Full Length Research Paper

ABSTRACT
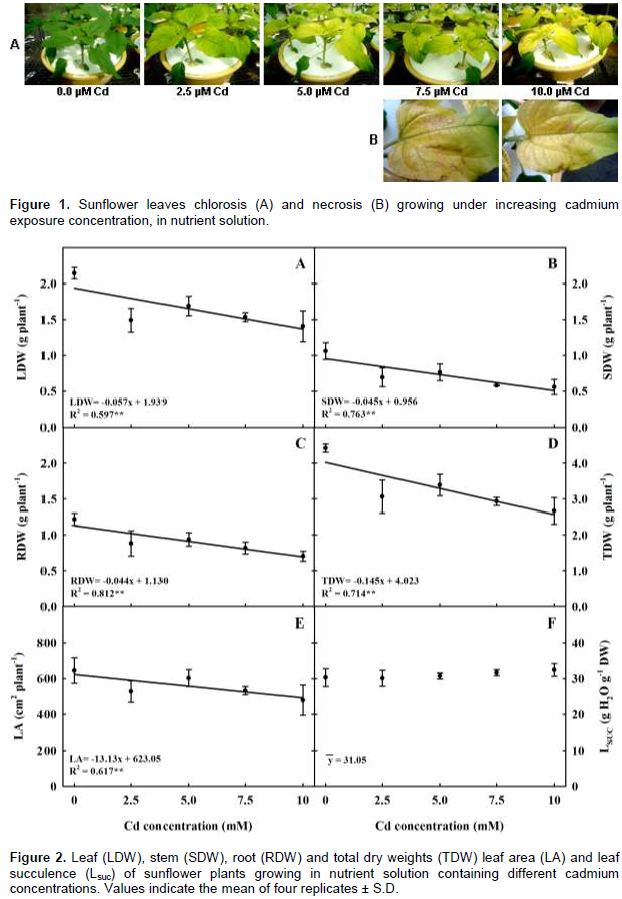
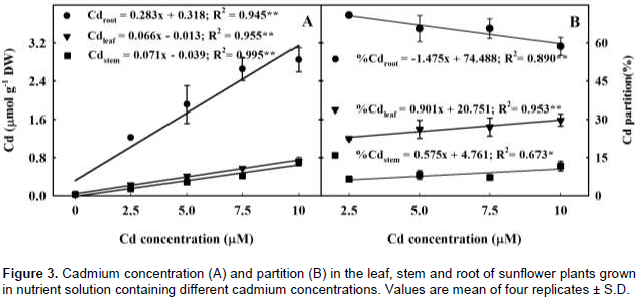
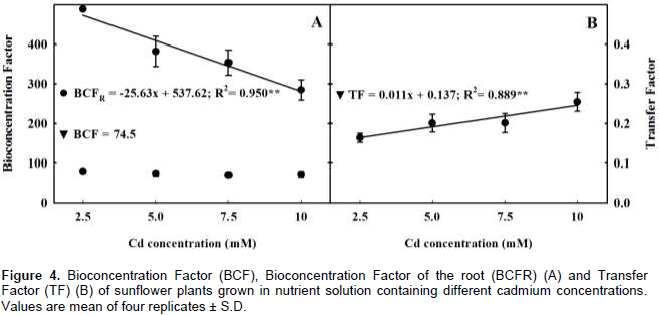
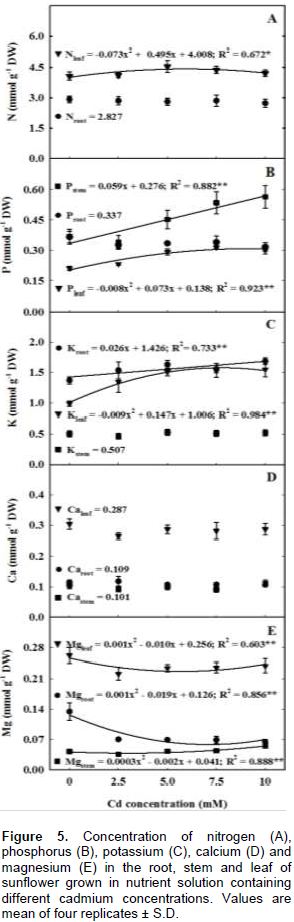
A hydroponic greenhouse study was carried out to evaluate the effects of increasing cadmium (Cd) concentration on plant growth, mineral nutrition and Cd distribution of H-250 sunflower genotype. Exposure to increasing Cd concentrations reduced plant biomass by 40, 34, 47 and 42% of the total, leaves, stem and roots dry weights as compared to the control. Regardless of the treatment most of Cd uptake by the genotype was allocated in the root, followed by leaf and stem. The higher bioconcentration factors values in both above ground and underground plant tissues and low transfer factor value indicated that this genotype may be an alternative for use in phytostabilization programs. The results also showed that increasing Cd concentration disrupted plant homeostasis as it increased the concentration of some nutrients and had adverse effect on others, impacting plant growth. In this context, the results suggest that the low magnesium, iron and manganese concentrations in the leaves were the main cause for plant biomass reduction and leaf chlorosis and necrosis, as each one of these elements plays a key role on the chlorophyll molecule and on photosynthesis process.
Key words: Bioremediation, bioaccumulation factor, heavy metal stress, Helianthus annuus, transfer factor.
INTRODUCTION
MATERIALS AND METHODS
RESULTS AND DISCUSSION
Potassium plays a central role in the water relations (Meurer, 2006), therefore the increase in leaf and root K concentrations is consistent with the need of the plant to keep the turgor, mainly in higher Cd concentrations. The increasing K concentration in sunflower roots may have helped the maintenance of water absorption by the roots and its flux to the shoot. Additionally, the increasing K concentration in the leaves may have favored the stomata opening at higher Cd concentrations, thereby maintaining the plant gas exchange during stress. The increase in K concentration in sunflower leaves obtained in this study differed from those obtained for sunflower, (Simon, 1998), cedar (Paiva et al., 2001) and maize roots, (Ciećko et al., 2004). However, the Cd concentration in the solution, the time of the exposure to Cd, and the plant age of these studies were higher than that tested in our study.
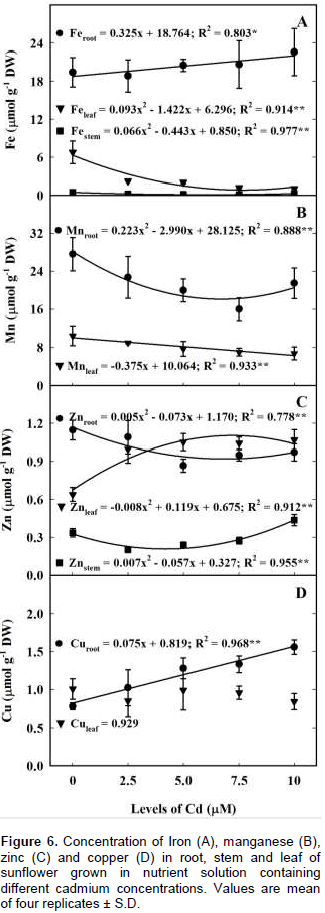
