ABSTRACT
The maize weevil Sitophilus zeamais is a primary pest of stored grains, and it is primarily controlled through the use of synthetic insecticides, which are directly connected to increases in the resistance of these insects. In this study, populations of S. zeamais acquired from small farmers in southern Piauí were obtained, and an experiment was performed with the insecticide K-Obiol EC25, with dose-response concentrations ranging from 3.5 ´ 10-4 to 0.05 g i.a/cm2. Five replicates containing 20 insects were used. For the amylase, lipase and total protease assays, 20 adult insects from each population were used separately for each treatment. The experiments performed with K-Obiol for the insect populations from different locations showed different LC50 values. The highest LC50 value was from Currais at 94.0 µg i.a/cm2, and the lowest value was from Bom Jesus at 14.0 µg i.a/cm2. The specific activities of the energy metabolism enzymes (lipase and amylase) responsible for sugar hydrolysis in four populations of S. zeamais exposed to the insecticide K-Obiol at different times (1, 6, 12 and 24 h) were determined. In all of the populations, the greatest amylase activity was observed in the group of insects exposed to the insecticides for 12 h. The population from Currais showed greater lipase activity in the treatment with 1 h of exposure to deltamethrin, whereas the population from Colônia showed the lowest lipase activity in the treatment with 24 h of exposure.
Key words: Deltamethrin, enzyme activities, amylase, lipase, protease.
The maize weevil, Sitophilus zeamais Motsch. (Coleoptera: Curculionidae), is one of the most important pests of stored grains in Brazil and is well-distributed globally. The primary control of this insect depends directly on chemical insecticides. However, the intensive use of insecticides has resulted in the emergence of generalised resistance in insects of this species, which has caused serious problems related to their control (Suthisut et al., 2011; Ishii et al., 2010; Araújo et al., 2011).
Metabolism or detoxification are the most likely mechanisms of resistance in insects to the most frequently studied insecticides. These mechanisms are performed through enzymes such as hydrolases, monooxygenases that are dependent on cytochrome P-450, and transferases, which allow the insect to detoxify insecticides at a sufficient rate to prevent action at the target site. These enzymes have been studied primarily in insects because of the important metabolic functions that they perform in various representatives of this class, such as the control of juvenile hormone levels and regulation of reproductive mechanisms, digestive processes and biochemical mechanisms that confer resistance to organophosphoric, carbamate and pyrethroid insecticides.
Lucena et al. (2012) analysed esterases during the larval, pupal, and adult phases in Sitophilus oryzae (Linné, 1763) (Coleoptera: Curculionidae) and their relationship with resistance to the insecticide Malathion, and they concluded that alterations observed in four esterase loci are of great importance in the development of resistance in insects exposed to this insecticide.
The production of genes related to the resistance of a specific xenobiotic implies an adaptive cost, inferring that the initial fitness cost increases gradually because of later mutations. As a result, an increased metabolic rate may be necessary for individuals with resistance to maintain the resistance mechanism, thus destabilising their gaseous exchanges. If an increase does not occur in energy metabolism, the reallocation of energy can cause a decrease in other physiological processes involved in insect development and reproduction (Coustau et al., 2000). A higher quantity of stored reserve molecules can increase the provision of energy for the production of detoxifying apparatuses without reducing the energy used for physiological processes.
Protein degradation is associated with elevated protease activity, which contributes to increases in free amino acids in the haemolymph. Under stress conditions, these free amino acids serve as precursors for the synthesis of essential constituents in the organism and contribute to compensatory physiological mechanisms against the toxicity of insecticides by providing intermediates for the Krebs cycle and homeostasis maintenance in the organism (Ahmed et al., 1998).
Specific changes in protease activity may be related to the development of resistant groups (Zhu and Baker, 1999). Thus, characterising protease activity is relevant in the context of resistance mechanisms of insects because of its direct relationship with metabolism and energy source allocation. This allocated energy is used in physiological processes and detoxification mechanisms and may be used to supply energy reserves.
