Full Length Research Paper

ABSTRACT
Ten accessions of Ricinus communis L. were grown in derived savannah agro-ecological zone for two cropping seasons under rain fed conditions. The work aimed at phenotypic characterization of R. communis for different qualitative and quantitative traits. The field characterization of the genotypes was done using randomized complete block design with three replications at the new site of Botanical garden, University of Nigeria, Nsukka. The different genotypes were randomly assigned to each of the ten plots within a block. Data collected on morphological and flowering attributes was subjected to statistical analysis. The results on phenotypic traits showed variations in fruit texture and stem color. Seedling emergence across the accessions was uniform in both years. The number of branches appeared stable within genotypes in years showing non-significant genotype by year mean square values. The number of leaves showed a significant variation (p≤0.05) among genotypes in 2013. There was a significant difference in the effects of years on the number of capsule and seed weight.
Key words: Castor, Ricinus communis, phenotypic traits.
INTRODUCTION
Castor (Ricinus communis L.), the single member of the African genus Ricinus, presents a wide variation regarding vegetative traits such as leaf and stem colors, number and size of leaf lobes and presence of wax. Castor bean plant occurs in dense stands and is frequently found along roadsides. It is both self- and cross-pollinated by wind, but controlled crossing studies suggested that out crossing is a frequent mode of reproduction (Meinders and Jones, 1995).
It originates from India and is cultivated in the tropical and sub-tropical climates of the world (Weiss, 2000). India is the world’s largest producer of castor seed and meets most of the global demand for castor oil; other major producers being China, Brazil, Ethiopia and Paraguay (FAO, 2013).
According to Weiss (2000), the castor bean plant varies greatly in its growth habit, colour of foliage, stems, seed size, colour and oil content, and varieties often bear little resemblance with each other. Colour differences in leaves, stems and inflorescences aid in the selection of horticultural and ornamental plants (Koutroubas et al., 1999). In its widespread naturalized state, castor is usually a fairly tall, mainly branched perennial but when cultivated commercially, it is short-lived, erect, little branched and treated as an annual crop (Ogunniyi, 2006).
Castor crop can be grown on a wide range of soils, provided they are fairly deep and well drained. Heavy clays, with poor drainage and marshy soils are unsuitable. The highly suitable soils for castor are deep, moderately fertile, with slightly acidic conditions (pH 5.0 – 6.5), well drained sandy loams. Excessively fertile soils are not desirable, as they favor excessive vegetative growth at the expense of seed yield (Weiss, 2000).
Castor flowers occur most periods of the year in dense terminal clusters with female flowers just above the male flowers. This species is clearly monoecious, with separate male and female flowers on the same individual. There are no petals and female flower consists of a little spiny ovary (which develops into the fruit or seed capsule) and a bright red structure with feathery branches that receives pollen from male flowers. Each male flower consists of a cluster of many stamens which literally smoke as they shed pollen in a gust of wind (Weiss, 2000).
The capsule is composed of three sections or carpels which split at maturity. Each carpel contains a single seed and as the carpel dries and spilt open, the seed is often ejected with considerable force (Mensah and Ochran, 2005). Castor seeds are nearly flattened and oval but differ in size and colour. The seeds may be white, black, brown or have several colours occurring as very attractive mottling on testa (Mensah and Ochran, 2005; Salunkhe and Desai, 1986). LI et al. (2008) described the seeds as shiny brown seeds with darker streaks or spots and resemble a blotted tick. A number of colours are usually found in castor seeds. The most striking ones are red, white and grey. Others are faint chocolate, deep chocolate and purple (Ikisan, 2000). The sizes of castor seed have been reported to vary (Weiss, 2000). The seed vary greatly in size, from 10 to about 250 mm long in giant types and from 5 to 16 mm in breadth. The 100 seed weight may vary from 10-100 g, averaging about 30 g in dwarfs (Li et al., 2008; Salunkhe and Desai, 1986). There is a great variation in seed sizes; the smallest seed noticed is 5.30 × 9.95 mm, while the largest is 13.50 × 20.95 mm. The common cultivated types have seed size ranging from 7.79 × 11.19 to 9.80 × 13.54 mm. The mean weight of castor seeds has been reported to vary from 10 - 100 g; for instance, seeds found in Nigeria and Kenya have been reported to weigh 69.3 and 59.2 g, respectively.