The concentration of Cd-solution has no effect in the Ca concentration in the root, stem and leaf (Figure 5D). In contrast, increasing Cd concentration has negative quadratic effect in plant Mg (Figure 5E). The estimated minimum concentration in leaf (0.231 mmol g-1 DM), stem (0.038 mmol g-1 DM) and root (0.036 mmol g-1 DM) occurred, respectively, at Cd5.0, Cd3.3 and Cd9.5.Iron, Mn, Zn, and Cu concentrations in sunflower leaves and roots are shown in Figure 6. Cadmium concentrations had positive effect on sunflower root Fe and Cu and leaf Zn but negative effect on leaf and stem Fe, leaf and root Mn, and stem and root Zn (Figure 6). The estimated minimum leaf Fe concentration (0.86 μM g-1 DW) was at Cd7.65 μM Cd, representing a reduction of 87% when compared with Fe concentration of the control (6.75 μM g-1 DW) (Figure 6A). Leaf Mn concentration decreased linearly (0.375 μM g-1 DW) with Cd solution (Figure 6B). Leaf Fe and Mn concentrations were present in concentration reverse to that of Cd. Sunflower leaf Zn concentration increased with Cd-solution (Figure 6C). The estimated maximum leaf Zn concentration 1.12 μM g-1 DW occurred at Cd7.4. Leaf Cu concentration had no change (0.929 μM g-1 DW) with Cd-solution (Figure 6D). The concentrations of Mn and Cu in the stem were below the level of detection. Sunflower root concentration of Fe and Cu increased in 0.325 and 0.075 μmol g-1 DW, respectively for each unit of Cd-solution (Figures 6A and 6D). At Cd10 the root Cu concentration, 1.57 μmol g-1 DW, was twice the value of the control plants 0.819 μmol g-1 DW. In contrast, the root concentration of Mn and Zn decreased quadratically with Cd-solution (Figures 6B and 6C). The minimum Mn 18.10 μM g-1 DW and Zn 0.90 μM g-1 DW concentrations were observed at Cd6.7 and Cd7.3, respectively.
CONCLUSIONS
CONFLICT OF INTERESTS
ACKNOWLEDGEMENTS
REFERENCES
|
Ansari R, Kazi TG, Jamali MK, Arain MB, Wagan MD, Jalbani N, Afridi HI, Shah AQ (2009) Variation in accumulation of heavy metals in different verities of sunflower seed oil with the aid of multivariate technique. Food Chem. 115(1):318-323. |
|
|
ATSDR - Agency for Toxic Substances and Disease Registry (2014). Priority List of Hazardous Substances. Available at: http://www.atsdr.cdc.gov/SPL/index.html. Accessed 16 May 2015. |
|
|
Azad HN, Shiva AH, Malekpour R (2011). Toxic effects of lead on growth and some biochemical and ionic parameters of sunflower (Helianthus annuus L.) seedlings. Curr. Res. J. Biol. Sci. 3(4):398-403. |
|
|
Belleghem FV, Cuypers A, Semane B, Smeets K, Vangronsveld J, d'Haen J, Valcke R. (2006). Subcellular localization of cadmium in roots and leaves of Arabidopsis thaliana. New Phytol. 173(3):495-508. |
