Full Length Research Paper

ABSTRACT
The objective of this study was to find out the influence of soil nutrition on the composition of volatile oils of Porophyllum ruderale subsp. macrocephalum and Porophyllum ruderale subsp. ruderale. The seedlings were transplanted into pots containing sand as substrate. Every seven days, different Hoagland solutions were applied: Complete solution and solution lacking, respectively nitrogen, phosphorus, potassium, calcium, magnesium, sulfur and a control. The experiment was done in a randomized block with eight treatments and five replicates. For biometrics, height, leaf area and dry mass of shoots were analyzed. The average was compared by Tukey test (5%) probability. Aerial parts were collected and oil was extracted by hydrodistillation in Clevenger apparatus for 4 h. The major components of P. ruderale subsp. ruderale were trans-β-ocimene monoterpene, limonene and hydrocarbon undecene. As for the P. ruderale subsp. macrocephalum, the major component was monoterpene limonene and undecene hydrocarbon. The results of biometric analyses in this work showed that the two species have different growth. Treatments without nitrogen, phosphorus and the control had the lowest average and increased undecene content in the two subspecies. The chemical composition of volatile oils nutrition did not interfere significantly in their composition. Monoterpene limonene was the highest in macrocephalum subspecies.
Key words: Brazilian medicinal plants, fertilizers.
INTRODUCTION
Brazil is a country with a rich genetic diversity in both flora and fauna. In its flora, there have been cataloged more than 55,000 species of an estimated total of 350,000 to 550,000 species. The increased genetic diversity and chemical diversity allow the use of plants as invaluable source of bioactive compounds for the development of new drugs (Gottlieb et al., 1996). Plants produce a wide variety of organic compounds that are economically important, such as alkaloids, resins, tannins, volatile oils, among others (Croteau et al., 2000). The growing demand for medicinal plants constitutes a concern for conservation. Because extraction threatens more native species (Ioris et al., 1999), a more detailed study is needed on native vegetation for the recovery of species of medicinal value (Dias, 2000).
According to Di Stasi et al. (2002), among the plant species of therapeutic interest, the Asteraceae family can be considered as one of the most important. This is because the large number of plants belonging to it, such as Baccharis trimera (gorse) and Achillea millefolium (“novalgina”) in addition to arnicas, guacos and bidens, are popularly used as drugs. Most of these species are native to Brazil, while several others have acclimatized and can be found throughout Brazilian territory, which have been incorporated in traditional medicine.
Porophyllum ruderale (Jacq.) Cass., popularly known as arnica is a ruderal herb used in folk medicine for healing, as haemostatic (Goleniowski et al., 2006), antitinflamatória (Lima et al., 2010) and for combating leishmaniasis (Takahashi et al., 2011). There are two subspecies of Porophyllum used for the same purpose: P. ruderale subsp. ruderale and P. ruderale subsp. macrocephalum. The two subspecies have the same common name and share some morphological characters, such as herbaceous and similar inflorescences, attributted to the misuse of species. The chemical composition of the volatile oil of the two subspecies is different. The major compounds in P. ruderale subsp. ruderale are monoterpenes trans-β-ocimene, α-pinene and limonene, while in P. ruderale subsp. macrocephalum, there are monoterpenes limonene and β-pinene (Raggi et al., 2010). There are few works done on the subspecies, ruderale and macrocephalum. Some authors have reported the presence of β-phellandrene (Fonseca et al., 2006), sabinene (Loayza et al., 1999), limonene and 7-tetradecene (Guillet et al., 1998) as major compounds in the chemical composition of the volatile oil of P. ruderale.
According to Martins et al. (1995), of all the factors that can interfere with the active principles of plants, nutrition is one of those that require more attention, as excess or deficient nutrients can be directly correlated to changes in the production of active substances. Evaluating the influence of mineral nutrition on yield and volatile oil composition of Ocimum basilicum (basil), Coriandrum sativum (coriander), Anethum graveolens L. (dill), and Mentha piperita L. (Mint) and Hornok (1983) reported the occurrence of variations based on the four levels of NPK used. With increased phosphorus level, there was an increase in the volatile oil content of mint and basil and low volatile oil content of biomass and dill. High nitrogen levels increased the essential oil of mint and basil, but reduced the percentage of menthol and linalool. Also in relation to nitrogen levels, the authors observed an increase in the production of green biomass of dill and cilantro, but not seeds.
