Full Length Research Paper

ABSTRACT
In vitro cultivation is widely used for micropropagation of several plant species; however, the increase in the gas concentration, such as ethylene gas, inside the growth vials can cause physiological changes. Thus, 4 experiments were carried out at the Federal University of Minas Gerais to evaluate the biochar use from sugarcane bagasse in vials with or without caps with porous membrane in the in vitro growing of pitaya (Hylocereus undatus Haw) (experiment 1) and strawberry (Fragaria x ananassa Duch) (experiment 2) and evaluating in combination of the use of biochar from sugarcane bagasse and AgNO3 in the in vitro growing of pitaya (experiment 3) and strawberry (experiment 4). The types of caps did not affect the growth and number of pitaya and strawberry; however, using caps with porous membrane obtained more magnificent roots dry matter yield. The addition of 4 g L-1 of biochar, regardless of cap type, favored the pitaya and strawberry roots' growth and development. AgNO3 alone contributed to the higher length and dry mass of pitaya roots and greater dry mass of strawberry roots. Adding biochar decreases the AgNO3 effects in vitro culture. Biochar is an alternative to improve the in vitro environment of plant species.
Key words: Fragaria x ananassa Duch, Hylocereus undatus Haw, Tissue culture, AgNO3, gas exchange.
INTRODUCTION
In vitro micro propagation is a rapid technique used to produce large-scale seedlings of various plant species. The vails used are generally sealed, which interferes in gas exchange with the external environment, favors the increase of relative air humidity and concentration of gases such as ethylene, and reduces CO2 and O2 inside the vails. High gas concentrations inside the vails can alter some plant physiological processes, such as multiplication, growth, development, and survival (Naing et al., 2014; Waikhom and Louis, 2014). Gaseous exchange between the vials atmosphere and the outside environments favors the growth and development of plant tissues (explant) during in vitro growth. It improves the plant's adaptation to the acclimatization stage since the gas exchange is directly involved in the quantity and stomatal efficiency and plants' metabolite production (Iarema et al., 2012; Mohamed and Alsadon, 2010; Taiz et al., 2017; Vieira et al., 2019). Some techniques have been used to control gas concentrations and gas exchange in vitro growth, such as caps with a microporous membrane (Mohamed and Alsadon, 2011; Santana et al., 2011). The addition of activated charcoal or AgNO3 is used to decrease the concentration of some undesirable gases as ethylene (Rodrigues et al., 2017). Ethylene gas is a plant hormone that can inhibit plant growth during in vitro growth (Di Lonardo et al., 2013). In addition to allowing gas exchange between the vials and the outside environments, the caps with microporous membranes favor increased CO2 concentration and relative air humidity inside the vials and increase plant transpiration and uptake of water and nutrients (Kozai, 2010; Xiao et al., 2011). Due to these effects, there may be an increase in photosynthetic capacity, multiplication rate, and plants' survival in the acclimatization stage (Hoang et al., 2020). The activated charcoal addition in both liquid and semi-solid media for tissue culture aims to reduce ethylene's concentration inside the vials since activated charcoal particles have surface electric charges that immobilize ethylene and other compounds that can damage plants in vitro growing (George et al., 2008). Besides, the active charcoal can inhibit tissue browning and favor the embryogenesis, androgenesis, rooting, shoot, and root elongation (Thomas, 2008).
Some authors recommend adding AgNO3 to the culture medium instead of activated charcoal. AgNO3 reduces the harmful ethylene effects in vitro culture since silver ions inhibit this gas's action (Cardoso, 2019; Taiz et al., 2017). Another possible alternative to improve the microenvironments of culture vials is the use of biochars. Biochar are carbon-rich compounds obtained by pyrolysis (thermal decomposition in the absence or partial presence of O2) of organic residues (Amoah-Antwi et al., 2020; Islam et al., 2021). Biochars have porous structures and surface functional groups with electric charges that can adsorb metals and gases (Butnan et al., 2015). Di Lonardo et al. (2013), working with Populus alba L. in vitro, found that the addition of biochar in the culture media reduced ethylene concentrations and favored plant development. The positive effects of activated charcoal and biochar on plant tissue culture are possibly related to the presence of some substances released by these materials that promote plant growth and development and the adsorption of some inhibitory compounds, such as ethylene (Di Lonardo et al., 2013; Thomas, 2008). Although there are few reports in the literature about the effects of biochar in vitro growth, this material is considered an adsorbent of organic and inorganic pollutants due to their properties (high aromaticity, surface charges, and porosity) (Bian et al., 2014; Wang et al., 2014). In general, there are significant increases in the number of thin root plants and plant yield in soils with biochar (Prendergast-Miller et al., 2014; Silva et al., 2017; Xiang et al., 2017). These biochar effects on the plant root system associated with their gas adsorption capacity are desirable for in vitro growth. Thus, the objective of this work is to evaluate the effects of biochar, the cap with or without porous membrane, activated charcoal, and the AgNO3 on the internal micro-environment of vials used for the in vitro growth of pitaya and strawberry.