There are a very few documented evidences describing morphological diversity in castor. Earlier taxonomists and botanists studied morphological diversity with the purpose of classifying the genus Ricinus. Moshkin (1986) reported existence of diverse morphological variants in many parts of the world for plant height, branching, stem colour, leaf size, waxy coating, length, shape and compactness of raceme, pedicle length, size and shape of capsule and seed. Woodend (1993) described white, black and dark brown seed with varying mottling intensity, dark green and dark red colour stem, prostrate to columnar growth habit, weak-framed, robust and tree-like plant types among Zimbabwe collections.
The seeds of castor bean contain more than 45% oil and its oil is rich (80 to 90%) in an unusual hydroxyl fatty acid, ricinoleic acid. The oil is characterized by a high viscosity, owing to the hydrogen bonding of its hydroxyl group. It is rich in unique hydroxyl fatty acid. Ricinoleic acid is a major component in castor oil. Ricinoleic acid is widely used for its lubricating properties and medicinal purposes. It is widely used for its lubricating properties and medicinal purposes in industry. It is also used for manufacturing soaps, lubricants, hydraulic and brake fluids, paints, dyes, coatings, inks, cold resistant plastics, waxes and polishes, nylon, pharmaceuticals and perfumes (Duke, 1998).
Uncultivated wild and semi-wild castor plants are widespread not only in its centres of origin, but also outside. They represent the tremendous variability existing in the species. Castor had been adapted to diverse ecological niches. Therefore, ecological heterogeneity, stresses, natural selection and its interaction with other evolutionary forces including mutation, migration and genetic drift might have contributed greatly to genetic diversity according to circumstances in the natural niches. Studies on genetic diversity are necessary to elucidate and categorize the naturally existing variability. Genetic diversity in castor was assessed mostly by using agro-morphological traits and to some extent by molecular techniques. The vast worldwide castor collections reported were poorly studied and barely tapped for castor genetic improvement. The development of new cultivars with traits of interest and adaptation to specific microclimates is only possible when there is available knowledge on the extent of genetic diversity among the species (Gepts, 2004). Due to increased demand for castor bean in many countries, improvement of varieties is drawing attention from breeders (Sujatha et al., 2008). Success in breeding for high yield is limited by a low genetic variability and lack of geographically structured genetic populations, for productivity traits and sources of resistance to diseases and pests (Weiss, 2000; Hegde et al., 2003; Allan et al., 2008; Foster et al., 2010). It is therefore necessary to characterize the genetic diversity present in R. communis germplasm from different geographic regions to develop a genotyping scheme that links castor bean to a particular source, geographic region or batch (Hinckley, 2006). Information on diversity and genetic structure is essential in organizing accessions in germplasm collections because it allows effective maintenance of distant genotypes, thus reducing costs (Allan et al., 2008).
Although, the phenotypic traits have been extensively studied in castor in other parts of the world, such information is virtually unavailable for castor under Nigerian environmental conditions. Hence, the aim and objective of this work was to characterize R. communis genotypes by field evaluation for different phenotypic traits.
MATERIALS AND METHODS
The experimental design was randomized complete block design (RCBD) measuring 390 m2 with three replications. Each block measuring 3 by 4 m was subdivided into ten plots with 0.5 m spacing between plots. A space of 1 m was allowed between blocks with row spacing of 30 cm. The different genotypes were randomly assigned to each of the ten plots. Randomization was repeated for each block. Standard cultural practices were applied to all plots. The data collected were analyzed using RCBD format. The traits observed in this work were randomly selected. Statistically significant differences were detected using LSD at P = 0.05 levels of significance. The following quantitative traits were observed in the course of the work:
Days to emergence
The number of days it took a seedling to emerge was recorded. The data were randomly selected.
Days to 50% emergence
Emergence count started when the seeds sowed have emerged. 50% emergence is when half of the accessions sown have emerged.
Plant height (cm)
The height of castor plants started after four weeks and was done using meter rule and measuring tape. This was taken at four weeks (4 weeks) interval throughout the duration of the work (16 weeks).
Number of branches
The number of branches was taken into record after four weeks and these continued throughout the duration of the work at the interval of four weeks. These data was collected by counting.