The species of the genus Porophyllum are widely used in folk medicine of South and Central America. However, there are phytotechnical works related to soil nutrition and the chemical composition of the volatile oils of species P. ruderale subsp. macrocephalum and P. ruderale subsp. ruderale. The objective of this project was, consequently, to study the effect nutrition management on plant development and production of volatile oils of arnica’s subspecies.
This work aimed to evaluate the management of mineral nutrition for plant growth, yield and chemical composition of the volatile oils of P. ruderale subsp. ruderale (Jacq.) Cass. and P. ruderale (Jacq.) Cass. subsp. macrocephalum (DC.) R. R. Johnson.
MATERIALS AND METHODS
Seedlings of P. ruderale subsp. ruderale and P. ruderale subsp. macrocephalum were obtained from seeds and grown in a greenhouse at the Experimental Field of Ornamental Plants Research Center, São Paulo Botanic Institute.
The seeds used were obtained from mother plants of PEFI (State Park Ipiranga Font) and sown in polystyrene trays with 128 cells, using Tropstrato HT Hortaliças® as substrate. After the appearance of four pairs of permanent leaves, the seedlings were transplanted into plastic pots of 10.5L, using sand as substrate and placed in a greenhouse equipped with microsprinkler irrigation system, in the Experimental Field Ornamental Plants. The pots were fertilized once a week with 50 ml of nutrient solutions of Hoagland and Arnon (1950), under different managements: complete solution, solution without nitrogen (-N3-), solution without phosphorus (-P5+), solution without potassium (-K+), solution without magnesium (-Mg2+), solution without calcium (-Ca2+), solution without sulfur (-S2-), and a control with 50 mL distilled water. The experiment was done in a randomized block design with eight treatments for each species, five replications and four vessels.
The height of both subspecies was taken with a measuring tape in centimetre from the soil surface to the highest part of the plant. The first measurement was done 30 days after transplanting, while the other was done every 20 days until the end of the experiment (130 days) after treatments application. To obtain dry mass, three individuals from each treatment were weighed on an analytical balance, placed in paper bags and oven-dried at 45°C for 15 days. The leaf area was determined in a meter Model LI-3100C expressed in square decimetres. Leaf area is defined as the average of the areas of all the leaf blades of the three subjects per treatment (Benincasa, 2003).
All data collected were submitted to Tukey test at 5% significance using the SISVAR program (Ferreira, 2002).
For extraction and identification of volatile oil, plant materials were collected at 130 days after treatment application, in the morning. The plant materials were stored in transparent bags and kept in a freezer at -22°C for later extraction in the Center for Research in Physiology and Biochemistry, São Paulo Botanic Institute. The shoots were subjected to extraction to obtain volatile oil in a continuous process by hydrodistillation using Clevenger apparatus. They were adapted to a round bottom flask of 6000 mL, for 4 h in three replicates for each treatment. After this period, the volatile oil was removed from the apparatus and stored in an Erlenmeyer. For the complete removal of the volatile oil, solvent pentane was added. Anhydrous sodium sulfate was used to remove any water present in the volatile oil. The volatile oil containing pentane was transferred to a vial with cap. With the aid of rotatory evaporator at room temperature, pentane was completely removed leaving only the pure volatile oil. The oil mass was determined with the aid of an analytical balance. The oils were stored in cardboard box in a freezer at -22°C for further identification of its constituents.
The yield of each oil was calculated from the fresh mass (Santos et al., 2004; Pino et al., 2006; Kelen and Tepe, 2008).