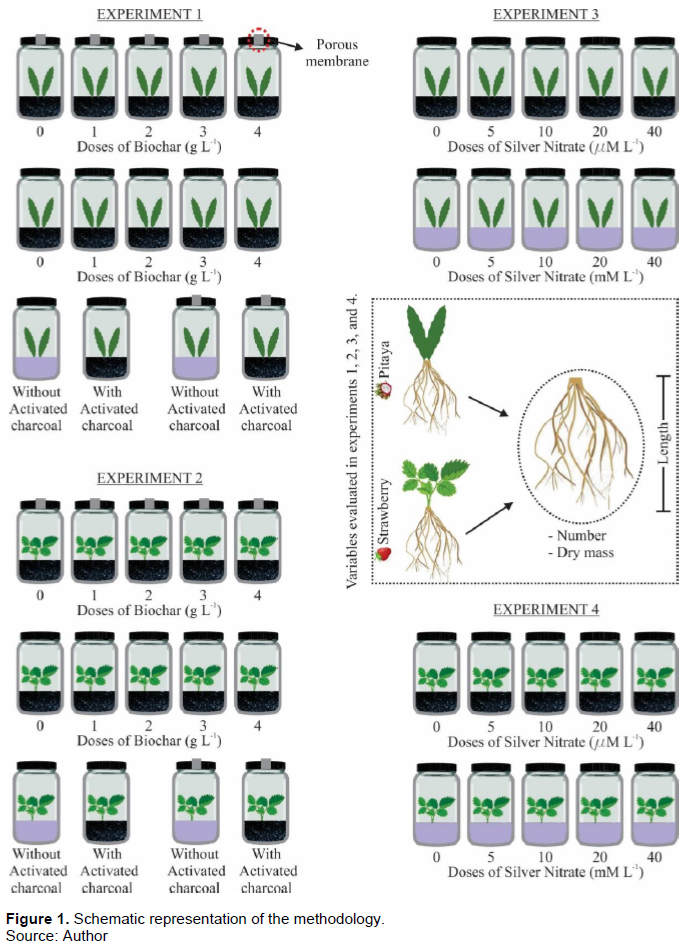
MATERIALS AND METHODS
The experiment was carried out at the Federal University of Minas Gerais, in the city of Montes Claros, MG, Brazil (16°40?57.50” S; 43°50?26.07” W; 650 m). In this study, four experiments were carried out. In the experiment 1 and 2, vials sealed with caps with or without porous membrane that allowed gas exchange (orifice sealed with microporous membrane) were used for in vitro growth of pitaya and strawberry, respectively. Explants of pitaya (1 dm in length) and strawberry (2 cm in length) were previously produced in vitro from seeds of Hylocereus undatus Haw and propagules of Fragaria x ananassa Duch, respectively. The experiments were carried out in a completely randomized design, in a 5x2+2 factorial scheme, with 6 replications and 5 explants per vial. The treatments consisted of 5 doses of biochar (0, 1, 2, 3, 4 g L-1), 2 types of caps (with and without microporous membrane), and 2 additional treatments with 2 g L-1 of activated charcoal in vials with and without caps with microporous membrane (Figure 1). Experiments 3 and 4 were carried out in a completely randomized design, 5x2 factorial schemes, with 6 replications and 5 explants per vial of pitaya and strawberry, respectively. The treatments consisted of five AgNO3 concentrations (0, 5, 10, 20, 40 µM L-1), without and with 4g L-1 of biochar from sugarcane bagasse (Figure 1). Each experimental unit consisted of a 250 cm3 vials containing 40 mL of on solid MS (Murashige and Skoog, 1962) gelled (0.7% agar, w/v) medium supplemented with 30 g L-1 of sucrose, 7 g L-1 of agar, 0.49 µM of Indolbutyric Acid (IBA), at pH 5.5. The media were sterilized by autoclaving at 121°C and 108 kPa for 20 min. The experimental units were maintained in a growth chamber at 25 ± 2ºC, 36 μmol mol m-2 s-1 of irradiance, supplied by 20 W LED lamps, and 16 h photoperiod. On 60 days after the experiments set up, the seedlings were removed from the culture medium and separated into shoots and roots, which were submitted to distilled water asepsis to remove the excess of medium adhered to them. At the end of the experimental period, the length, number, and dry mass of roots were evaluated. For dry mass evaluation, the roots were dried in a forced-air oven at 70°C for 72 h (Figure 1). The biochar was produced from sugarcane bagasse after the mechanical extraction of the juice. The temperature was elevated at a rate of approximately 5°C min-1 until 450°C (temperature was controlled by a thermocouple inserted in the center of the carbonized mass) with a residence time of 30 min. The biochar was ground and passed through a 0.5 mm mesh sieve for the experiment and analysis. Biochar yield was calculated as the pyrolyzed biochar (dry wt.) ratio to the dry mass of the unpyrolyzed feedstock (dry wt.) (biochar yield (%) = (biochar dry mass/feedstock dry mass) × 100) (Table 1).