Thus, to elucidate the spectrum of proteases present in S. zeamais and their interaction with the resistance mechanism, this study aimed to test for differences in the activity of energy metabolism and digestive enzymes in populations of S. zeamais after exposure to the insecticide compound.
The populations of S. zeamais were acquired from small farms located in southern Piauí (Vale do Gurgueia), including Palmeira, Colônia, Currais and Bom Jesus. The insects were transported to the zoology laboratory and stored in plastic flasks (approximately 25 cm in length and 15 cm in diameter) containing corn that was free of insecticides and pest insects; the flasks were then sealed with a voile-type fabric to permit air circulation and prevent the escape of insects and entry of possible predators or parasites. The insecticide used to establish the dose-response concentration was K-Obiol (Deltamethrin) which is a specific insecticide for the treatment of stored grains.
Bioassay of the response concentration
Initially, preliminary assays were performed to determine the insecticide concentration ranges that caused mortality greater than zero and less than 100%. Based on this information, seven concentrations were established for the treatments of insects in definitive exposure bioassays.
The K-Obiol concentrations ranged from 3.5 x 10-4 to 0.05 g i.a/cm2. Five replicates containing 20 adult unsexed S. zeamais individuals were used per concentration, for a total of 100 insects/ concentration. In each replicate, the inside of the 20 ml glass flask was covered with 200 μl insecticide dissolved in acetone, which was distributed uniformly on the entire internal surface of the flask. For each concentration, a control containing only the solvent acetone was also prepared. After complete evaporation of the solvent, the 20 insects were released into the flask.
To evaluate mortality, the opening of each glass was capped to avoid the escape of insects and those that were incapable of walking after 48 h exposure to the insecticide residues were considered dead.
Bioassays of exposure to the insecticide compounds
From the LC50 values calculated for each population, the insects were subjected to exposure to residues of the insecticide K-Obiol for different time periods: 1, 6, 12 and 24 h. The bioassays were conducted in an entirely random experimental setup, with four replicates. To verify whether the activity of amylase, lipase, serine and cysteine proteases was altered, the insects from each insecticide exposure time period were used in the bioassays of enzymatic activity.
Enzyme preparation
For the amylase, lipase and total protease assay, a BIOCLIN kit (QUIBASA – QuímicaBásica Ltda., Belo Horizonte, Minas Gerais, Brazil) was used.Twenty unsexed adult insects from each population from each treatment (time of exposure to the insecticide) were washed separately in a 1.5% KCl solution, macerated and homogenised in 5.0 ml distilled water at pH 3.0. The extract was filtered on cotton gauze and centrifuged at 3500 gmax for 30 min. The aliquots of the supernatant were removed to determine the concentration of proteins, and the pellet was discarded. A water bath at 37°C was used to evaluate the enzyme activity. The kinetic parameters of the amylase, lipase, serine and cysteine protease were analysed at the following wavelengths: 405, 410, 260 and 545 nm, respective. The readings were performed using a QUIMIS spectrophotometer (UV-VIS Double Beam, Model Q798UV-DB).
Statistical analyses
The mortality results were subjected to a probit analysis using the PROC PROBIT procedure of the Statistical Analyses System (SAS) (SAS Institute, 2002), which generated the concentration x mortality curves. The mortality data were corrected using the control mortality. The data for the enzymatic activities were subjected to an analysis of variance (ANOVA) followed by a means comparison using Tukey's test at 5% probability.
Bioassay of the K-Obiol concentration-response
In bioassays for the detection of resistance in populations of
S. zeamais, the

value was between 27.52 and 79.07, the
p-value ranged from 0.19 to <0.001, and the slope ranged from 1.23 to 2.00. The experiments performed with K-Obiol for the populations from Currais, Palmeira, Bom Jesus and Colônia had different LC
50 values (Table 1).