To identify the constituents, the extracted volatile oils were dissolved in acetone 1:10 (v/v) and 2 uL of each diluted sample was analyzed by gas chromatography on Agilent apparatus (6890 series) HP, coupled to spectrophotometer mass with quadrupole system (Agilent 5973 Network mass Selective Detector), and 70 eV ionization energy. The capillary column used was HP-5 MS (30 m × 0.25 mm internal diameter, 0.25 thickness) under the following conditions: gun (with flow-split split/splitless) to 250°C (split ratio 1: 20), the column of heating temperature of 40 to 240°C to 3°C min-1, 240°C for 10 min using helium as carrier gas at a pressure of 80 kPa and a linear speed of 1 ml min-1. The total analysis time was 78 min. Nitrogen, synthetic air and hydrogen are used as auxiliary gases in the ratio of 1:1:10, respectively. The retention index (RI) was calculated on HP-5 MS column using a homologous series of n-alkanes (C5 to C30) under the same conditions for chromatographic analysis. The identification of the compounds was made by comparing mass spectra to those registered in the library database 275.
RESULTS AND DISCUSSION
Growth of P. ruderale subspecies under differents nutrient solutions of Hoagland
It was observed that the growth of plants P. ruderale subsp. ruderale and P. ruderale subsp. macrocephalum was differently influenced by the different treatments.
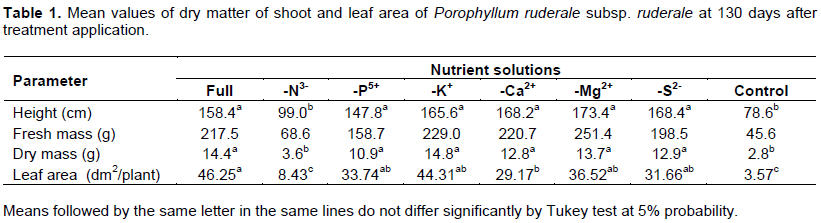
The mean height of the subspecies ruderale treated with complete nutrient solution at 130 days after treatment application (DAT) was 144.8 cm; while macrocephalum was 126.1 cm in the same period.
The highest mean of plant height was obtained with the complete solution treatment and treatments without Ca2+, Mg2+ and S2-, respectively. Subspecies ruderale that had the lowest plant height mean was treated with the nutrient solution minus nitrogen (–N3-) and the control. The means obtained with these treatments (–N3 and control) signifcantly differed from the others height means.
The descending order height for macrocephalum subspecies was: -Ca2+, -Mg2+, - S2-, complete solution, -K+, -P5+, -N3- and control; for subspecies ruderale: -Mg2+, -Ca2+, -S2-, -K+, complete solution, -P5+, -N3- and control. Importantly for both subspecies, in -N3- treatment and control, the plants are not developed satisfactorily.
In this study, the two subspecies responded similarly to the treatments without nitrogen (N3-) and the control; they had significantly lower growth compared to other treatments (Tables 1 and 2). According to Kerbauy (2008), nitrogen is essential for plant growth and its deficiency significantly influences the growth of the plant. Nitrogen is the mineral element required in larger quantities by plants and is the constituent of many components of the plant cell (Castro, 2007). It has a structural function in the plant and parts of many cell components such as proteins, nitrogenous bases, nucleic acids, enzymes, co-enzymes, vitamins and pigments. It, also, participates in processes such as ion absorption, photosynthesis, respiration, cell multiplication and differentiation (Malavolta, 2006). Its deficiency rapidly inhibits plant growth (Taiz and Zeiger, 2004).
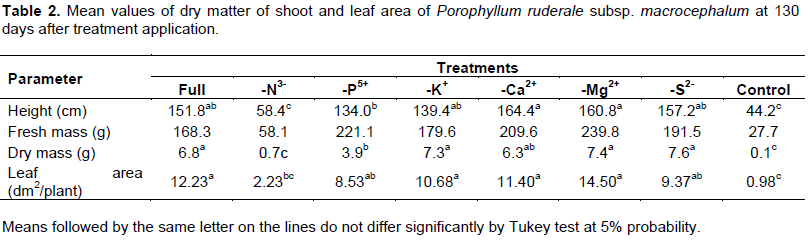
At 130 days, the height mean of plants treated with the nutrient solutions -Ca2+ and -Mg2+ were not statistically different from each other and were similar to the one of the plants which received full Hoagland nutrient solution. This showed that the absence of this nutrient did not negatively affect the development of macrocephalum subspecies (Tables 2 and 3).

Although many studies conducted showed that the absence of calcium and magnesium dramatically affects the development of plant, their absence effect differed in the subspecies Porophyllum since in both species the absence of these nutrients did not affect their growth.