Number of leaves
The number of leaves was counted at interval of four weeks.
Days to first flower
The number of days to flowering was counted from number of days after emergence to first flower.
Days to 50% flowering
The number of days in which 50% of the plants have flowered per plot was also recorded.
Days to maturity of castor bean plant
Days to maturity was calculated by counting the number of days from sowing date to the day when capsules colour changed from green to brown.
Number of capsules per plant
The number of capsules per plant was determined by dividing the total number of the capsule by the number of plants that emerged.
Seed weight
The seeds of each accession were weighed at the end of the experiment at the Department of Pure and Industrial Chemistry, University of Nigeria, Nsukka. The weighing machine used was mettle P1200.
Stem and leaf color
In order to ascertain the exact color of the stem and leaf of castor plant, the traits were subjected to eye estimation.
Data analysis
The data collected were analyzed using RCBD format. Statistically significant differences were detected using least significant difference (LSD) at P = 0.05 levels of significance.
RESULTS AND DISCUSSION
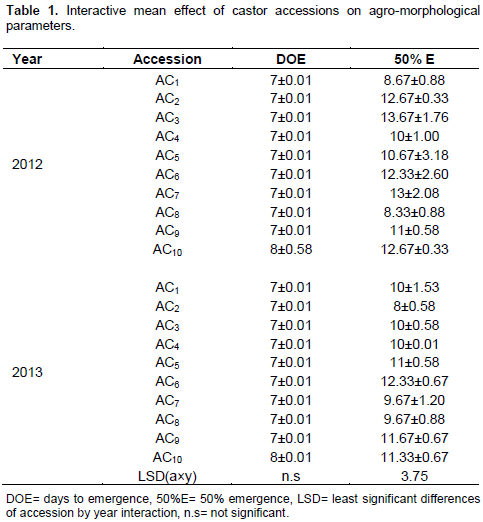
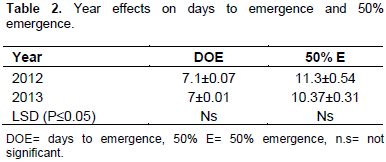
The data collected on the evaluation of castor accessions under rain fed conditions were analyzed and the means are reported. The phenotypic traits showed marked variations. The stem colour varied from light green to reddish. Most of the accessions had mean values of 7 days in both years on days to emergence. All the accessions had the lowest 50% emergence in 2012 evaluation season (Table 1). The value of number of days to 50% emergence ranged from 8 to 13.67. Accession 3 had the highest mean value of 13.67 on days to 50% emergence (Table 1). Ac 1 collected from Ibagwa-Aka started emerging 7 days after planting and within another two days (8.67), it attained 50% emergence. The year effects on days to emergence and 50% emergence was shown (Table 2). From the data presented, days to emergence and 50% emergence were statistically insignificant (Table 2). The lowest number of days to 50% emergence was recorded in 2012 evaluation season. In the second year, however, the gap between days to emergence and a number of days to 50% emergence widened, giving 7 to 12 days gap between days to emergence and number of days to 50% emergence. In the environmental condition of year two, accession 10 had the shortest emergence and could be selected in a breeding strategy to escape drought. The uniform emergence reported among the accessions aids the crop in avoiding drought and environmental stress. Uniformity in emergence recorded in this study could be attributed to earliness in sowing castor seed. Different sowing dates have profound influence on the germination of the seeds.