For the biochars characterization, pH and EC in water (1:10 v/v) were determined following (Rajkovich et al., 2012), and ash was measured according to ASTM D1762-84. Total nutrients and trace elements were determined via ICP-MS/MS (Agilent 8800 triple quadrupole), after microwave digestion (MARS 6 - Microwave Digestion System, CEM Mars Corporation) with concentrated nitric acid using method described in USEPA 3051; and full C and N content by dry combustion method using a LECO CN-2000 elemental analyzer (Leco Corp., St. Joseph, MI, USA). Data were assessed for normality and heterogeneity of variance. In experiments 1 and 2, data were submitted to analysis of variance. When significant, the F (P < 0.05) test compared the type of caps, and each dose of biochar was compared individually with the control treatment (activated charcoal) by Dunnett’s test (P < 0.05). For the biochar doses, regression equations were adjusted. For the variable number of roots, the data were transformed to (x + 1)0.5. In experiments 3 and 4, the data were submitted to analysis of variance, and, when significant, AgNO3 and biochar were compared by the F test (P < 0.05). For AgNO3 doses, regression equations were adjusted. All static analyses were performed using the R Studio software (R Development Core Team, 2017).
RESULTS AND DISCUSSION
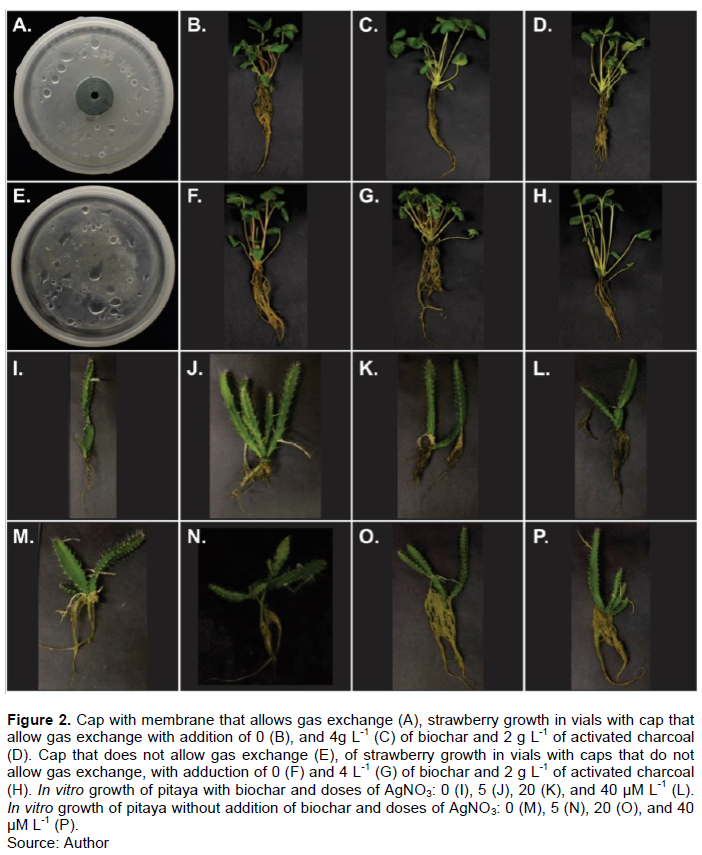
In vitro micropropagation is a technique used to produce seedlings of Pitaya and Strawberry. However, the presence of biochar and AgNO3 in the culture media and porous membrane in caps causes changes in the in vitro culture environment, which promotes changes in the growth and development of the cultivated seedlings. Figure 2 illustrates a cap with a membrane that allows gas exchange (A) and in vitro strawberry growth in vials with caps with membrane and addition of 0 (B) and 4 g L-1 (C) of biochar and 2 g L-1 of activated charcoal (D). Cap without a membrane that allows gas exchange (E) and in vitro strawberry growth in vials with caps without membrane and addition of 0 (F) and 4 L-1 (G) of biochar and 2 g L-1 of activated charcoal (H).. In vitro growth of pitaya with biochar and doses of AgNO3: 0 (I), 5 (J), (K), and 40 µM L-1 (L). In vitro growth of pitaya without addition of biochar and doses of AgNO3: 0 (M), 5 (N), 20 (O), and 40 µM L-1 (P). The type of vials caps, regardless of the addition of biochar or activated charcoal to the culture medium, did no effect on the length, number, and dry mass of roots of pitaya and strawberry, except in doses 0 and 2 g L-1 of biochar, where the dry mass of the strawberry root was more significant when using caps with microporous membrane (Table 2).
