Full Length Research Paper

ABSTRACT
Family agriculture is a rich germplasm source wherever it takes place; however, information on genetic variability of this type of culture in Brazil is scarce. Samples of melon (Cucumis melo L.) accessions grown by family agriculture were rescued and this study aimed at characterizing the genetic variability in one sample of these accessions so as to identify the melon subspecies and the corresponding varieties. Fifteen accessions and their S1 progenies were characterized (quantitative and qualitative descriptors) in two field experiments carried out in randomized blocks. Data on the parental and S1 generations were compared and it was possible to identify the subspecies Cucumis melo subsp. agrestis and their varieties C. melo var. makuwa and C. melo var. momordica, and the subspecies C. melo subsp. melo and its variety C.melo var. cantalupensis, although some sub-accessions remained unidentified. A total of 26 subaccessions were found. UPGMA grouping method showed a high genetic diversity among and within accessions and sub-accessions. Clusters were formed by the melon subspecies, although there were discrepancies. Nonetheless, there is indication of trait introgression from the two melon subspecies and their varieties in the material grown by the family farmers of the state of Maranhão.
Key words: Cucumis melo, botanical variety, traditional agriculture.
INTRODUCTION
Melon (Cucumis melo L.) is a vegetable belonging to the family Cucurbitaceae A. Juss, subfamily Cucurbitoidae with great economic importance in several parts of the world. It is originated in tropical Africa (Burger et al., 2010) although there are studies indicating that the genus Cucumis L. is originated in Asian-Australian (Renner et al., 2007; Schaefer et al., 2009; Sebastian et al., 2010). In Brazil, the Northeastern region is responsible for 94% of the national melon production (Agrianual, 2014) and the most frequently produced melon is the Yellow type (Cucumis melo var. inodorus), to which several cultivars and hybrids commonly recognized as AF-682, Tropical, AF-646, Gold Mine, Vereda, Goldex, and Jangada. In addition, there are other types that belong to the botanical variety cantalupensis: Piel de Sapo, Galia, Charentais, Cantaloupe, and Honey Dew (Costa, 2008). Since they are genetically improved products, these cultivars are uniform and have good plant and fruit traits despite some limitations regarding tolerance or resistance to biotic and abiotic stresses. Some breeding programs have been conducted at Embrapa units and at the Federal Rural University of the Semiarid Region (UFERSA); consequently, there is a demand for germplasm in order to progress further in the development of new cultivars (Aragão, 2015, personal communication).
On the other hand, there is a high variability between melon accessions grown by small farmers, mainly in the states of Piauí and Maranhão, from which, a sample was rescued and is stored in the Active Germplasm Bank of Cucurbitaceae of the Brazilian Northeast (BAG) (Silva et al., 2010) and this sample has more than 150 accessions. The species C. melo shows high polymorphism, mainly in fruit traits, such as size, shape, color, texture, and flavor, and is considered the most diverse species in the genus Cucumis (Bates and Robinson, 1995).
Cucurbit species were introduced in the Brazilian Northeast through different routes (Correa, 2010), and to date, have been dry farmed in small-sized agricultural establishments where farmers use their own seeds in the rainy season every year, thereby constituting traditional varieties or populations. Variability studies on the rescued cucurbits have fairly contemplated watermelon accessions (Citrullus lanatus (Thunb) Matsun & Nakai). However, few studies have been conducted with C. melo accessions. Neitzke et al. (2009) studied genetic variability in melon landraces in the South of Brazil, but, the authors do not allude to the subspecies or their varieties of the studied accessions. On the other hand, Torres Filho et al. (2009) characterized 42 accessions of melon plants grown by traditional agriculture in the Brazilian Northeast, and found a high variation among accessions, as well as different botanical varieties, following the classification by Munger and Robinson (1991). Furthermore, Aragão et al. (2013) evaluated the genetic diversity among melon accessions from traditional agriculture of the Brazilian Northeast using data from the morphological characterization conducted by Torres Filho et al. (2009), and they also performed molecular characterization. They observed a wide variability among accessions and different botanical varieties using the classification by Robinson and Decker-Walters (1997). Recently, a study was conducted on genetic diversity following the botanical classification by Pitrat et al. (2000), where a sample of melon accessions grown by traditional agriculture in the state of Maranhão (data not published). Despite its relatively small size, the sample was large enough to identify the occurrence of both melon subspecies (C. melo subsp. agrestis and C. melo subsp. melo) and some varieties (C. melo var. conomon, C. melo var. momordica, C. melo var. chandalak and C. melo var. cantalupensis). Nonetheless, there are over 100 accessions in the melon Germplasm Bank that have not been studied in depth, out of which approximately 80 accessions have also been collected from traditional agriculture, which emphasizes the need for furthering this research.