The population with the highest LC50 value (94.0 µg i.a/cm2) was from Currais, and the population with the lowest value was from Bom Jesus at 14.0 µg i.a/cm2. These populations were maintained in the laboratory for more than two years, and elevated concentrations of deltamethrin are required to produce LC50 values in the studied populations. The large-scale use of insecticides has occurred in the sampled municipalities since the 1990s.
Enzyme activities
The specific activities of the energy metabolism enzymes lipase and amylase, which are responsible for the hydrolysis of sugars, were determined in four populations of S. zeamais exposed to the insecticide K-Obiol for different times periods: 1, 6, 12 and 24 h (Table 2). Factors such as fasting, pre-reproduction, drug exposure and hormonal regulation were not considered. According to the results, lipase and amylase enzymes showed a significant difference in their activities among the four populations exposed to deltamethrin for different amounts of time.
For the energy metabolism enzymes amylase and lipase, differences were not observed between the populations in the control treatment. For all of the populations, there was greater amylase activity in the group of insects exposed for 12 h to the insecticides. Thus, amylase may be directly connected to the enzymes involved in energy metabolism and act as a provider of sugars for metabolism, which is directly related to growth (Table 2).
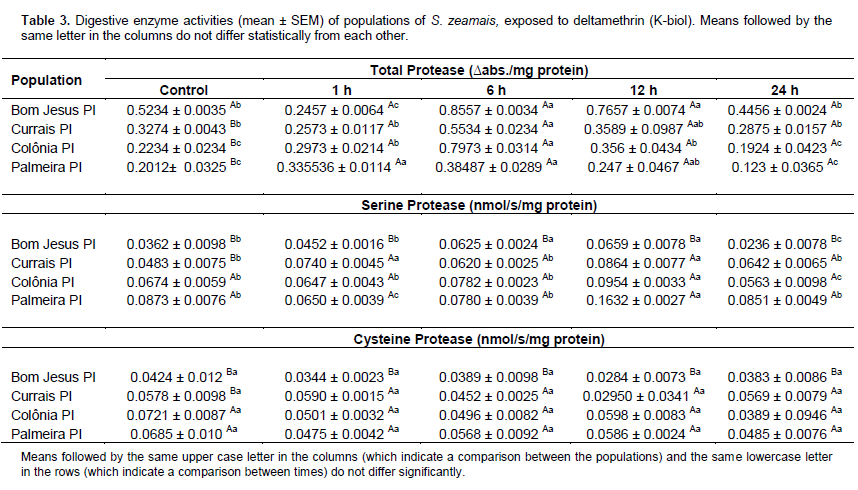
The population from Currais showed the highest lipase activity in the treatment exposed to deltamethrin for 1 h, whereas the population from Colônia showed the lowest lipase activity in the treatment exposed to deltamethrin for 24 h. According to the observations, the pattern of enzymatic activity is altered as a function of the treatments, population and exposure time. After 24 h of exposure to deltamethrin, the population from Currais showed higher amylase and lipase activity relative to the other populations, which may be directly related to resistance because in the dose response concentrations of this same experiment, this population had the highest LC50. These findings show that these insects increase their metabolism over time, thereby increasing their defence mechanisms and conferring resistance.Specific activities of digestive system enzymes were determined (total protease, serine and cysteine protease) in the four populations of S. zeamais (Table 3).

According to the results of the ANOVA and subsequent means testing, only the activity of total protease did not differ significantly between the studied populations. The population of Bom Jesus showed a significant difference with the other populations and had the lowest activities of serine protease and cysteine protease. In contrast, the highest activity of serine protease was only observed in the population of Palmeira PI after 12 h of exposure to deltamethrin.
The activity of specific classes of proteases (serine proteases and cysteine proteases) indicated that there was a significant difference among the exposure times to the residues of deltamethrin.