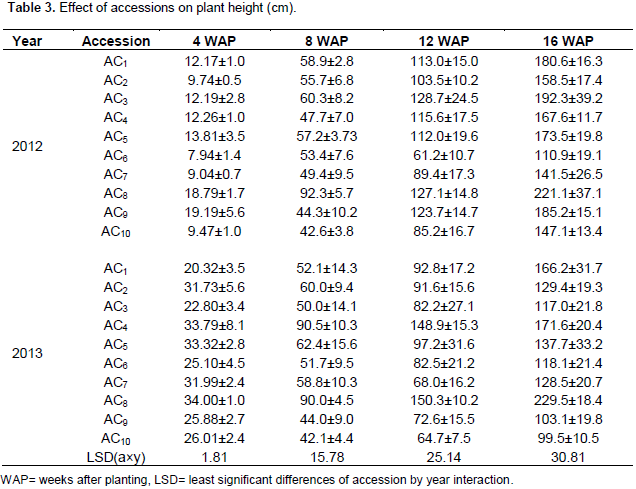
Variations exist within genotypes in a particular year and across the different growing seasons for plant height (Table 3). The lowest plant height was recorded in 2012 growing season. Accession 1 collected from Adani attained the plant height of 180.6 cm at 16 weeks after planting (WAP) in 2012 growing season. Similar effect was recorded in accession one in 2013 growing season at 16 weeks after planting (16WAP). This height of plant recorded is disadvantageous as it blocked other crop from sunlight thereby reducing their growth and photosynthetic activity in the leaves. The perennial growth habit of castor plants limits mechanical harvest because the plant grows very tall when the environmental conditions are desirable. Different temperature and moisture levels may have affected the plant height to some extent within the genetic potential of the plant. Overall plant height remained the same over years which imply the existence of dwarf and tall genotypes.
The number of branches had the highest value in 2012 growing season except at 4WAP (Figure 1). The number of branches at 4 weeks after planting (DAP) varied from 5 to 6 in 2012 and 5 to 7 in 2013. Ac 1 had the highest significant number of branches (7) in 2013. Accessions 7 and 10 had the same number of leaves (6) in 2013 (Figure 1). Ac 8 had significantly the highest value (15) in number of branches at 8WAP in 2012 (Figure 2). The number of branches at 8WAP ranged from 8 to 15 in 2012 and 6 to 8 in 2013. A similar number of branches were recorded in ac 2 in 2012 and ac 1 in 2013 (Figure 2). AC 6 had the highest number of branches in 2012 at 12WAP. The number of branches ranged from 9 – 16 in 2012 and 6 – 8 in 2013. Similar number of branches was attained