Hence, this study aimed at morphologically characterizing a second sample of melon accessions from the Cucurbitaceae Active Germplasm Bank of the Brazilian Northeast, which were collected in the state of Maranhão, using part of IPGRI (2003) descriptors as a basis, as well as the descriptors used by Pitrat et al. (2000) to define the melon subspecies and their botanical varieties.
MATERIALS AND METHODS
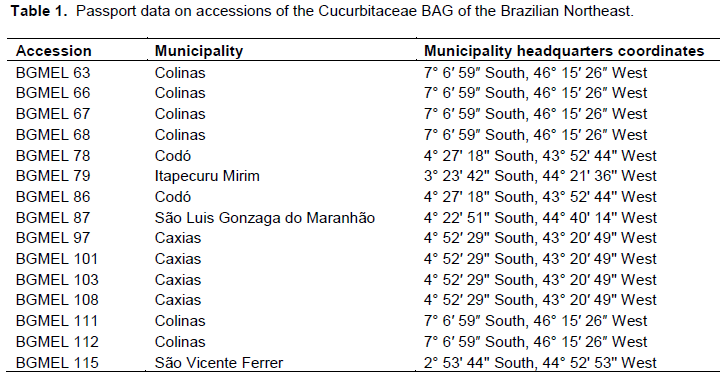
15 accessions collected from traditional agriculture of the state of Maranhão between 1996 and 1998 were evaluated (Table 1); the accessions went through a multiplication phase and were stored in cold chamber with temperature of 10°C and relative humidity of 40%, in the Cucurbitaceae Active Germplasm Bank of the Brazilian Northeast, located at the Embrapa Semiárido, Petrolina – PE.
Experiments were conducted at the Department of Technology and Social Sciences of the State University of Bahia (DTCS/UNEB), located in the municipality of Juazeiro-BA, situated at 09°24'50’’ South latitude and 40°30’10’’ West longitude, with an altitude of 368 m, in the years of 2014 and 2015.
To deploy the first experiment seeds of the accessions derived from open pollination (parental generation) were used, and in the second experiment, seeds of the S1 generation were used. Progenies were obtained through controlled pollination (self-fertilization).
For seedling production, 20 seeds from each accession were sowed on polystyrene trays filled with commercial substrate Plantmax® in a greenhouse covered with shade cloth with 50% light penetration, and irrigated twice a day. After 15 days of sowing, seedlings were transplanted to a definitive site where soil had been previously prepared by ploughing, harrowing, and furrowing. Experimental design was a completely randomized block with four replicates in the first experiment and two replicates in the second experiment. Five plants were used per plot with a spacing of 2.5 m between rows and 0.8 m between plants, watered by furrow irrigation.
In order to perform self-fertilization, one pistillate flower bud and two staminate flower buds from each plant were protected using disposable 200-mL cups and wooden support, and after 24 h, pollination was performed (Figure 1). While conducting the experiments, weeding was performed and the phytosanitary status of the plant was monitored.

Characterization was conducted in both experiments using IPGRI’s (2003) quantitative and qualitative descriptors of plants and fruits, and including the descriptors proposed by Pitrat et al. (2000). Quantitative descriptors were evaluated when fruits were harvested between 30 and 45 days after they were fixed, and were as follows: MF - mean mass of fruits (kg), SND – blossom scar diameter (mm), SIL - blossom scar length (mm), SPT - pulp thickness in the side part of the fruit (cm), UPT - pulp thickness in the upper part of the fruit (cm), LPT - pulp thickness at the lower part of the fruit (cm), CL - fruit cavity length (cm), CD - fruit cavity diameter (cm), PL - fruit peduncle length (mm), PD - fruit peduncle diameter (mm), FD - fruit diameter (cm), FL - fruit length (cm), LSS - Soluble solids in the side part of the fruit (ºBrix), HSS – homogenized soluble solids (ºBrix), and MS - mean mass of 100 seeds (g). Qualitative descriptors were: Fruit shape (globular, flattened, elliptical, pyriform, ovate, elongate, acorn, and malformation), fruit skin color (light yellow, yellow, greenish yellow, bright yellow, light yellow with medium green spots, yellow with dark green spots, pale green, dark green, pale green with dark green stripes, pale green with medium green stripes, and pale green with dark green spots), fruit stripe color (absent, pale green, medium green, and dark green), degree of ribbing in fruits (absent, superficial, intermediate, and deep), fruit skin cracking (present and absent), skin roughness of fruit (absent, superficial, intermediate, pronounced), reticulation (absent, superficial, intermediate, and pronounced), fruit pulp color (white, greenish, orange), fruit placenta color (white, greenish, and orange), fruit aroma (absent, present), fruit abscisson (absent, present), ovary and young fruit pubescence (short, long), and sex expression (monoecious, andromonoecious). To complement the study on fruit phenotypes, a systematic photographic documentation of the fruits (inner and outer parts), of their parental progenies, and respective S1 generation was performed.