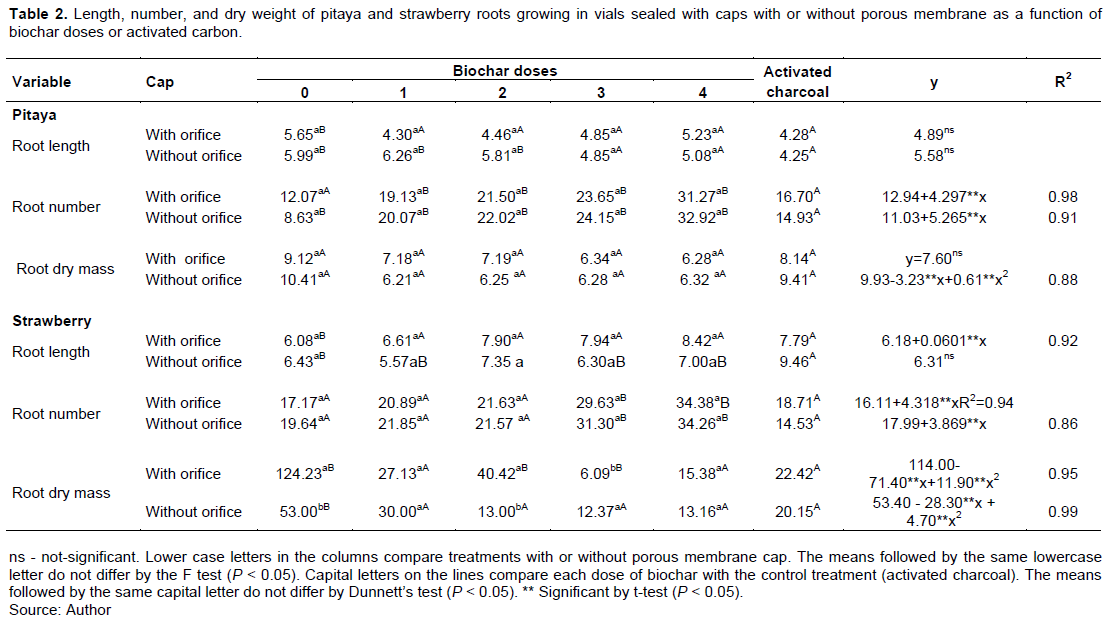
Using microporous membrane caps in vials for in vitro growth allows gas exchange between the inner microenvironmental (inside the vial) and the outer environment (out of the vial). Although we did not obtain significant differences between the types of caps in this study, less formation of water droplets was observed on the vials' inner wall with a porous membrane on the caps, demonstrating the influence of the type of cap the internal microenvironment of the vials. By allowing gas exchange, porous membrane caps had similar effects to activated charcoal in reducing ethylene gas concentrations inside the vials and favored Annona glabra L. root growth in vitro culture (Santana et al., 2011). High concentrations of ethylene inside the vials can have deleterious effects, such as mortality and abscission of plants (Gupta and Pandey, 2019). In vitro culture of Capsicum annuum L. (Mohamed and Alsadon, 2011), Thymus vulgaris L. (Bandeira et al., 2007), and Hancornia speciosa Gomes (Sá et al., 2012), when growing in vials with the caps' porous membrane, showed better growth and development due to the gas exchange. The number and length of roots are essential characteristics of plant adaptability during the acclimatization stage, after in vitro growth. Corroborating the results of this study, other authors also detected significant increases in root length with the addition of biochar in vitro cultivation of lettuce (Lactuca sativa L.) and arabidopsis (Arabidopsis thaliana L.) (Viger et al., 2015).