The application of nutrient solution minus P did not negatively affect the height, dry mass and leaf area of P. ruderale subsp. ruderale comparatively to the ones of the plants that received full nutrient solution of Hoagland. Phosphorus is involved in the storage and transfer of energy and nitrogen fixation; nitrogen is connected to the rapid development of roots, accelerated fruit ripening, increased fruiting as carbohydrate content, oils, fats and proteins (Malavolta, 2006).
According to Malavolta (2006), nitrogen is required for flowering. Without this nutrient, the emission of flowers will be retarted. This is because the limitation of nutrient reduces the assimilation of CO2 through photosynthesis, and after a variable period it ends up decreasing biomass production, photosynthesis and stomatal conductance (Fujita et al., 2003).
Phosphorus has fundamental function in the life of plants. As part of compounds, it is rich in energy like adenosine triphosphate (ATP) (Malavolta, 1985), and through this energy, seeds germinates, plant performs photosynthesis, absorbs actively soil nutrients and synthesizes various organic compounds (Naiff, 2007). Phosphorus is also involved in essential functions of cellular metabolism, which acts in the synthesis of metabolites and complex molecules such as DNA, RNA and phospholipids, in the electron transport chain, redox reactions promoting the regulation of the rate of several enzymatic reactions and metabolic processes, such as respiration and photosynthesis (Alves et al., 1996).
As also noted with ruderale subspecies, the indicative biometric variables of macrocephalum subspecies growth were also significantly affected by nitrogen omission.
Symptoms of deficiency or excess of a mineral element are similar in all plant species (Leal and Prado, 2008). According to Pozza et al. (2001), nutrients perform specific functions in plant metabolism, not only in influencing its growth, but also its production. Thus, a nutrient in abnormal levels can damage production causing a nutritional stress (Deon, 2007). The dry matter production data of shoot and leaf area of the subspecies Porophyllum are shown in Table 2.
Analyzing the variables, for the shoot dry matter production was differently affected by the treatments. The plants treated with the nutrient solution minus nitrogen and the control had significantly lower shoot dry matter of 3.6 and 2.8 g, respectively. As well, the leaf area was differently influenced by the treatments: the control had the lowest mean of 3.57 dm2 per plant and the highest mean of 46.25 dm2 per plant was obtained with the complete nutrient solution.
Table 3 shows the height mean values of shoot dry matter and leaf area of the subspecies macrocephalum. Significant differences were observed between the treatments: the treatments without potassium (-K+), calcium (-Ca2+) and magnesium (-Mg2+) and the complete nutrient solution had the highest mean leaf area.