If segregation occurs between the parental and S1 generations in each accession, an additional code representing the variation should be added to the accession codes in the Cucurbitaceae BAG of the Brazilian Northeast (Table 1), and these variants would thus be labeled sub-accessions. Therefore, a zero (0) should be added to the accession code of offspring that did not have variations between parental and S1 generations. For instance, if BGMEL63 accession did not show segregation, its new code would be BGMEL63.0, or sub-accession BGMEL63.0. On the other hand, accessions that showed variation between phenotypes of both generations regarding plant, flower, and fruit traits should receive a number according to the number of subdivisions that occurred. Taking accession BGMEL63 as an example again, if three types of offspring occurred in S1 generation, then sub-accessions should be designated BGMEL63.1, BGMEL63.2, and BGMEL 63.3, and so on.
Diversity analysis was based on qualitative and quantitative descriptors. To determine the distance matrix of joint analysis (quantitative and qualitative descriptors), Gower's algorithm (1971) was used. From the dissimilarity matrix, the grouping analysis using the UPGMA method was obtained. In order to validate the clusters generated by the UPGMA method, a cophenetic correlation coefficient (CCC) was estimated based on Pearson's correlation coefficient between the distance matrix and the cophenetic matrix (distance matrix between genotypes) (Cruz et al., 2011). All these analyses were performed with the help of R program (R Development Core Team, 2012).
RESULTS AND DISCUSSION
Comparing the phenotypes of the parental and S1 generations, it was observed that some accessions did not show variation in flower and fruit traits, while others showed high variation (Table 2). Hence, the 15 accessions initially studied were subdivided in 26 sub-accessions according to the traits shown in the table.
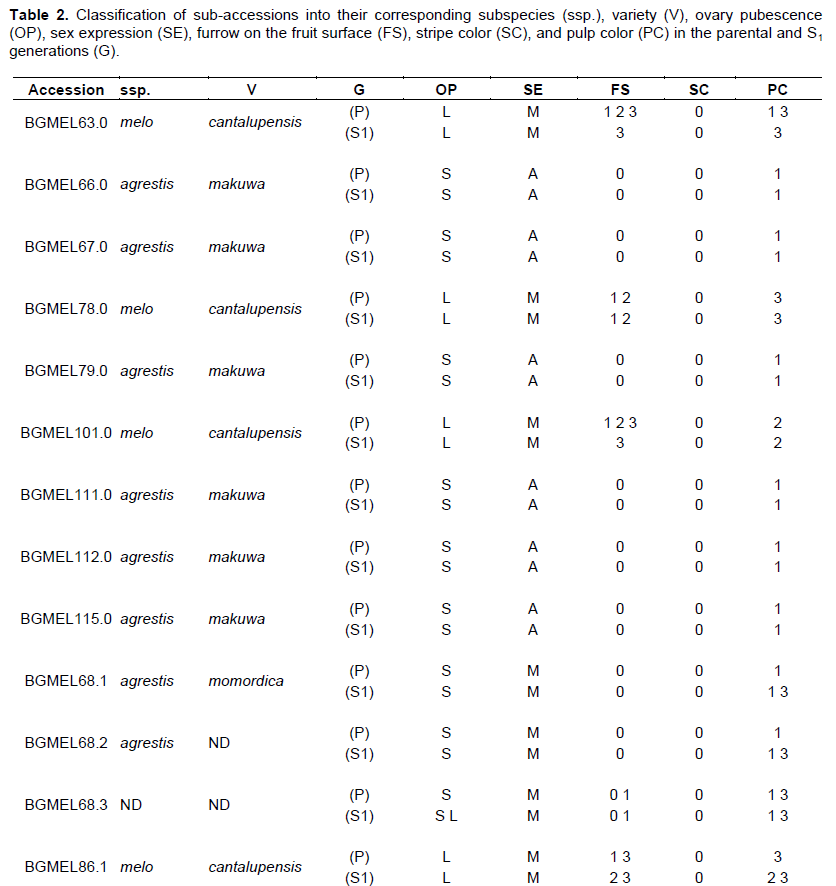
Comparing data on both generations, four sets were identified. The first one had nine subaccessions; some of them were comprised of C. melo subsp. melo variety cantalupensis (BGMEL63.0, BGMEL78.0, and BGMEL101.0) and the others were comprised of C. melo subsp. agrestis var. makuwa (BGMEL66.0, BGMEL67.0, BGMEL79.0, BGMEL111.0, BGMEL112.0, and BGMEL115.0). All plants in these sub-accessions showed the same phenotype of flower and fruit traits (homozygotes) and they also showed descriptors which allowed for the identification of subspecies and respective botanical varieties (Figure 2 and Table 2).