According to Xiang et al., (2017), roots are the interfaces between biochar particles and plants. Xiang et al (2017) found that, in 32 and 52% of the studies, biochar’s application increased the biomass and length of roots, respectively and concluded that the application of biochar favors root morphological development to increase water and nutrient uptake. According to Viger et al. (2015), biochars alter plant metabolism, such as auxin biosynthesis-related to root system development. The length and dry mass of pitaya and strawberry roots grown with activated charcoal (control treatment, TC) were generally similar to those obtained with the biochar, regardless of the caps types (Table 2), except the dry mass of strawberry roots in the absence of biochar was higher than that obtained with activated charcoal. On the other hand, the number of roots obtained in the activated charcoal treatments was lower than in the higher doses of biochar. Besides, the roots in biochar treatments were thinner than in activated charcoal treatments. Di Lonardo et al., (2013), working in vitro cultivation of two clones of Populus alba L. found that the addition of biochar or activated charcoal contributed positively to the growth of the shoot and number of roots. According to these authors, activated carbon and biochar significantly reduce the ethylene concentration inside the vials by adsorbing this gas in their particles. High ethylene concentrations inside vials for in vitro growth may inhibit organogenesis, cause leaf collapse and abscission, while low ethylene concentrations may contribute to plant growth (Taiz et al., 2017). (Firoozabady et al., 2006) also identified the positive effects of activated charcoal to the culture medium for in vitro growth. These authors observed significant increases in rooting and sprouting of pineapple in cultivate in vitro with activated charcoal. There is little information available on the effects of biochar as a plant hormone adsorbent (Di Lonardo et al., 2013). However, several studies show biochars' ability to adsorb pesticide molecules (Song et al., 2012; Sopeña et al., 2012) and immobilize organic and inorganic pollutants (Bian et al., 2014). On the other hand, some organic compounds can be produced during the pyrolysis process, including ethylene (Spokas et al., 2010). Thus, for the use of biochar in vitro growth, the presence and quantity of toxic compounds must be considered. Besides, depending on the pyrolysis conditions and the raw material, biochar may have high toxic compound concentrations, such as dioxins and furans (Free et al., 2010).
According to some studies, plants can uptake carbon nanoparticles from biochar, which can inhibit the expression of some genes related to secondary metabolism, including ethylene production. The uptake of biochar nanoparticles may also negatively affect the expression of genes responsible for the jasmonic and salicylic acid pathways that participate in ethylene signaling, plant defense against insect attack and pathogens, and stress control (Viger et al., 2015). Therefore, in addition to ethylene adsorption, the biochar effects on the metabolism of plants grown in vitro should be considered. In this sense, many authors have verified the positive impact of biochars on root development and morphology, improving water and nutrient uptake (Prendergast-Miller et al., 2014) and crop yield (Xiang et al., 2017).
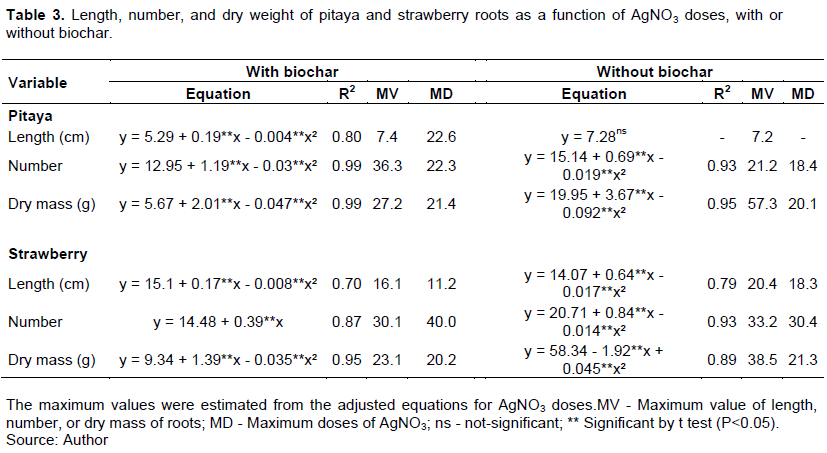
From the equations adjusted for the biochar doses, regardless of the type of the cap, no significant effect of biochar on pitaya root length was identified. In contrast, the strawberry root length increased linearly with biochar doses in treatments with porous membranes in the caps. Comparing the doses 0 and 4g of biochar, the strawberry root length at the highest dose of biochar was 1.4 times longer than at zero. From the equations adjusted for the biochar doses, regardless of the type of the cap, no significant effect of biochar on pitaya root length was identified. In contrast, the strawberry root length increased linearly with biochar doses (Table 2) in treatments with porous membranes in the caps. Comparing the doses 0 (6.18 cm) and 4g L-1 (8.59 cm) of biochar, the strawberry root length at the highest dose of biochar was 1.4 times longer than at zero (Table 2). The number of pitaya and strawberry roots increased linearly with the biochar doses, regardless of the type of caps (Table 2). For the pitaya grown in vials with the porous membrane in the caps, the number of roots increased by 2.32 times from 0 dose (12.94) to the dose 4 g L-1 of biochar (30,11), while