Chemical composition of the essential oils of the aerial parts of P. ruderale subsp. ruderale as influenced by different nutrient solution of Hoagland
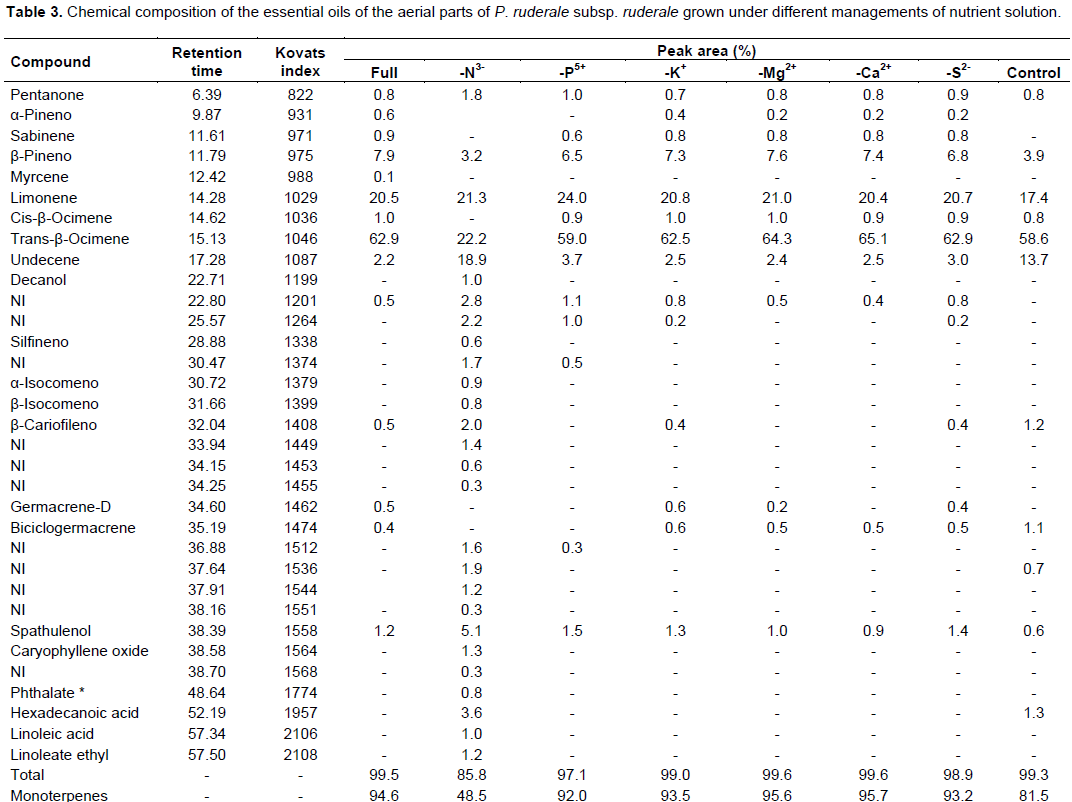
The chemical composition of volatile oils of P. ruderale subsp. ruderale is presented in Table 3. Twenty-two compounds were identified out of thirty compounds. The yield of volatile oil was 0.07% for treatments with complete Hoagland solution and without sulfur (-S2-); 0.06% for treatments without phosphorus (-P5+), potassium (-K+), magnesium (-Mg2+) and calcium (-Ca2+); 0.03% for the control treatment and 0.02% for treatment without nitrogen (-N3-). Monoterpenes were the predominant constituents, ranging from 48.5% (treatment -N3-) to 95.7% (-Ca2+ treatment). The other major compounds were trans-β-ocimene, limonene and hydrocarbon. The different nutrient solution handlings also influenced the chemical composition of the volatile oils except limonene; even the treatments with a greater influence negative (-N3- and control) remained constant at about 20% of the total chemical composition. The major compound was trans-β-ocimene, accounting for about 62%, on average, of the total chemical composition of the volatile oil of subspecies ruderale for almost all treatments except N in which it was 22.2%. The content of the hydrocarbon undecene was greater about eight times in the N treatment (18.9%) and six times in the control treatment (13.7%), relative to the treatment with addition of the complete Hoagland solution.
The presence of sesquiterpenes was also higher in the treatment without nitrogen. The hydrocarbon of undecene was also observed by Raggi et al. (2012) who compared the chemical composition of volatile oils of green material collected at 270 days and aging plant collected at 300 days. Undecene increased by about 10 times in plants of 300 days old relative to the green material. Though, there are no studies on the hydrocarbon of undecene in different plant growth stages.
For the biometric analysis, treatments that led to nutritional deficiency in the chemical composition of volatile oils were -N3- and control.
Chemical composition of the essential oils of the aerial parts of P. ruderale subsp. macrocephalum as influenced by different nutrient solution of Hoagland
The yield of volatile oil of macrocephalum subspecies was lower compared to that of ruderale. For -K+ and -Ca2+ treatments, the yield was 0.04%; for complete and -S2-, -Mg2+ treatments, the yield was 0.03% and for phosphorus, the yield was 0.02%. In -N3- and control treatments, there was no much oil yield due to the small amount of oil obtained. As noted in the biometric analysis, -N3-, -P5+ and control treatments did not produce sufficient biomass to conduct the volatile oil extraction in three repetitions, as the other treatments.