The second set was comprised of seven sub-accessions derived from variations within accessions (parental generation): BGMEL68, BGMEL87, BGMEL108, BGMEL86, BGMEL 87, BGMEL 97, and BGMEL 103 (Table 2 and Figure 3). Some progenies were homozygous and had plant and fruit traits that also allowed for the identification of their subspecies and respective botanical varieties. Hence, C. melo subsp. agrestis var. momordica (sub-accessions BGMEL68.1 and BGMEL87.1), as well as variety makuwa (subaccession BGMEL108.1) was identified. C. melo subsp. melo was also identified, with the variety cantalupensis (sub-accessions BGMEL86.1, BGMEL87.2, BGMEL97.1, and BGMEL103.1) (Figure 3). Therefore, although these sub-accessions were selected based on the offspring that did not show segregation between the parental and S1 generations, they did show morphological traits in fruits and flowers that allowed the identification of the subspecies and varieties of each sub-accession.

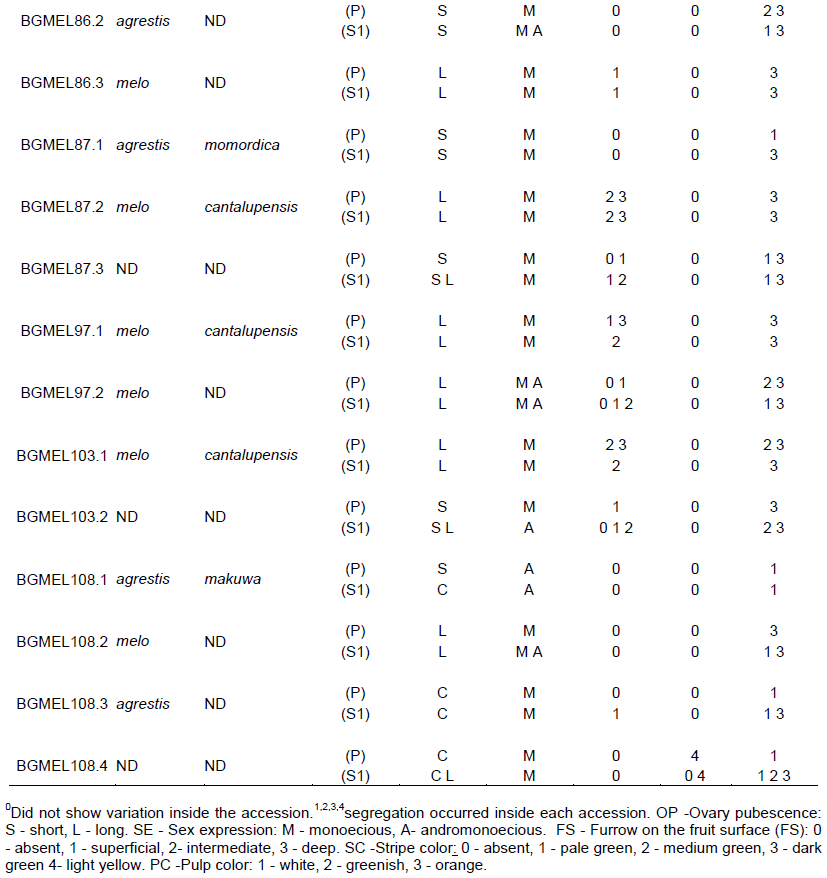
In the third set, a group of offspring showed marked variations in fruit traits between both generations (Figure 4); however, ovary pubescence characters did not vary between the parental and S1 generations (Table 2) and it was therefore possible to identify the subspecies. There are six sub-accessions in this set: BGMEL68.2, BGMEL86.2, BGMEL108.3, BGMEL86.3, BGMEL97.2, and BGMEL108.2; the first three belong to C. melo subsp. agrestis, and the three latter belong to C. melo subsp. melo (Figure 4 and Table 2).

Most of the traits of sub-accession BGMEL68.2, C. melo subsp. agrestis, fit into variety momordica; however, there was no fruit cracking (Figure 4). Accessions belonging to var. momordica showed fruit skin cracking when ripe, white, cream, or orange pulp color, and absence of sugar and aroma (Fergany et al., 2010), and therefore, the data in this study do not support the inclusion of this sub-accession in the variety momordica. Although fruit cracking is determining in variety momordica, it is an undesired trait since it affects appearance and decreases post-harvest durability, compromising commercialization (Neitzke et al., 2009). Nonetheless, melon populations of variety momordica were observed with a slight aroma when ripe, little sweetness, and pulp color ranging from cream to orange (Manohar and Murthy, 2012). On the other hand, sub-accession BGMEL 86.2, also belonging to C. melo subsp. agrestis, showed a variation in traits, and it was therefore impossible to determine its variety. It showed pale green and light yellow skin color, greenish and orange pulp color, absence of furrow and superficial reticulation (Figure 4), possibly a typical trait of groups derived from C. melo subsp. melo. Sub-accession BGMEL108.3, C. melo subsp. agrestis, showed fruit traits typical of C. melo var. acidulus; however, skin color and some data related to fruit pulp did not allow its inclusion in this variety, since accessions belonging to variety acidulus are mainly characterized by having a very hard white pulp, and absence of sugar and aroma (Figure 4) (Fergany et al., 2010).