in accessions 7 and 8 in 2013 while in 2012; accessions 8 and 9 had the same value (Figure 3). There was a significant variation in the number of branches among the accessions in sixteen weeks after planting (Figure 4). The number of branches at 16WAP ranged from 10 to 15 in 2012 and 8 to 11 in 2013. In 2013, accessions 3 and 9 had the similar value (8). Accessions 6 and 9 had the highest significant number of branches (15) in 2012 while accessions 5 and 10 had same value (11) (Figure 4).
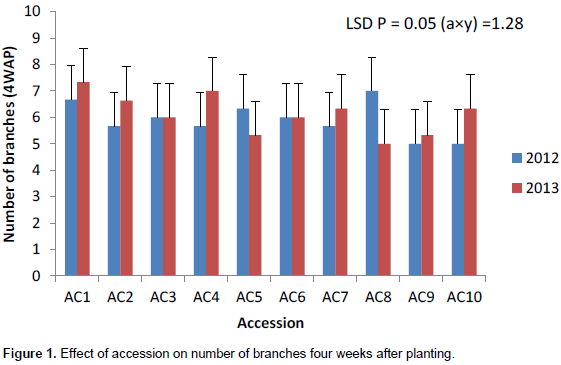
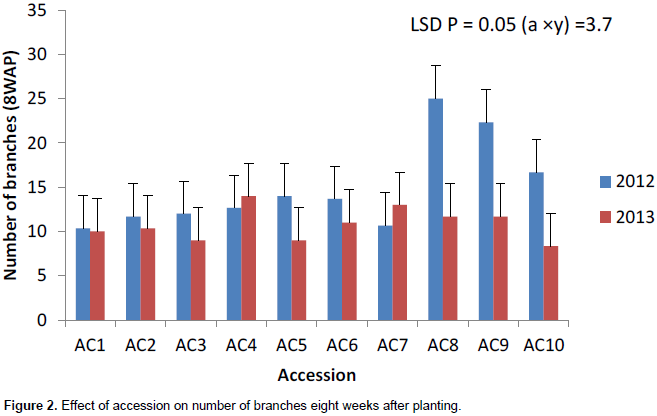
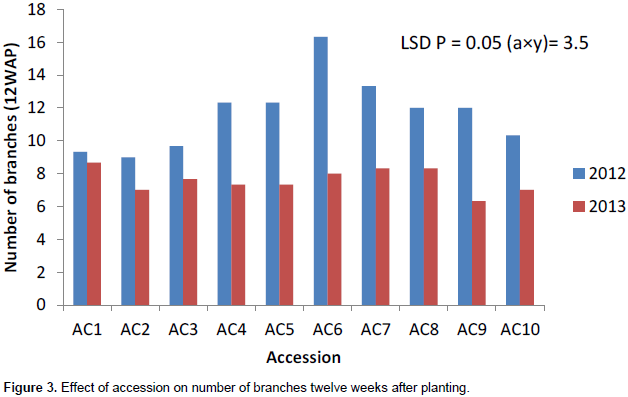
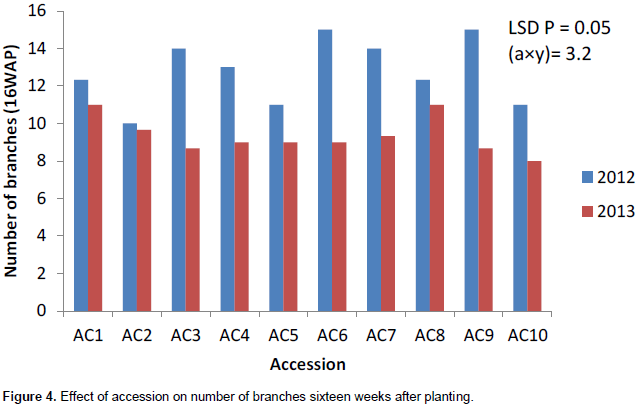
Generally, the number of branches performed better in 2012 growing season. The number of branches seemed stable within genotype in years but varied among genotypes. The number of branches had the highest value in 2012 while Accession 1 had the highest significant number of branches. The accessions seemed to be stable in year one but varied across the different growing seasons.
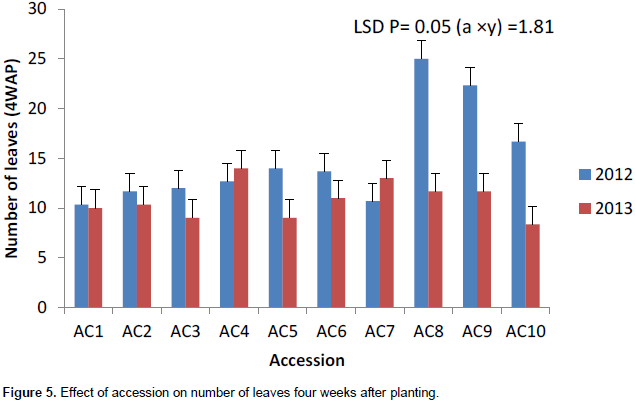
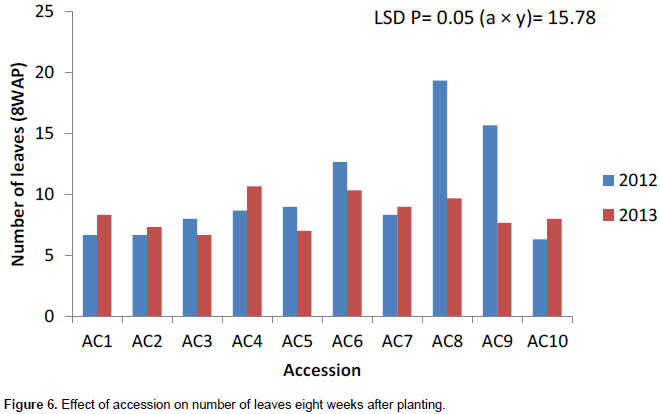
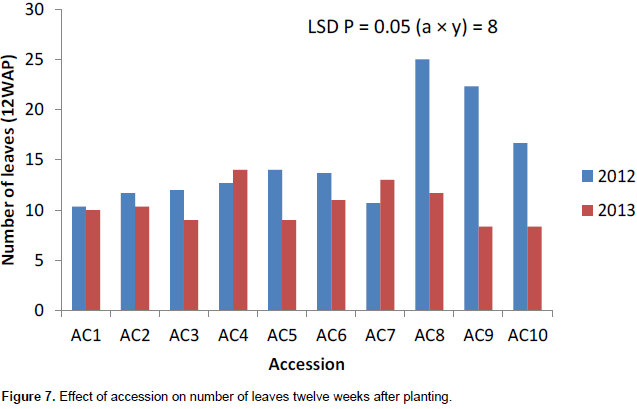
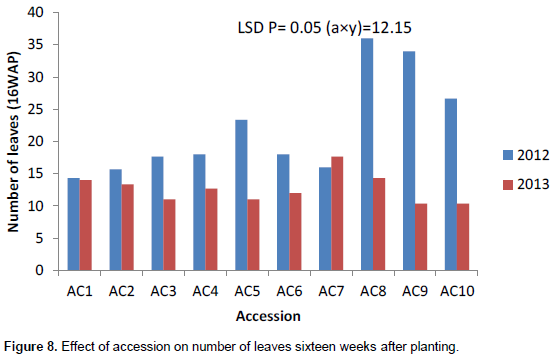
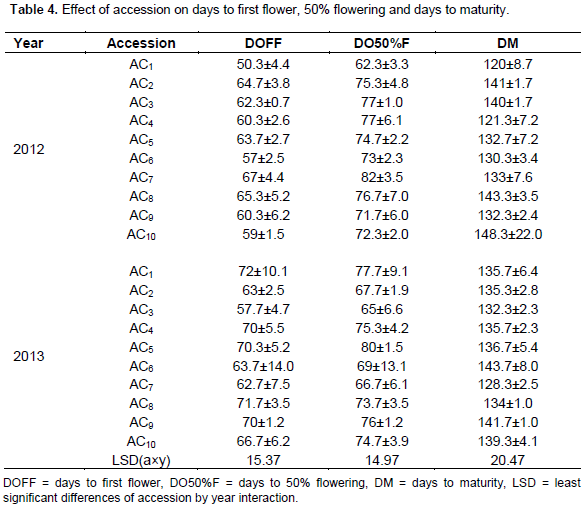
Accession 10 had the highest significant number of leaves (8) in 2013 at 4WAP (Figure 5). At 8, 12 and 16WAP, accession 8 had the highest significant number of leaves of 19, 25 and 36, respectively (Figures 6 to 8) in 2012. The number of leaves in both cropping season was attacked by Semi looper Achaea janata Linn. The smooth greyish-brown caterpillars feed on the leaves and sometimes cause wholesale devastation of the crop-damage to defoliation. Older larvae are voracious feeders and leave bare stems veins. This was controlled