in the vials without membrane in the caps, the increase from 0 dose (11.03) to 4 g L-1 of biochar (32, 04) was 2.90 times higher (Table 2). For strawberry (Figure 2D), the increase in the number of roots from 0 dose (16.11) to 4 g L-1 of biochar (33.38) was 2.07 times higher in the treatments with the membrane in the caps, and 1.86 times higher (17.9 roots at 0 dose and 33.46 roots at 4g L-1 of biochar) for caps without membrane (Table 2). For the dry mass of pitaya roots, it was verified that there were no significant differences for biochar doses, regardless of the type of the cap. In contrast, for strawberry, increasing doses of biochar, regardless of caps type, decreased root dry mass production (Table 2). Thus, for the strawberry, it is observed that, although there was an increase in the number of roots, the roots were thin, reflecting in the lower production of dry mass. Silva et al. (2017) also found many thin roots in common bean plants when biochar was added to the soil. Biochar may probably affect plant metabolism, and thinner roots may be an ecological advantage for plant adaptation in low fertility soils (Viger et al., 2015). In the experiments with biochar and AgNO3, the addition of biochar in the culture medium did not influence the length and number of pitaya and strawberry roots. On the other hand, the dry mass yield of pitaya and strawberry roots decreased in biochar treatments. The length, number, and dry mass of pitaya and strawberry roots were adjusted to a quadratic model in the function of AgNO3 doses, except for the number of strawberry roots in treatments with biochar (linear model) and length of pitaya roots the treatments without biochar (there were no differences between AgNO3 doses) (Table 3). According to the results, for both the pitaya and strawberry, there was a reduction in root dry mass production in treatments with biochar (Table 3).The purpose of AgNO3 addition of in vitro culture medium is to inhibit ethylene's action on plants. In this study, biochar may have probably retained ethylene molecules in the surface charges of particles, thereby reducing the concentrations of this gas inside the vials, as discussed earlier. However, the lower root dry mass yield can be attributed to the immobilization of silver ions (Ag+) by biochar. In general, due to ash content, rich in alkali metal oxides and hydroxides (De Almeida et al., 2020), biochars raise the medium's pH, and, consequently, precipitation of cationic metal ions can occur (Rajapaksha et al., 2016; Rizwan et al., 2016). Besides, biochars are very efficient adsorbents of cationic metal ions due to the high surface area, particle porosity, and functional groups that exhibit negative electric charges (Bian et al., 2014; Lu et al., 2014; Tang et al., 2013). The biochar functional groups O-H, C-H, C=O, and C-O, for example, when deprotonated, show negative charges on the particle surface, and can hold Ag+ ions. When making the relationship between the length and the number of roots with the production of dry mass, the results showed that the treatments with biochar provided thinner roots. In treatments without biochar, root dry mass production was 2.1 and 1.7 times higher than in the treatment with biochar, for pitaya and strawberry, respectively (Table 3). The use of microporous membrane caps, activated charcoal, and, or AgNO3 to the culture medium to reduce ethylene concentrations inside vials for in vitro growth has been explored in the literature (Cardoso, 2019; Galdiano Junior et al., 2012; Santana et al., 2011). However, according to our results, the addition of biochar to the culture medium inhibits the effects of AgNO3. Sugarcane bagasse biochar favors the growth and development of pitaya and strawberry roots, regardless of the use of vials with caps with or without porous membranes for gas exchange. There is no difference between vial cap types in the presence of biochar or activated charcoal in the culture medium. Thus, porous membrane caps, sugarcane bagasse biochar, and AgNO3 are viable alternatives to improve the conditions for in vitro growth of pitaya and strawberry. However, it is not recommended to add AgNO3 and biochar together to the same culture medium. Using biochar for in vitro micropropagation of plants proved to be a promising alternative in different conditions (presence and absence of activated charcoal or porous membranes in cap). However, there are still issues to address in future research as: to whether the type of raw material, temperature, and pyrolysis time used to generate the biochar influence the growth and development of plants in vitro.