Among the sub-accessions of C. melo subsp. melo, sub-accession BGMEL86.3 showed all characters of the variety reticulatus; nonetheless, its monoecious sex expression does not coincide with this variety (Table 2 and Figure 4). In sub-accession BGMEL108.2 (Figure 4), most traits (roughness, late maturation, fruit shape and color) fit into variety inodorus; however, due to sex expression and pulp color this sub-accession was not assigned to this variety. Finally, sub-accession BGMEL97.2 showed fruit traits of variety chito. However, the fruits showed pulp color orange, hampering its classification into this variety (Figure 4). On the other hand, the fruit size ranged from 0.4 to 0.8 kg indicating fruit size small to intermediate (Ipgri, 2003) which is not typical of chito variety that has very small fruit size, less than 0.4 kg (Ipgri, 2003; Pitrat et al., 2000).
Sub-accessions where subspecies or botanical groups were not identified (sub-accessions BGMEL87.3, BGMEL103.2, BGMEL68.3 and BGMEL 108.4) comprised the last set. Cracking, pulp color and fruit shape in sub-accessions BGMEL68.3 and BGMEL87.3 indicated the variety momordica (Figure 5); however, there was segregation of the descriptor ovary pubescence that defines the melon subspecies (Jeffrey, 1980) as shown in Table 2, since some plants had long trichomes, both in the parental and S1 generations, and others had short trichomes inside the same accession. This made it impossible to determine the subspecies, and consequently, the variety. The same performance occurred in the other sub-accessions of this set (Table 2). These results show trait introgression between the two melon subspecies. Dogimont (2011), when studying melon genes, did not allude to inheritance of ovary pubescence, and therefore, it is important to study this trait to help define melon subspecies. Regarding the defining traits of varieties (Pitrat et al., 2000), the presence of furrow and pulp color in sub-accession BGMEL103.2 fit into variety cantalupensis, and it was not possible to associate sub-accession BGMEL108.4 to any variety due to the great variation in traits such as stripes, fruit surface and cracking in the parental generation, and pulp color (Figure 5). Due to the fact that it was not possible to determine the subspecies, therefore, it was not possible to assign sub-accessions to a specific variety. However, these results show strong evidence of trait introgression between different melon varieties.

Torres Filho et al. (2009), conducting the morphological characterization of melon accessions collected in the Brazilian Northeast following the classification by Munger and Robinson (1991), identified 80.9% of the 42 accessions analyzed as to their varieties; however, 19.1% remained indeterminate. Among the varieties identified are var. conomon, var. cantalupensis, var. momordica, and var. inodorus (commercial cultivar). In a previous study (data not published), analyzing melon accessions collected in the state of Maranhão following the classification by Pitrat et al. (2000), it was possible to classify the subspecies and varieties of 44.0% of the studied accessions; 32.0% were identified by subspecies, although their varieties remained not defined, and 25.0% was not defined for both subspecies and varieties. Among the varieties defined are: C. melo var. momordica, C. melo var. cantalupensis, C. melo var. conomon, and C. melo var. chandalak. In this study, despite the marked variations in traits that allowed the distinction between subspecies and varieties, only 61.5% of the studied sub-accessions were classified by subspecies and varieties, while 23.1% had only subspecies defined, and 15.4% remained undefined for both subspecies and varieties. However, data showed that there was trait introgression between the different subspecies and their varieties and these results indicate that the management of seeds of different types of melons by family farmers might have contributed to cross-pollination between subspecies and varieties, since there are no barriers to cross-pollination between different botanical varieties (Decker-Walters et al., 2002). The exchange of seeds between family farmers is quite common, and it is possible that these seeds were mixed through germplasm exchange among family farmers.