Bioassay of the K-Obiol concentration response
Genetic modifications that allow an individual to survive in a new environment, such as grains treated with insecticides in the case of weevils and beetles, are normally associated with an adaptive cost because any adaptation to a new condition (such as tolerating doses of insecticide that would be lethal to the majority of individuals) involves ancestral modifications. In this context, resistant individuals would be at an advantage in the presence of the insecticide; however, they would be less fit than susceptible insects when the insecticide is not used. However, studies have demonstrated the absence of the physiological costs associated with resistance. For example, certain resistant populations of S. zeamais to pyrethroids showed similar reproductive performance to that of the susceptible population, suggesting that resistance to the insecticide is fixed in the population and the expression of the genes responsible for this characteristic no longer incur a cost for the individuals (Guedes et al., 2006). Thus, it is not expected that susceptibility will be re-established in situations where a specific insecticide has not been used for a certain period of time, which explains the results obtained in the bioassays to determine the mortality concentration.
The results of this study are consistent with those of the study by (Silva et al., 2013), who performed an experiment that evaluated dose responses with the insecticide permethrin in populations from southern Piauí and observed high resistance relative to the population from Sete Lagoas, Minas Gerais, which is the standard for susceptibility as described in the studies by Guedes et al. (2006) and Braga et al. (2011).
This resistance may be directly related to the large-scale use of this insecticide in the region since the 1990s because of the large soy plantations in the Cerrado, which may have led to the selection of these insects and their presence at the small farms. The propagation of resistance to insecticides in insect populations is related to the frequency of insecticide use and a result of selective pressure by these toxic compounds on the population as well as inherited characteristics of the species of insects involved.
Enzyme activities
Insecticide degradation occurs through various metabolic processes in which the product is transformed into a form that does not harm the insect or that can be rapidly eliminated from the body. Various enzymes are involved in this process, such as esterases, oxidases, transferases and others that can increase their efficiency in the resistant populations (Beckel et al., 2006).
Corrêa et al. (2011) analysed the enzymatic activity in populations of S. zeamais and obtained results with muchlarger variations than those obtained in this experiment. One of the populations with large amylase production is from Juiz de Fora, and this increased production may be directly related to the resistance of this population as reported in the studies by Pereira et al. (2009). Mujeeb and Shakoori (2012) analysed the effect of the pyrethroid Fury (S-cyano (2-phenoxyphenyl (±)-cis/trans 3-(2,2-dichloroethenyl)-2,2-dimethylcyclopropane carboxylate) on esterases at different stages of development of Tribolium castaneum and observed that significant changes had not occurred on the insect at 15 days of age. Thus, this is one of the resistance mechanisms of pest insects when the entire body of the insect is already sclerotic.
In the insects, the fatty body is the primary location for synthesis and storage of carbohydrates, lipids and proteins, which are readily used as an energy source for a wide range of activities. Carbohydrates are stored primarily in the form of trehalose and glycogen, and lipids are stored in the form of diacylglycerols or triacylglycerols.
Differences in fatty body morphology and average respiratory rate were reported in a study performed by Guedes et al. (2006) in populations of S. zeamais with distinct susceptibility to pyrethroid insecticides. These cytomorphological patterns in cells of the fatty body of the resistant populations suggest a greater availability of total proteins and neutral carbohydrates in insects from these populations.
The greater accumulation most likely contributes to resistance in insects with a greater capacity for mobilising reserves; thus, these insects can supply the energetic demands required to activate their resistance mecha-nisms to insecticides and resist the toxic action of these compounds, which may be associated with mitigating the adaptive costs of the resistance phenomenon in the absence of insecticides.
The maintenance of mechanisms that confer resistance to insecticide molecules may demand an additional and sufficient quantity of energy to confer a disadvantage to the resistant phenotypes relative to the susceptible phenotypes (Boivin et al., 2003).