|
|
Benavides MP, Gallego SM, Tomaro ML (2005). Cadmium toxicity in plants. Braz. J. Plant Physiol. 17(1):21-34. |
|
|
Capone A, Santos ER, Ferraz EC, Santos AF, Oliveira JL, Barros HB (2012). Desempenho agronômico de cultivares de girassol no sul do Estado de Tocantins. J. Biotec. Biodivers. 3(3):13-23. |
|
|
Chaves LHG, Estrela MA, Souza RS (2011). Effect on plant growth and heavy metal accumulation by sunflower. J. Phytol. 3(12):4-9. |
|
|
Ciećko Z, Kalembasa S, Wyszkowski M, Rolka E (2004). Effects of soil contamination by cadmium on potassium uptake by plants. Polish J. Environ. Stud. 13(3):333-337. |
|
|
Clemens S (2006). Toxic metal accumulation, responses to exposure and mechanisms of tolerance in plants. Biochim. 88(11):1707-1719. |
|
|
Di Toppi LS, Gabbrielli R (1999). Response to cadmium in higher plants. Environ. Exp. Bot. 41:105-130. |
|
|
Dong J, Mao WH, Zhang GP, Wu FB, Cail Y (2007). Root excretion and plant tolerance to cadmium toxicity – a review. Plant Soil Environ. 53(5):193-200. |
|
|
Faithfull NT (2002). Methods in agricultural chemical analysis: A practical handbook. CABI Publishing, Wallingford. |
|
|
FAO/WHO (1972). Evaluation of certain food additives and the contaminants mercury, lead, and cadmium. Sixteenth Report. FAO Nutrition Meetings Report Series No.51. WHO Technical Report Series No. 505. World Health Organization, Geneva. |
|
|
Gajdos E, Lévai L, Veres S, Kovács B (2012). Effects of biofertilizers on maize and sunflower seedlings under cadmium stress. Commun. Soil Sci. Plant Anal. 43(1-2):272-279. |
|
|
Goussias C, Boussac A, Rutherford W (2002). Photosystem II and photosynthetic oxidation of water: an overview. Phil. Trans. R. Soc. Lond. B 357(1426):1369-1381. |
|
|
Guimarães MA, Santana TA, Silva EV, Zenzen IL, Loureiro EM (2008). Toxicidade e tolerância ao cádmio em plantas. Rev. Tróp. 1(3):58-68. |
|
|
Higuchi K, Tsuchiya M, Nakata S, Tanabe A, Fukawa S, Kanai M, Miwa E (2013). Detoxification of cadmium (Cd) by a novel Cd-associated and Cd-induced molecule in the stem of common reed. J. Plant Physiol. 170(18):1553-1560. |
|
|
Hoagland DR, Arnon DI (1950). The water-cultured method for growing plants without soil. Calif. Agr. Expt. Sta. Circ. 347(1):1-32. |
|
|
Hossain MA, Hasanuzzaman M, Fujita M (2010). Up-regulation of antioxidant and glyoxalase systems by exogenous glycinebetaine and proline in mung bean confer tolerance to cadmium stress. Physiol. Mol. Biol. Plant 16(3):259-272. |
|
|