In addition to studies conducted in Brazil, other countries have also been developing studies with melon germplasm derived from family farming. Szamosi et al. (2010) observed a wide diversity of morphological characters comparing germplasm of Hungarian melons and Turkish melons of the varieties reticulatus, inodorus, cantalupensis, dudaim, chate, chito, and flexuosus. Yildiz et al. (2014), when comparing morphological and molecular characterization of melons of varieties inodorus, cantalupensis, reticulatus, canomon, flexuosus, dudaim, momordica, and six unknown accessions, observed a high variation in the traits of Turkish melons. Trimech et al. (2013), following the classification by Munger and Robinson (1991), observed significant differences in Tunisian melons among accessions and among and within sampling sites, as well as in different varieties reticulatus, inodorus, and dudaim, and some accessions remained indeterminate because they did not fit into any melon variety, similar to the study conducted in Brazil. However, all these studies, Brazil included indicated that the variation found in these melon subspecies and their varieties make this germplasm very important to melon breeding. It is important to emphasize that different melon varieties, notably the wild types that generally belong to C. melo subsp. agrestis, pose great interest from the germplasm standpoint, since they have the majority of genes responsible for controlling biotic stresses caused by fungi such as powdery mildew (Podosphaera xanthii), gummy stem blight (Didymela bryoniae), alternaria leaf blight (Alternaria cucumerina), potyviruses (PRSV-W, WMV, ZYMV), and resistance to insects such as the leaf miner (Liriomyza spp.), among other biotic stresses (Dogimont, 2011).
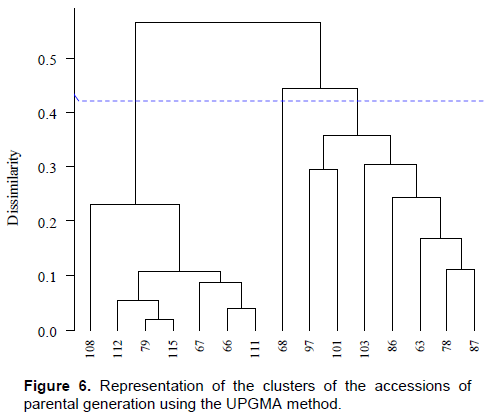
On the other hand, in addition to the high variation found in subspecies and respective varieties, the study on the diversity found in accessions and sub-accessions using morphological descriptors gives a dimension of the existing variability. Clusters formed by the joint analysis of quantitative and qualitative descriptors obtained using the UPGMA method showed that data from the first experiment comprised three groups (Figure 6). The cophenetic correlation coefficient (CCC) was 0.96, indicating a good representation (Cruz et al., 2011), and the cut-off point of dendograms was obtained by Mojena (1977) method. The first cluster was comprised of accessions BGMEL108, BGMEL112, BGMEL79, BGMEL115, BGMEL67, BGMEL66, and BGMEL111. These accessions belong to the subspecies agrestis variety makuwa. It is worth noting that accession BGMEL 108 showed variation in fruit traits among plants within the accession (Figures 3 to 5), as described previously, although it was found one sub-accession of the variety makuwa (sub-accession 108.1) in this variation. Accession BGMEL 68 was the only one comprising the second group and accessions BGMEL 97, BGMEL 101, BGMEL 103, BGMEL86, BGMEL63, BGMEL78, and BGMEL87 comprised the third cluster. This group was comprised of the accessions of subspecies melo variety cantalupensis, although some accessions, such as BGMEL86 and BGMEL87, had variations coinciding with subspecies agrestis (Table 2).
The cluster analysis of sub-accessions (Figure 7) showed that four groups were formed with good consistency (CCC = 0.82). Sub-accessions (BGMEL108.3, BGMEL108.4, BGMEL111.0, BGMEL112.0, BGMEL115.0, BGMEL108.1, BGMEL79.0, BGMEL66.0, and BGMEL67.0) comprised the first group. It was possible to identify two subgroups in this first group: one comprised of BGMEL108.3, belonging to subspecies agrestis and with indeterminate varieties although it had traits of the variety acidulus, and sub-accession BGMEL108.4, whose subspecies was not identified due to segregation of ovary pubescence between the parental and S1 generations (Table 2). The second subgroup gathered all the other sub-accessions of subspecies agrestis variety makuwa.
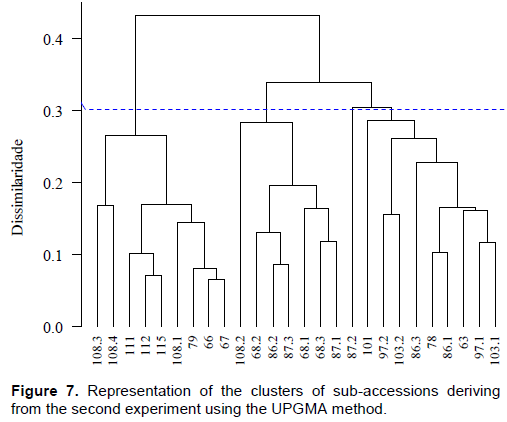
In the second cluster, comprised of seven sub-accessions, there were also two subgroups, the first one comprised of sub-accession BGMEL108.2, belonging to C. melo subsp. melo and variety not defined yet with characters of var. inodorus, and the other one comprised of the other six sub-accessions; two from C. melo subsp. agrestis, var. momordica (BGMEL 68.1 and 87.1), two from C. melo subsp. agrestis and indeterminate variety (BGMEL68.2 and BGMEL86.2), and two with indeterminate subspecies and varieties (BGMEL87.3 and BGMEL68.3), although both had characters of var. momordica. Therefore, the two subspecies and sub-accessions with characters of different varieties were assigned to this group, which indicates again the occurrence of trait introgression both in subspecies and varieties and that this might be a consequence of the management of seeds by farmers, as indicated previously, since there is no cross-pollination barrier between subspecies or between their respective varieties (Decker-Walters et al., 2002).