According to the results, the lipase and amylase enzymes showed significant differences in their activities between the four populations at different times of expo-sure to deltamethrin. The enzymatic activity of the lipase enzyme was highest in the population with the highest LC50 value (Currais), which had 6 hours of exposure to the insecticide, whereas amylase showed the lowest activity at this time.
In this study, the production of lipase was higher on average at the first time point. Relative to the control, which contained only the solvent acetone, lipase production at 1 h of exposure was more than four times greater, which indicated that when subjected to an unfavourable condition, physiological metabolism acts immediately to combat the foreign body. According to Atella et al. (2012), lipids perform various functions, including acting as hormones and important energy reserves that are essential in situations that require great metabolic demand, such as those detailed in this study in which insects were subjected to doses of the insecticide deltamethrin.
Amylase activity was greater at 12 h in all of the populations, and according to Mehrabadi et al. (2011), the presence of a large number of amylase enzymes in insects is an efficient strategy to escape toxicities. Amylases constitute a family of endoamylases that catalyse the hydrolysis of α-1,4 glycosidic bonds in starch, glycogen and other carbohydrates; therefore, these enzymes are important for the development and survival of larvae and/or adults, including S. zeamais.
This study’s results for protease activity are similar to the results for amylase and lipase, and the populations showed differentiated enzymatic activity as a function of the treatments. In insects, it is known that amylases are synthesised and segregated along with other digestive enzymes by epithelial cells of the middle intestine (Sousa and Conte, 2013).
It is reported in literature that on the middle intestine of pest insects have been performed to improve our understanding of resistance mechanisms.
According to the results of this study, we suggest that the efficient energetic metabolism observed in the insect population from Currais can mitigate the physiological cost associated with resistance in this population (exposure to the highest doses of the insecticide). It is believed that insects from this population allocate their energetic reserves in a balanced manner when required by the organism. Thus, the defence mechanism appears to be active only when required.
Proteases are an important group of hydrolytic enzymes in insects because they are involved in digestive processes, pro-enzyme activation, physiologically active peptide release, complement system activation and inflammation processes. Proteases are classified accor-ding to their mechanism of catalysis: serine proteases, cysteine proteases, aspartic proteases, and metalloproteinases (Macedo and Freire, 2011; Wang et al., 2014).
Serine proteases are proteolytic enzymes dominant in the middle intestine of the insect and can be distinguished by the presence of the Asp-His-Ser “charge relay” or “catalytic triad” system. These proteases are present in all of the phylogenetic kingdoms, including viruses, and involved in many physiological processes. Trypsins and chymotrypsins have a particularly important role in the digestion of foods by insects as well as in defence and immunity (Hou et al., 2014; Wang et al., 2014). In higher organisms, approximately 2% of the organism's genes encode these enzymes (Oliveira and Macedo, 2011).
Chen et al. (2013) analysed the genes of proteases, such as serine and cysteine protease, in the genome of the Hessian fly as well as their interactions with the genotypes of host plants, and they identified 105 genes for cysteine and serine proteases. These genes were named mayetiola destructor protease (MDP1 to MDP105). An enzymatic analysis of the active sites of the predicted proteins revealed that 94 are protease proteins (such as serine) and the remaining 11 proteins are proteases (such as cysteine).Cysteine proteases hydrolyse endopeptidyl bonds with a cysteine residue in the active site and are normally identified based on the effect of their active-site inhibitors (Grudkowska and Zagdańska, 2004). In insects, cysteine proteases are used in digestive processes, but found in various tissues, which indicates that they can also perform other functions (Macedo and Freire, 2011), such as the promotion of resistance to insecticides.
Mendonça et al. (2011) performed an enzymatic characterisation of the isoforms of cysteine protease from Anticarsia gemmatalis (HÜBNER, 1818) (Lepidoptera: Noctuidae) and found cysteine proteases in the middle intestine of this pest insect that presented activities at different pH values. These enzymes aid in the protein digestive process in the insect and can serve as targets for the control of agricultural pests, for insects with this characteristic has a greater amount of free amino acids in the hemolymph, enabling greater detoxificativas amplification enzymes.