Järup L, Åkesson A (2009). Current status of cadmium as an environmental health problem. Toxicol. Appl. Pharmacol. 238:201-208. |
|
|
Jones JB (2001). Laboratory guide for conducting soil tests and plant analysis. CRC Press, Boca Raton 363 p. |
|
|
Lopes Júnior CA, Mazzafera, P, Arruda, MAZ (2014). A comparative ionomic approach focusing on cadmium effects in sunflowers (Helianthus annuus L.). Environ. Exp. Bot. 107(11):180-186. |
|
|
López-Millán A, Sagardoy R, Solanas M, Abadía A, Abadía J (2009). Cadmium toxicity in tomato (Lycopersicon esculentum) plants grown in hydroponics. Environ. Exp. Bot. 65(2-3):376-385. |
|
|
Lux A, Martinka M, Vaculík M, White PJ (2011). Root responses to cadmium in the rhizosphere: a review. J. Exp. Bot. 62(1):21-37. |
|
|
Malik RN, Husain SZ, Nazir I (2010). Heavy metal contamination and accumulation in soil and wild plant species from industrial area of Islamabad, Pakistan. Pak. J. Bot. 42(1):291-301. |
|
|
Mantovani A (1999). A method to improve leaf succulence quantification. Braz. Arch. Biol. Technol. 42:9-14. |
|
|
Marschner P (2012). Marschner's Mineral Nutrition of Higher Plants. Academic Press, London 651p. |
|
|
Mason RP (2013). Trace metals in aquatic systems. John Wiley & Sons, Chichester 440 p. |
|
|
Meurer EJ (2006). Potássio. In: Fernandes MS (Ed.) Nutrição mineral de plantas. Sociedade Brasileira de Ciência do Solo, Viçosa pp. 282-298. |
|
|
Mobin M, Khan NA (2007). Photosynthetic activity, pigment composition and antioxidative response of two mustard (Brassica juncea) cultivars differing in photosynthetic capacity subjected to cadmium stress. J. Plant. Physiol. 164(5):601-610. |
|
|
Nazar R, Iqbal N, Masood A, Khan MIR, Syeed S, Khan NA (2012). Cadmium toxicity in plants and role of mineral nutrients in its alleviation. Am. J. Plant Sci. 3(10):1476-1489. |
|
|
Nickelsen J, Rengstl B (2013). Assembly of photosystem II: from cyanobacteria to plants. Annu. Rev. Plant Biol. 64:609-635. |
|
|
Nocito FF, Lancilli C, Dendena B, Lucchini B, Sacchi G (2011). Cadmium retention in rice roots is influenced by cadmium availability chelation and translocation. Plant Cell Environ. 34(6):994-1008. |
|
|
Obata H, Umebayashi M (1997). Effects of cadmium on mineral nutrient concentrations in plants differing tolerance for cadmium. J. Plant Nutr. 20(1):97-105. |
|
|
Ouariti O, Boussama N, Zarrouk M, Cherif A, Ghorbal MH (1997). Cadmium- and copper-induced changes in tomato membrane lipids. Phytochemical 45(7):1343-1350. |
|
|
Paiva HN, Carvalho JG, Siqueira JO (2001). Effect of the cadmium application on nutrients content in cedro (Cedrela fissilis VELL.) seedlings. Cienc. Florest. 11(2):153-162. |