The third group was comprised only of sub-accession BGMEL 87.2, assigned to C. melo subsp. melo var. cantalupensis, and the last group included nine sub-accessions (BGMEL101.0, BGMEL97.2, BGMEL103.2, BGMEL86.3, BGMEL78.0, BGMEL86.1, BGMEL63.0, BGMEL97.1, and BGMEL103.1), all from C. melo subsp. melo, except for sub-accession BGMEL 103.2, which remained indeterminate due to segregation in ovary pubescence (Table 2) although it had characters of var. cantalupensis. Sub-accession BGMEL 97.2 showed characters of variety chito, BGMEL86.3 had characters of variety reticulatus, and all the others belonged to var. cantalupensis. Thus, this cluster was practically comprised of sub-accessions of C. melo subsp. melo, with only one exception. Thus, clusters of sub-accessions showed a reasonable agreement, as all accessions of C. melo subsp. agrestis were in the first group, except for sub-accession BGMEL108.4, which is indeterminate (as detailed previously). The second group had a mixture of sub-accessions of both subspecies, with predominance of C. melo subsp. agrestis, again indicating introgression of characters of subspecies and their varieties. The third group had only one accession of C. melo subsp. melo var. cantalupensis (BGMEL87.2, Figure 7) and the fourth group had all sub-accessions, with only one exception, of C. melo subsp. melo, and most of them belonged to var.
cantalupensis. It is worth noting that, in addition to the morphological traits that are determining in the identification of subspecies and their varieties, some other morphological traits might also have been important to form the groups. Fruit mass is one example, since fruits were found with a high variation between sub-accessions, ranging from 400 to 2500 g, and there were other very contrasting characters. Therefore, this variation might have been determining to form the third group; even though this group is comprised of C. melo subsp. melo var. cantalupensis, it has been separated from other sub-accessions of the same species and variety of the fourth cluster.
Although there was a trend towards clustering according to subspecies and varieties (groups I and IV, Figure 7), different subspecies were included in group II; moreover, detailed data on subspecies and varieties indicate that there is a high trait introgression, which was more evident in the study of the samples in the present study than in the study performed previously (data not published), in which a sample of accessions from traditional agriculture was analyzed. It is also important to emphasize that some varieties could not be identified only because of the lack of agreement of one or few traits. When these traits are not fixed in the accessions, obtaining new inbred generations might allow the separation of characters that enable the identification of the species and new varieties. Even if such separation is not possible, the existence of characters of different varieties, notably C. melo subsp. agrestis, indicates that this germplasm is very valuable for the identification of resistant genes to biotic stresses faced by commercial melon plant cultures. Therefore, it should be preserved in the short, medium, and long term, as a treasure that, once unveiled, might help develop new commercial melon cultivars to be used in several melon production systems throughout the country.
CONCLUSIONS
1. Several different melon varieties occur in the genetic material of melon grown by family farming in the Brazilian Northeast.
2. There is variability among and within samples of melon accessions and sub-accessions in the melon germplasm bank derived from family farming in the Northeastern Brazil.