CONCLUSION
The use of porous membrane caps contributed only to increase the roots dry matter yield of strawberries grown in vitro with culture media with 0, 1, and 2 g L-1 of biochar. The use of porous membrane caps did not influence the growth and development of pitaya roots grown in vitro. The addition of 4 g L-1 of biochar in the culture media favored the growth and development of the pitaya and strawberry roots, regardless of the use or not of porous membrane caps in vials of the in vitro culture. The AgNO3 alone contributed to the greater length and dry mass of pitaya roots and higher dry mass of strawberry roots. The addition of biochar in the culture medium decreases the AgNO3 effects in vitro growth.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
The authors are grateful to Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG), and Pró-Reitoria de Pesquisa da Universidade Federal de Minas Gerais (PRPq -UFMG) for their financial support
REFERENCES
|
Amoah-Antwi C, Kwiatkowska-Malina J, Thornton SF, Fenton O, Malina G, Szara E (2020). Restoration of soil quality using biochar and brown coal waste: A review. Science of The Total Environment 722:137852. |
|
|
Bandeira JDM, Lima CSM, Rubin S, Ribeiro MV, Falqueto AR, Peters JA, Braga EJB (2007). Diferentes tipos de vedações dos frascos e concentrações de sacarose na micropropagação de Thymus vulgaris L. Revista Brasileira de Biociências 5(s2):472-474. |
|
|
Bian R, Joseph S, Cui L, Pan G, Li L, Liu X, Zhang A, Rutlidge H, Wong S, Chia C, Marjo C, Gong B, Munroe P, Donne S (2014). A three-year experiment confirms continuous immobilization of cadmium and lead in contaminated paddy field with biochar amendment. Journal of Hazardous Materials 272:121-128. |
|
|
Butnan S, Deenik JL, Toomsan B, Antal MJ, Vityakon P (2015). Biochar characteristics and application rates affecting corn growth and properties of soils contrasting in texture and mineralogy. Geoderma 237-238:105-116. |
|
|
Cardoso JC (2019). Silver nitrate enhances in vitro development and quality of shoots of Anthurium andraeanum. Scientia Horticulturae 253:358-363. |
|
|
De Almeida JB, Santos LS, Pandey SD, Nunes CF, Colen F, Sampaio RA, Frazão LA, Pegoraro RF, Fernandes LA (2020). Alleviation of aluminum and cadmium toxicity by biochar and its potential toxicity to Sorghum. Semina: Ciências Agrárias 41(1):95-108. |
|
|
Di Lonardo S, Vaccari FP, Baronti S, Capuana M, Bacci L, Sabatini F, Lambardi M, Miglietta F (2013). Biochar successfully replaces activated charcoal for in vitro culture of two white poplar clones reducing ethylene concentration. Plant Growth Regulation 69:43-50. |
|
|
Firoozabady E, Heckert M, Gutterson N (2006). Transformation and regeneration of pineapple. Plant Cell, Tissue and Organ Culture 84:1-16. |
|
|
Free HF, McGill CR, Rowarth JS, Hedley MJ (2010). The effect of biochars on maize (Zea mays) germination. New Zealand Journal of Agricultural Research 53:1-4. |
|
|
Galdiano Junior RF, Mantovani C, Pivetta KFL, Lemos EGM (2012). In vitro growth and acclimatization of Cattleya loddigesii Lindley (Orchidaceae) with actived charcoal in two light spectra. Ciência Rural 42(5):801-807. |
|
|
George EF, Hall MA, Klerk GJ (2008). Plant Propagation by Tissue Culture. Netherlands: Springer. |
|
|
Gupta S, Pandey S (2019). Unravelling the biochemistry and genetics of ACC deaminase-An enzyme alleviating the biotic and abiotic stress in plants. Plant Gene 18:100175. |
|
|
Hoang NN, Kitaya Y, Shibuya T, Endo R (2020). Effects of supporting materials in in vitro acclimatization stage on ex vitro growth of wasabi plants. Scientia Horticulturae 261:109042. |
|
|
Iarema L, da Cruz ACF, Saldanha CW, Dias LLC, Vieira RF, de Oliveira EJ, Otoni WC (2012). Photoautotrophic propagation of Brazilian ginseng [Pfaffia glomerata (Spreng.) Pedersen]. Plant Cell, Tissue and Organ Culture 110:227-238. |
|
|
Islam MS, Chen Y, Weng L, Ma J, Khan ZH, Liao Z, Magid ASIA, Li Y (2021). Watering techniques and zero-valent iron biochar pH effects on As and Cd concentrations in rice rhizosphere soils, tissues and yield. Journal of Environmental Sciences 100:144-157. |
|
|
Kozai T (2010). Photoautotrophic micropropagation - environmental control for promoting photosynthesis. Propagation of Ornamental Plants 10(4):188-204. |
|
|
Lu K, Yang X, Shen J, Robinson B, Huang H, Liu D, Bolan N, Pei J, Wang H (2014). Effect of bamboo and rice straw biochars on the bioavailability of Cd, Cu, Pb and Zn to Sedum plumbizincicola. Agriculture, Ecosystems & Environment 191:124-132. |
|
|
Mohamed MAH, Alsadon AA (2011). Effect of vessel type and growth regulators on micropropagation of Capsicum annuum. Biologia plantarum 55:370-374. |