|
|
Pál M, Horváth E, Janda T, Páldi E, Szalai G (2006). Physiological changes and defense mechanisms induced by cadmium stres in maize. J. Plant Nutr. Soil Sci. 169(2):239-246. |
|
|
Redjala T, Sterckeman T, Morel JL (2009). Cadmium uptake by roots: Contribution of apoplast and of high- and low-affinity membrane transport systems. Environ. Exp. Bot. 67(1):235-242. |
|
|
Sandalio LM, Dalurzo HC, Gómez M, Romero-Puertas MC, Del Río LA (2001). Cadmium-induced changes in the growth and oxidative metabolism of pea plants. J. Exp. Bot. 52(364):2115-2126. |
|
|
Sarwar N, Saifullah, Malhi SS, Zia MH, Naeen A, Bibi S, Farid G (2010). Role of mineral nutrition in minimizing cadmium accumulation by plants (Review). J. Sci. Food Agric. 90(6):925-937. |
|
|
SAS - Statistical Analysis System (2003). Release 9.1 (software). SAS Institute Inc., Cary. |
|
|
Shi G, Liu C, Cai Q, Liu Q, Hou C (2010). Cadmium accumulation and tolerance of two sunflower cultivars in relation to photosynthesis and antioxidative enzymes. Bull. Environ. Contam. Toxicol. 85:256-263. |
|
|
Silva EC, Albuquerque MB, Azevedo Neto AD, Silva Junior CD (2013). Drought and its consequences to plants from individual to ecosystem. In: Akinci Åž (Ed.) Responses of Organisms to Water Stress. InTech, Rijeka pp. 17-47. |
|
|
Simon L (1998). Cadmium accumulation and distribution in sunflower plant. J. Plant Nutrit. 21(2):341-352. |
|
|
Smeets K, Cuypers A, Lambrechts A, Semane B, Hoet P, Laerve AV, Vangronsveld J (2005). Induction of oxidative stress and antioxidative mechanisms in Phaseolus vulgaris after Cd application. J. Plant Physiol. Biochem. 43:437-444. |
|
|
USDA-FAS (2015). Oilseeds: World Markets and Trade. |
|
|
Van Assche F, Clijsters H (1990). Effects of metals on enzyme activity in plants. Plant Cell Environ. 13:195-206. |
|
|
Vassilev A, Yordanov I (1997). Reductive analysis of factors limiting growth of cadmium-treated plants: A Review. Bulg. J. Plant Physiol. 23(3-4):114-133. |
|
|
Yoon J, Cao X, Zhou Q, Ma LQ (2006). Accumulation of Pb, Cu, and Zn in native plants growing on a contaminated Florida site. Sci. Total Environ. 368:456-464. |
|
|
ŽaltauskaitÄ— J, ŠliumpaitÄ— I (2013). Evaluation of Toxic Effects and Bioaccumulation of Cadmium and Copper in Spring Barley (Hordeum vulgare L.). Environ. Res. Eng. Manag. 64(2):51-58. |
|
|
Zou J, Xu P, Lu X, Jiang W, Liu, D (2008). Accumulation of cadmium in three sunflower (Helianthus annuus L.) cultivars. Pak. J. Bot. 40(2):759-765. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