by hand picking of older larvae during early stages. The number of leaves was affected in year two by the tall plant height as it limits the amount of sunlight; thereby reducing its photosynthetic process. This also leads to significant difference in leaf area, plant height and higher seed yield. The use of green manure with inorganic fertilizer enhances the growth, morphological indices and yield of castor as compared to the application of either of them. Days to first flower had the highest value in 2013. The number of days to first flower was 72 days in accessions 4, 5 and 9 which had the same days to first flower. The value of this trait did not vary significantly (Table 4). Variation in days to 50% flowering was not significant in most of the accessions for the two years the accessions were evaluated but ac 7 had the highest significant days to first flowering in 2012 evaluation season. Stability in the manifestation of these traits was observed across years for the ten genotypes. Days to maturity of castor fruit did not vary (Table 4). All the genotypes had the highest number of days to maturity in 2013 evaluation season (Table 4). The flowers are borne in terminal panicle-like inflorescences of green or, in some varieties, shades of red monoecious flowers without petals. The male flowers are yellowish-green with prominent creamy stamens and are carried in ovoid spikes up to 15 centimeters long; the female flowers, borne at the tips of the spikes, have prominent red stigmas. Days to first flower had the highest value in 2013. Variation in 50% flowering was not significant in most of the accessions for the two years the genotypes were evaluated. Stability in the manifestation of these traits was observed across years for the ten genotypes.