3. There is trait introgression between the two melon subspecies and among their different varieties in the germplasm cultivated by small farmers.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Agrianual (2014). Anuário da agricultura brasileira. Cultura do melão. |
|
|
Aragão FAZ, Torres Filho J, Nunes GHS, Queiróz MA, Bordallo PN, Buso GSC, Ferreira MA, Costa ZP, Bezerra Neto F (2013). Genetic divergence among accessions of melon from traditional agriculture of the Brazilian Northeast. Genet. Mol. Res. 12(4):6356-6371. |
|
|
Bates DM, Robinson RW (1995). Cucumber, melons and watermelons. In: Smartt JE, Simmonds NW. Evolution of Crop Plants, 2nd edn. Longman, London 2:89-96. |
|
|
Burger Y, Paris HS, Cohen R, Katzir N, Tadmor Y, Lewinsohn E (2010). Genetic Diversity of Cucumis melo. Hortic. Rev. 36:165-198. |
|
|
Correa SMS (2010). Africanidades na paisagem brasileira. INTERthesis: Rev. Int. Interdis. 7(1):96-116. |
|
|
Costa ND (2008). A cultura do melão. Embrapa Semi-Árido; 2. Ed. Versão Ampliada - Brasília, DF: Embrapa Inform. Tecnol. pp. 18-20. |
|
|
Cruz CD, Ferreira FM, Pessoni LA (2011). Biometria aplicada ao estudo da diversidade genética. Viçosa: UFV, 620 p. |
|
|
Decker-Walters DS, Chung SM, Staub JE, Quemada HD, López-Sesé AI (2002). The origin and genetic affinities of wild populations of melon (Cucumis melo, Cucurbitaceae) in North America. Plant Syst. Evol. 233:183-197. |
|
|
Dogimont C (2011). Gene list for melon. Cucurbit Genet. Coop. Rep. 33-34:104-133. |
|
|
Fergany M, Kaur B, Monforte AJ, Pitrat M, Rys C, Lecoq H, Dhillon NPS, Dhaliwal SS (2010). Variation in melon (Cucumis melo) landraces adapted to the humid tropics of southern India. Genet. Resour. Crop Evol. 58: 225-243. |
|
|
Gower JC (1971). A general coefficient of similarity and some of its properties. Biometrics Arlington 27(4):857-874. |
|
|
IPGRI (2003). Descriptors for melon (Cucumis melo L.). Rome: IPGRI. 65 p. |
|
|
Jeffrey C (1980). Further notes on Cucurbitaceae: V. The Cucurbitaceae of the Indian subcontinent. Kew Bull. 34:789-809. |
|
|
Manohar SH, Murthy HN (2012). Estimation of phenotypic divergence in a collection of Cucumis melo, including shelf-life of fruit. Sci. Hortic. 148:74-82. |
|
|
Mojena R (1977). Hierarchical grouping method and stopping rules: an evaluation. Comput. J. 20:359-363. |
|
|
Munger HM, Robinson RW (1991). Nomenclature of Cucumis melo L. Cucurbit Genet. Coop. 14:43-44. |
|
|
Neitzke RS, Barbieri RL, Heiden G, Büttow MV, Oliveira CS, Corrêa LB, Schwengber JE, Carvalho FIF (2009). Caracterização morfológica e dissimilaridade genética entre variedades crioulas de melão. Hortic. Bras. 27:534-538. |
|
|
Pitrat M, Hanelt P, Hammer K (2000). Some comments on interspecific classification of cultivars of melon. Acta Hortic. Belgium 510:29-36. |
|
|
R Development Core Team (2012). A language and environment for statistical computing.Vienna. R Foudation for Statistical Computing. |
|
|
Renner SS, Schaefer H, Kocyan A (2007). Phylogenetics of Cucumis (Cucurbitaceae): Cucumber (C. sativus) belongs in an Asian/Australian clade far from melon (C. melo). BMC. Evol. Biol. 7:58-69. |
|
|
Robinson RW, Decker-Walters DS (1997). Cucurbits. CAB International. Oxon (GB). 226 p. |
|
|
Schaefer H, Heibl C, Renner SS (2009). Gourds afloat: a dated phylogeny reveals an Asian origin of the gourd family (Cucurbitaceae) and numerous oversea dispersal events. Proc. Biol. Sci. 276:843-851. |
|
|
Sebastian P, Schaefer H, Telford IRH, Renner SS (2010). Cucumber (Cucumis sativus) and melon (C. melo) have numerous wild relatives in Asia and Australia, and the sister species of melon is from Australia. Proc. Natl. Acad. Sci. USA.107:14269-14273. |
|
|
Silva AF, Santos CAF, Araújo FP, Lima Neto FP, Moreira JN, Ferreira MAJF, Leão PCS, Dias RCS, Albuquerque SG (2010). Recursos genéticos vegetais conservados na Embrapa Semiárido. In: Sá IB, Silva PCG (eds) Semiárido Brasileiro: Pesquisa, Desenvolvimento e Inovação. Petrolina: Embrapa Semiárido pp. 275-315. |
|
|
Szamosi C, Solmaz I, Sari N, Barsony C (2010). Morphological evaluation and comparison of Hungarian and Turkish melon (Cucumis melo L.) germplasm. Sci. Hortic. 124:170-182. |
|
|
Torres Filho J, Nunes GHS, Vasconcelos JJC, Costa Filho JH, Costa GG (2009). Caracterização morfológica de acessos de meloeiro coletados no nordeste brasileiro. Rev Caatinga 22(3):174-181. |
|
|
Trimech R, Zaouali Y, Boulila A, Chabchoub L, Gheza I, Boussaid M (2013). Genetic variation in Tunisian melon (Cucumis melo L.) germplasm as assessed by morphological traits. Genet. Resour. Crop Evol. 60:1621-1628. |
|
|
Yildiz M, Akgul N, Sensoy S (2014). Morphological and Molecular Characterization of Turkish Landraces of Cucumis melo L. Notulae Bot. Horti Agrobot. Cluj-Napoca 42(1):51-58. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