Although these insects had been maintained in the laboratory for a significant amount of time, elevated concentrations of insecticides were still necessary because the analysed populations presented acute resistance. A significant difference was observed in the activities of specific classes of proteases (serine proteases and cysteine proteases) among the times of exposure to the deltamethrin residues. The population from Currais may present the greatest efficiency in mitigating the physiological course of this insecticide and is considered the most resistant.
The authors have not declared any conflict of interest.
The authors would like to thank the CNPq for providing a grant and the CPCE for providing the opportunity to conduct this research.
REFERENCES
|
Ahmed S, Wilkins RM, Mantle D (1998). Comparison of proteolytic enzyme activities in adults of insecticide resistant and susceptible strains of the housefly Musca domestica L. Insect Biochem. Mol. Biol. 28:629-639.
Crossref
|
|
|
|
Araújo RA, Williamson MS, Christopher B, Field LM, Duce IR (2011). Pyrethroid resistance in Sitophilus zeamais is associated with a mutation (T929I) in the voltage-gated sodium channel. Insect Mol. Biol. 204:437-445.
Crossref
|
|
|
|
|
Atella GC, Majerowicz D, Gondim KC (2012). Lipid Metabolism. National Institute of Science and Technology in Molecular Entomology. 22p.
|
|
|
|
|
Beckel HS, Lorini I, Lazzari SMN (2006). Efeito do sinergista butóxido de piperonila na resistência de Oryzaephilus surinamensis (L.) (Coleoptera: Silvanidae) a deltametrina e fenitrotiom [Synergistic effect of piperonyl butoxide in the resistance of Oryzaephilus surinamensis (L.) (Coleoptera: Silvanidae) to deltamethrin and phenitrothion]. Rev. Bras. Entomol. 501:110-114.
Crossref
|
|
|
|
|
Boivin T, Bouvier JC, Beslay D, Sauphanor B (2003). Phenological segregation of insecticide resistance alleles in the codling moth Cydia pomonella (Lepidoptera: Tortricidae): a case study of ecological divergences associated with adaptive changes in populations. Gene Res. 81:169-177.
Crossref
|
|
|
|
|
Braga LS, Corrêa AS, Pereira EJG, Guedes RNC (2011). Face or flee? fenitrontion resistance and behavioral response in populations of the maize weevil, Sitophilus zeamais. J. Stored Prod. Res. 47:161-167.
Crossref
|
|
|
|
|
Corrêa AS, Pereira EJG, Cordeiro EMG, Braga LS, Guedes RNC (2011). Insecticide resistance, mixture potentiation and fitness in populations of the maize weevil (Sitophilus zeamais). Crop Protect. 30:1655-1666.
Crossref
|
|
|
|
|
Coustau C, Chevillon C, Constant FR (2000). Resistance to xenobiotic and parasites: can we count the cost? Trends Ecol. Evol. 15:378-383.
Crossref
|
|
|
|
|
Chen H, Zhu YC, Whitworth RJ, Reese JC, Chen MS (2013). Serine and cysteine protease-like genes in the genome of a gall midge and their interactions with host plant genotypes. Insect Biochem. Mol. Biol. 438:701-711.
Crossref
|
|
|
|
|
Grudkowska M, Zagdańska B (2004). Multifunctional role of plant cysteine proteinases. Acta Biochim. Pol. 51:609-624.
PMid:15448724
|
|
|
|
|
Guedes RNC, Oliveira EE, Guedes NMP, Ribeiro BM, Serrão JE (2006). Cost and mitigation of insecticide resistance in the maize weevil, Sitophilus zeamais. Physiol. Entomol. 31:30-38.