The chemical composition of volatile oils of macrocephalum subspecies is described in Table 4. Of the twenty compounds, thirteen were identified. Monoterpene limonene was the major constituent in almost all treatments except control, whose main component was undecene hydrocarbon (29%). The other treatments showed, on average, 15% of hydrocarbon. -Mg2+ treatment found to have the highest percentage of limonene (70.5 %) and the lowest content of this constituent was found in the control treatment (27.4%). Treatments without potassium and sulfur had the highest percentages of myrcene (18.2 and 18.4%, respectively), compared to the same constituent in the other treatments on average, 8%, except the control treatment containing only 2.6%. The sesquiterpenes content was higher in the control (12.4%) compared to other treatments, 2.7% on average.
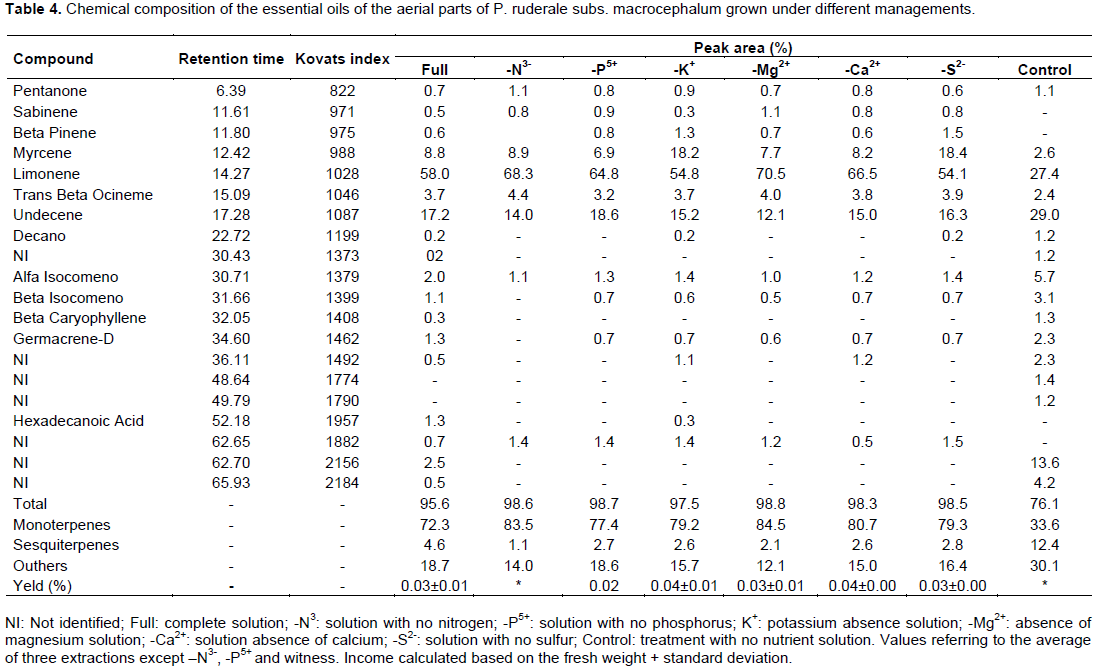
The chemical composition of volatile oil of Porophylum is well reported in the literature. However, little is said about the studied subspecies. The levels of the major constituents were very different. Rondón et al. (2008), working with P. ruderale collected in Venezuela, identified a mixture of limonene and β-phellandrene as the main components of the chemical composition of volatile oils. For plants collected in Bolivia, the main constituent was sabinene (Loayaza et al., 1999). Fonseca et al. (2006) identified β-phellandrene monoterpene in plants collected in Minas Gerais. In Ceará, Neto et al. (1994) said the major compound was limonene (74%).
Many articles report that geographical and climatic variations are responsible for the variation in the chemical composition of volatile oils. However, besides these, there is also correct identification of the material to be studied and the quality of tillage. Raggi et al. (2014), working with both species, concluded that the major components in subspecies macrocephalum and ruderale were limonene and E-β-ocimene, respectively. They noted that the difference in the two species is not caused by climate, but by genetic issues.
Monoterpene limonene present mainly in subspecies macrocephalum has different biological activities such as antimicrobial and anti-inflammatory. These activities are related to its medicinal use (Souza et al., 2003; Lorenzi and Matos, 2008).
CONCLUSION
Both species did not have proper growth development with nitrogen deficiency, phosphorus and control. The chemical composition of volatile oils nutrition did not interefere significantly in their composition. Monoterpene limonene was the highest in macrocephalum subspecies,
CONFLICT OF INTERESTS
The authors have not declared any conflict of interest.