|
|
Mohamed MAH, Alsadon AA (2010). Influence of ventilation and sucrose on growth and leaf anatomy of micropropagated potato plantlets. Scientia Horticulturae 123:295-300. |
|
|
Murashige T, Skoog F (1962). A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiologia Plantarum 15(3):473-497. |
|
|
Naing AH, Jeon SM, Han JS, Lim SH, Lim KB, Kim CK (2014). Factors influencing in vitro shoot regeneration from leaf segments of Chrysanthemum. Comptes Rendus Biologies 337(6):383-390. |
|
|
Prendergast-Miller MT, Duvall M, Sohi SP (2014). Biochar-root interactions are mediated by biochar nutrient content and impacts on soil nutrient availability. European Journal of Soil Science 65(1):173-185. |
|
|
R Development Core Team (2017). R: A language and environment for statistical computing, reference index version 3.4.2. |
|
|
Rajapaksha AU, Chen SS, Tsang DCW, Zhang M, Vithanage M, Mandal S, Gao B, Bolan NS, Ok YS (2016). Engineered/designer biochar for contaminant removal/immobilization from soil and water: Potential and implication of biochar modification. Chemosphere, 148:276-291. |
|
|
Rajkovich S, Enders A, Hanley K, Hyland C, Zimmerman AR, Lehmann J (2012). Corn growth and nitrogen nutrition after additions of biochars with varying properties to a temperate soil. Biology and Fertility of Soils 48:271-284. |
|
|
Rizwan M, Ali S, Qayyum MF, Ibrahim M, Zia-ur-Rehman M, Abbas T, Ok YS (2016). Mechanisms of biochar-mediated alleviation of toxicity of trace elements in plants: a critical review. Environmental Science and Pollution Research 23:2230-2248. |
|
|
Rodrigues FA, Rezende RALS, Pasqual M, Lopes MTG (2017). Solidifying agents and activated charcoal for in vitro culture of Solanum sessiliflorum. Pesquisa Agropecuária Brasileira 52(11):1123-1126. |
|
|
Sá ADJ, Lédo AS, Lédo CAS, Pasqual M, Silva AVC, da Silva Junior JF (2012). Tipo de vedação e explantes na micropropagação de mangabeira. Ciência e Agrotecnologia 36(4):406-414. |
|
|
Santana JRF, Paiva R, Souza AV, Oliveira LM (2011). Effect of different culture tube caps and concentrations of activated charcoal and sucrose on in vitro growth and budding induction of Annona glabra L. Ciência e Agrotecnologia 35(5):916-923. |
|
|
Silva ICB, Basílio JJN, Fernandes LA, Colen F, Sampaio RA, Frazão LA (2017). Biochar from different residues on soil properties and common bean production. Soils and Plant Nutrition 74(5):378-382. |
|
|
Song Y, Wang F, Bian Y, Kengara FO, Jia M, Xie Z, Jiang X (2012). Bioavailability assessment of hexachlorobenzene in soil as affected by wheat straw biochar. Journal of Hazardous Materials, 217-218:391-397. |
|
|
Sopeña F, Semple K, Sohi S, Bending G (2012). Assessing the chemical and biological accessibility of the herbicide isoproturon in soil amended with biochar. Chemosphere 88:77-83. |
|
|
Spokas KA, Baker JM, Reicosky DC (2010). Ethylene: Potential key for biochar amendment impacts. Plant and Soil 333:443-452. |
|
|
Taiz L, Zeiger E, Moller IM, Murphy A (2017). Fisiologia e Desenvolvimento Vegetal. Porto Alegre, BR: Artmed. |
|
|
Tang J, Zhu W, Kookana R, Katayama A (2013). Characteristics of biochar and its application in remediation of contaminated soil. Journal of Bioscience and Bioengineering 116(6):653-659. |
|
|
Thomas TD (2008). The role of activated charcoal in plant tissue culture. Biotechnology Advances 26(6):618-631. |
|
|
Vieira CF, Carvalho FEL, Lima-Melo Y, Carvalho CPS, Lima Neto MC, Martins MO, Silveira JAG (2019). Integrative approach reveals new insights into photosynthetic and redox protection in ex vitro tobacco plantlets acclimatization to increasing light intensity. Biotechnology Research and Innovation 3:59-72. |
|
|
Viger M, Hancock RD, Migletta F, Taylor G (2015). More plant growth but less plant defence? First global gene expression data for plants grown in soil amended with biochar. GCB Bioenergy 7(4):658-672. |
|
|
Waikhom SD, Loui B (2014). An effective protocol for micropropagation of edible bamboo species (Bambusa tulda and Melocanna baccifera) through nodal culture. The Scientific World Journal 2014:345794. |
|
|
Wang F, Ji R, Jiang Z, Chen W (2014). Species-dependent effects of biochar amendment on bioaccumulation of atrazine in earthworms. Environmental Pollution 186:241-247. |
|
|
Xiang Y, Deng Q, Duan H, Guo Y (2017). Effects of biochar application on root traits: a meta-analysis. GCB Bioenergy 9(10):1563-1572. |
|
|
Xiao Y, Niu G, Kozai T (2011). Development and application of photoautotrophic micropropagation plant system. Plant Cell, Tissue and Organ Culture 105:149-158. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