Crossref
|
|
|
|
|
Hou MZ, Shen GM, Wei D, Li YL, Dou W, Wang JJ (2014). Characterization of Bactrocera dorsalis serine proteases and evidence for their indirect role in insecticide tolerance. Int. J. Mol. Sci. 152:3272-3286.
Crossref
|
|
|
|
|
Ishii t, Matsuzawa H, Vairappan CS (2010). Repellent activity of common spices against the rice weevil, Sitophilus zeamais Motsch (Coleoptera, Curculionidae). J. Trop. Biol. Conserv. 7:75-80.
|
|
|
|
|
Lucena ALM, Gigliolli AAS, Lapenta AS (2012). Analysis of the esterases during the development phases in Sitophilus Oryzae (Coleoptera: Curculionidae) and their relationship with resistance to the insecticide malathion. Revista de Saúde e Biologia. 73:36-44.
|
|
|
|
|
Macedo MR, Freire MGM (2011). Insect digestive enzymes as a target for pest control. Invertebrate Survival J. 8:190-198.
|
|
|
|
|
Mehrabadi M, Bandani AR, Saadati F, Mahmudvand M. (2011). Amylase activity of stored products insects and its inhibition by medicinal plant extracts. J. Agric. Sci. Technol. 13:1173-1182.
|
|
|
|
|
Mendonça EG, Visôtto LE, Costa NCS, Ribeiro FR, Oliveira JA, Oliveira MGA (2011). Enzymatic characterization of isoforms of cysteine protease from Anticarsia gemmatalis (HÜBNER, 1818). Ciência e Agrotecnologia. 353:446-454.
Crossref
|
|
|
|
|
Mujeeb KA, Shakoori AR (2012). Effect of Fury, a synthetic pyrethroid, on esterases of different developmental stages of stored grain pest, red flour beetle, Tribolium castaneum (Herbst.) – spectrophotometric analysis. J. Zool. 443:601-613.
|
|
|
|
|
Oliveira CFR de, Macedo MLR (2011). Use of protease inhibitors Vegetables Alternatively Biotechnology tool not Pest Control. Biol. Saúde 1:1-11.
|
|
|
|
|
Pereira CJ, Pereira EJG, Cordeiro EMG, Lucia TMCD, Tótola MR, Guedes RNC (2009). Organophosphate resistance in maize weevil Sitophilus zeamais: magnitude and behavior. Crop Prot. 28:168-173.
Crossref
|
|
|
|
|
SAS Institute (2002). SAS/STAT User’s Guide, version 8. SAS Institute, Cary.
|
|
|
|
|
Silva LB, Silva LS, Mancin AC, Carvalho GS, Silva JC, Andrade LH (2013). Behaviour of the corn weevil challenged with doses of permethrin. Comunicata Scientiae. 41:26-34.
|
|
|
|
|
Sousa GE, Conte H (2013). Midgut morphophysiology in Sitophilus zeamais Motschulsky, 1855 (Coleoptera: Curculionidae). Micron 51:1-8.
Crossref
|
|
|
|
|
Suthisut D, Fields O, Chandrapatya A (2011). Fumigant toxicity of essential oils from three Thai plants (Zingiberaceae) and their major compounds against Sitophilus zeamais, Tribolium castaneum and two parasitoids. J. Stored Prod. Res. 47:222-230.
Crossref
|
|
|
|
|
Wang C, Wang L, Su B, Lu N, Song J, Yang X, Fu W, Tan W, Han B (2014). Serine protease inhibitor Kazal type 1 promotes epithelial-mesenchymal transition through EGFR signaling pathway in prostate cancer. Prostate 74:689-701.
Crossref
|
|
|
|
|
Zhu YC, Baker JE (1999). Characterization of midgut trypsin-like enzymes and three trypsinogen DNA from the lesser grain borer, Rhyzopertha dominica (Coleoptera: Bostrichidae). Insect Biochem. Mol. Biol. 29:1053-1063.
Crossref
|
|