REFERENCES
|
Alves VMC, Novais RF, Oliveira MFG, Barros NF (1996). Efeito da omissão de fósforo na absorção de nitrogênio por híbridos de milho (Zea mays L.). Revista Ceres, Viçosa 58:248. |
|
|
Castro ACR (2007). Deficiência de macronutrientes em helicônia 'Golden Torch'. Programa de Pós-Graduação em Botânica. Universidade Federal Rural de Pernambuco. Recife. |
|
|
Croteau R, Kutchan T, Lewis N (2000). Natural products (secondary metabolites). In: Buchanan, B. B., Gruissen, W. & Jones, R.L. (orgs). Biochemistry & Molecular Biology of Plants. American Society of Plant Biologists. Rockville. |
|
|
Deon MD (2007). Crescimento e nutrição mineral da soja submetida a excesso de P, S, K, Ca e Mg em solução nutritiva. Tese de Mestrado, USP, Escola Superior de Agricultura Luiz de Queiroz, Piracicaba. |
|
|
Di Stasi LC, Hiruma-Lima CA, Santos CM, Guimarães EM (2002). Asterales medicinais. In: Di Stasi L.C.; Hiruma-Lima C.A. (Eds) Plantas medicinais na Amazônia e na Mata Atlântica Editora UNESP. São Paulo. |
|
|
Dias JAB (2000). Produção de Plantas Medicinais e Agricultura Urbana. ANAIS. 40º Congresso de Olericultura; II Congresso Ibero-Americano sobre Utilização de Plástico na Agricultura; I Simpósio Latino-Americano de Produção de Plantas Medicinais, Aromáticas e Condimentares. SOB/FCAV-UNESP 18:140-143. |
|
|
Ferreira DF (2002). SISVAR. Sistemas de análises de variância para dados balanceados: programa de análises estatísticas e planejamento de experimentos. Versão 4.3. Lavras: UFLA. |
|
|
Fonseca MCM, Barbosa LCA, Nascimento EA, Casali VWD (2006). Essential oil from leaves and flowers of Porophyllum ruderale (Jacq.) Cassini (Asteraceae). Journal of Essential Oil Research 18:345-347 |
|
|
Fujita K, Okada M, Lei K, Ito J, Ohkura K, Adu_Gyamfi JJ, Mohapatra PK (2003). Effect of P-deficiency on photoassimilate partitioning and rhytlmic changes in fruit and stem diameter of tomato (Lycopersicon esculentum) during fruit growth. Journal of Experimental Botany Oxford 54:2519-2528. |
|
|
Goleniowski ME, Bongiovanni GA, Palacio L, Nunez CO, Cantero JJ (2006). Medicinal plants from the "Sierra de Comechingones" Argentina. Journal of Ethnopharmacology107:324-341 |
|
|
Gottlieb OR, Kaplan MAC, Borin MRMB (1996). Biodiversidade um enfoque quimico-biologico. Editora UFRJ. Rio de Janeiro. |
|
|
Guillet G, Belanger A, Arnason JT (1998). Volatile monoterpenes in Porophyllum gracile and P. ruderale (Asteraceae): identification, localization and insecticidal synergism with α-tertienil. Phytochemistry 49:423-429 |
|
|
Hornok L (1983). Influence of nutrition on the yield and content of active compounds in some essential oil plants. Acta Horticulturae 132:239-247. |
|
|
Ioris E (1999). Plantas medicinais do cerrado: perspectivas comunitárias para a saúde, o meio ambiente eo desenvolvimento sustentável. Fundaçao Integrada Municipal de Ensino Superior, Projeto Centro Comunitário de Plantas Medicinais. |
|
|
Kelen M, Tepe B (2008). Chemical composition, antioxidant and antimicrobial properties of the essential oils of three Salvia species from Turkish flora. Bioresource Technology 99:4096-4104. |
|
|
Kerbauy GB (Coord) (2008). Fisiologia Vegetal. 2 ed., Guanabara Koogan, São Paulo. 452p. |
|
|