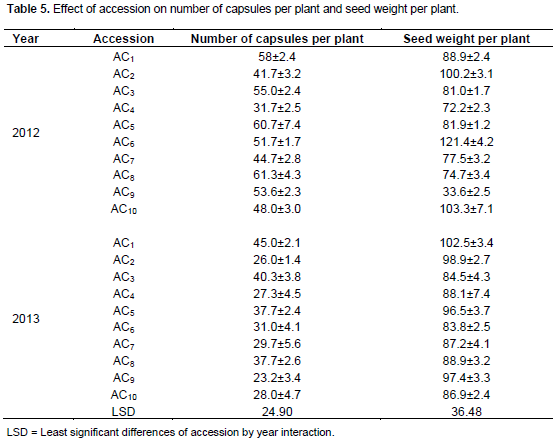
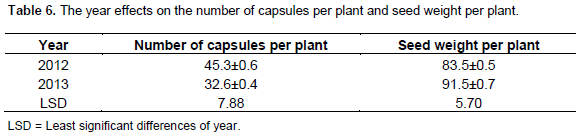
The effect of the genotype on the number of capsules per plant and seed weight was not statistically significant (Table 5). There was significant difference in the year effects on seed weight and number of capsules (Table 6). The highest number of days to maturity was recorded in accession 10 in 2012 and several accessions seemed to attain maturity within 130 to 136 days. Accession 2 produced the highest seed weight and all accessions produced heavier seeds in 2012 evaluation seed. Accessions 4 and 8 in 2013 had the same seed weight. Capsule weight is one of the important components contributing to the yield of castor bean crop. Accession 8 had the highest significant number of capsules per plant in 2012. For traits associated with yield, the spacing seems to have a significant effect on several of the study variables during both cropping seasons. The capsule weight during the cropping season decreased linearly. This could be attributed to competition for nutrients, reduced amount of light and less space available for plant development, thus less development of capsule.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Duke JA (1998). Ricinus communis from Purdue University New Crop Resource online Program. View Accessed on 16/5/2013 |
|
|
FAO (2013). The State of Food and Agriculture. United Nations, Rome 270 p. |
|
|
Foster JT, Allan GJ, Chan AP, Rabinowicz PD, Ravel J, Jackson PJ Keim P (2010). Single nucleotide polymorphisms for accessing genetic diversity in castor bean (Ricinus communis). Plant Biol. 10:1-11. |
|
|
Gepts P (2004). Crop domestication as a long-term selection experiment, In: Plant Breeding Reviews. John Wiley & Sons, Oxford, United Kingdom P 236. |
|
|
Hegde DM, Sujatha M, Singh NB (2003). Castor in India. Directorate of Oilseeds Research, Hyderabad, India. |
|
|
Hinckley AC (2006). Genotyping and bioforensics of Ricinus communis. Lawrence Livermore National Laboratory. Ph. D. Dissertation University of California P 170. |
|
|
Ikisan (2000). Castor Morphology and Growth. http://www.ikisan.com/ap-castor-morphology-and-growth.html Accessed on 20/6/2013. |
|
|
Koutroubas SD, Papakosta DK, Doitsinis A (1999). Adaptation and yielding ability of castor plant (Ricinus communis L.) genotypes in a Mediterranean climate. Eur. J. Agron. 11:227-237. |
|
|
Li FJ, Wang CL, Wang CF, Chen ZQ, Chen MH, Gao, LF (2008). Fatty acid composition of the castor bean seed of nine castor bean hybrids. China Oils Fats 33:62-64 |
|
|
Meinders HC, Jones MD (1995). Pollen shedding and dispersal in the castor plant Ricinus communis L. J. Agron. 4:206-209. |
|
|
Mensah B, Ochran R (2005). Physiochemical characteristics of castor oil from local wild castor plant in Ghana. Ghana J. Sci. 45:41-44. |
|
|
Moshkin VA (1986). Castor. Amerind Publishing Co. PVT Ltd, New Delhi. |
|
|
Ogunniyi DS (2006). Castor Oil: A vital industrial raw material. Bioresour. Technol. 97:1086-1091. |
|
|
Salunkhe DK, Desai BB (1986). Postharvest Biotechnology of oilseeds. CRC Press Boca Raton, Florida. 264 p. |
|
|
Sujatha M, Reddy TP, Mahasi MJ (2008).Role of biotechnological interventions in the improvement of castor bean (Ricinus communis L.) and Jatropha curcas L. Biotechnol. Adv. 26:424-435. |
|
|
Weiss EA (2000). Castor, Sesame and Safflower. Leonard Hill London. P 901. |
|
|
Woodend JJ (1993). Collecting and evaluation of indigenous perennial castor (Ricinus communis L.) in Zimbabwe. Pl. Genet. Resour. Newslett. 94/95:18-20. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


