani A, Mohammad AM, Rahim HA, Ghazali H, Mohammad AL, Isiaka K (2014). "Genetic Variability and Selection Criteria in Rice Mutant Lines as Revealed by Quantitative Traits". The Scientific World Journal.
|
|
|
|
Rao IM (2014). Advances in improving adaptation of common bean and Brachiaria forage grasses to abiotic stresses in the tropics. In: M. Pessarakli (ed). Handbook of Plant and Crop Physiology, Third Edition. pp. 847-889. CRC Press, Taylor and Francis Group, USA.
Crossref
|
|
|
|
Rosales MA, Campo OE, Rodríguez-Valentín R, Olvera-Carrillo Y, Acosta-Gallegos J, Covarrubias AA (2012). Physiological analysis of common bean (Phaseolus vulgaris L.) cultivars uncovers characteristics related to terminal drought resistance. Plant physiology and biochemistry: PPB / Sociétéfrançaise de physiologie végétale 56:24-34.
|
|
|
|
Singh SP (1995). Selection for water stress tolerance in interracial populations of common bean. Crop Sci. 35:118-124.
Crossref
|
|
|
|
Schmalenbach I, Zhang L, Reymond M, Gomez J (2014). The relationship between flowering time and growth response to drought in the Arabidopsis Landsberg erecta Antwerp-1 population. Front Plant Sci. 5:609.
Crossref
|
|
|
|
White JW, Castillo JA (1992). Evaluation of diverse shoot genotypes on selected root genotypes of common bean under soil water deficits. Crop Sci. 32:762-765.
Crossref
|