Leal RM, Prado RM (2008). Desordens nutricionais no feijoeiro por deficiência de macronutrientes, boro e zinco. Rev. Bras. de Ciências Agrárias Recife 3:301-306. |
|
|
Loayza I, Groot W, Lorenzo D, Delacassa E, Mondello L, Dugo G (1999). Composition of the essential oil of Porophyllum ruderale (Jacq.) Cass. from Bolivia. Flavour and Fragrance Journal14:393-398 |
|
|
Lorenzi H, Matos FJA (2008). Plantas Medicinais no Brasil: nativas e exóticas cultivadas. Nova Odessa, Instituto Plantarum, São Paulo. |
|
|
Malavolta E (1985). Nutrição mineral. In: FERRI MG. (Coord.). Fisiologia vegetal. 2 ed. São Paulo: EPU. |
|
|
Malavolta E (2006). Manual de nutrição mineral de plantas. São Paulo: Editora. |
|
|
Martins ER., Castro DM, Castellani DC, Dias JE (1995). Plantas medicinais. Viçosa: UFV, Imprensa Universitária. |
|
|
Naiff APM (2007). Crescimento, Composição Mineral e Sintomas Visuais de Deficiências de Macronutrientes em Plantas de Alpinia Purpurata Cv. Jungle King. Tese de Mestrado. Universidade Federal Rural Da Amazônia. Belém. |
|
|
Neto MA, Cunha AN, Silveira ER (1994). Volatile constituents of Porophyllum ruderale Cass. Journal of Essential Oil Research6:415-417. |
|
|
Pino JA, Marbot R, Payo A, Chao D, Herrera P (2006).Aromatc plants from wester Cuba VII. Composition of the leaf oils of Psidium wrightii Krug et Urb., Lantana involucrate L., Cinnamomum montanum (Sw.) Berchtold et J. Persl. And Caesalpinia violaceae (Mill.) Standley. Journal of Essential Oil Research18:170-174. |
|
|
Pozza AAA, Martinez HEP, Caixeta LSC, Cardoso AA, Zambolim L, Pozza EA (2001). Influência da nutrição mineral na intensidade da mancha-de-olho-pardo em mudas de cafeeiro. Pesquisa Agropecuária Brasileira, Brasileira 36:53-60. |
|
|
Raggi L, Cordeiro I, Moreno PRH, Young MCM (2012). Porophyllum ruderale subs. ruderale: composição química dos óleos voláteis de partes aéreas de material verde e senescido. CD-Room da 35ª Reunião da Sociedade Brasileira de Química. Águas de Lindóia. Disponibilizado em: |
|
|
Raggi L, Cordeiro I, Moreno PRH, Young MCM (2010). Composição química dos óleos voláteis de duas subspécies de Porophyllum ruderale. Anais do XXI Simpósio de Plantas Medicinais do Brasil. Universidade Federal de João Pessoa. |
|
|
Raggi L, Young MCM, Cordeiro I, Moreno PRH (2014). Differenciation of two Porophyllum ruderale (Jacq.) Cass. subspecies by the essential oil composition. Journal of Essential Oil Research 27:1-4. |
|
|
Rondón ME, Delgado J, Velasco J, Rojas J, Rojas LB, Morales A, Carmona J (2008). Chenical composition and antibacterial activity of the essential oil from aerial parts of Porophyllum ruderale (Jacq.) Cass. collected in Venezuela, Cience 16:5-9. |
|
|
Santos DMM (2004). Nutrição Mineral. Apostila de Fisiologia Vegetal. UNESP. |
|
|
Souza MC, Siani AC, Ramos MFS, Menezes-de-Lima O, Henri MGMO (2003). Evaluation of anti-inflamatory activity of essential oils from two Asteraceae species. Pharmazie 58:582-586. |
|
|
Taiz L, Zeiger E (2004). Fisiologia vegetal. 3.ed. Porto Alegre: Artmed 49 p. |
|
|
Takahashi HT, Novello CR, Ueda-Nakamura T, Dias Filho BP, Mello JCP, Nakamura CV (2011). Thiophene Derivatives with Antileishmanial Activity Isolated from Aerial Parts of Porophyllum ruderale (Jacq.) Cass. Molecules 16:3469-3478. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


