Full Length Research Paper

ABSTRACT
The root parasitic weed Striga hermonthica constitutes a threat to cereals production in sub-Saharan Africa. Cowpea has been used as a rotational or a companion crop to combat the parasite on cereals with varying success. The present investigation was set to study the effects of root powder from 12 cowpea genotypes and root exudates from two genotypes on germination and radicle length in S. hermonthica, sorghum and millet strains. At 5 to 15 mg root powders from T198K-409-4 and T198K-317-2, the least and most active on S. hermonthica sorghum strain, induced 41-60 and 79-86% germination, respectively. Root powders from B301 and T100K-1263, the least and most active on the pearl millet strain induced 33-46 and 94-95%, germination, respectively. Maximum germination in response to root exudates of B301 and T100K-901-6, each applied at 5, 10 and 15 µl, was 54, 45 and 36% and 52, 43 and 31%, respectively for the sorghum strain and 43, 53 and 61% and 44, 49 and 53%, respectively for its pearl millet congener. B301and T100K-901-6 root exudates induced maximum germination at pH 10 and 7, respectively. Root powder reduced radicle length of both strains, while root exudates reduced radicle length of the sorghum strain, but increased that of its pearl millet congener. Employment of cowpea as a rotational or a companion crop to combat S. hermonthica on cereals implies rigorous selection through initial laboratory screening for stimulant production.
Key words: Striga hermonthica strains, root powder, root exudates, germination, sorghum, pearl millet.
INTRODUCTION
Sorghum [Sorghum bicolor (L.) Moench] and pearl millet [Pennisetum glaucum (L) R. BR.], are elemental crops for food security and income generation in sub-Saharan Africa, where the prevailing drought limits crop choice to only a few (Babiker, 2007). Population pressure and market demands have led to advancement of agriculture into marginal lands and intensification of sorghum and pearl millet planting, often in monocultures (Parker and Riches, 1993). Nevertheless, yields are by far below the international average (Babiker, 2007). The low yields are attributable to a multitude of factors among which, poor soil fertility, low inputs and heavy infestations by the root parasitic weed Striga hermonthica (Del.) Benth. are of paramount importance (Babiker, 2007). Available reports showed that Striga spp. are prevalent in over 50 million hectares of the cereals growing areas in Africa and inflict considerable damage amounting to complete crop loss under heavy infestations (Welsh and Mohamed, 2011; Spallek et al., 2013).
S. hermonthica, well adapted to its environment and tolerant to a wide range of temperature and soil moisture stress, has developed two distinct strains. The first, specific to pearl millet, is predominant in the drier northern regions of sub-Saharan Africa, while the second attacks sorghum and is found farther south in the wetter regions (Welsh and Mohamed, 2011). The parasite produces thousands of minute dust-like seeds with prolonged viability and special germination requirements. To germinate a Striga seed requires being in close vicinity of a host root, a pre-treatment (conditioning) in a warm moist environment for several days and a subsequent exposure to an exogenous stimulant. Radicles of the resulting seedlings elongate and haustoria are initiated. Haustori attach penetrate the host roots, and establish connection with the vascular system. Subsequent to establishment of connection with the host xylem, the parasite develops and remains subterranean for 6 to 8 weeks prior to emergence.
The unavoidable low crop productivity arising from the damage the parasite inflicts while subterranean makes farmers reluctant to adopt post-emergence control measures (Babiker, 2007). Post-emergence control measures are essential to curtail replenishment of the parasite seed reserves. The need for simple affordable, environmentally benign management practices which enhance seed bank demise is thus, imperative. The ubiquitous nature of strigolactones (SLs), the natural Striga germination stimulants, within the plant kingdom, confers some flexibility on designing agroecologically and socioeconomically acceptable cropping systems.
Traditional African farming, based on prolonged fallows and intercropping with legumes, particularly cowpea (Vigna unguiculata L., Walp), allowed for rejuvenation of soil fertility and demise of the parasite seed bank (Ejeta et al., 1993; Kureh et al., 2006). However, S. hermonthica sorghum and millet strains differ in their response to natural germination stimulants (Parker and Riches, 1993). The sorghum strain is more responsive to the strigol-type SLs, but less so to the orobancol-type, whereas the millet strain is less responsive to the strigol-type (Parker and Reid, 1979; Cardoso et al., 2014). Consequently performance of cowpea, reported to produce mainly orobancol-type SLs (Ueno et al., 2011), as an efficient rotational or companion crop to combat S. hermonthica on sorghum is contestable and merits further research.
The present investigation was therefore set to study the effects of root powder from 12 cowpea genotypes and root exudates from two genotypes on germination and radicle length in S. hermonthica, sorghum and millet strains.
MATERIALS AND METHODS
S. hermonthica, sorghum and pearl millet strains, seeds were collected in 2012 from under sorghum in the Gezira, Central Sudan, and pearl millet in Kordofan State Western Sudan, respectively. The seeds were surface disinfected by immersion in 1% sodium hypochlorite, obtained by dilution of the respective amount of commercial bleach solution, for 1 min washed with sterilized distilled water and air-dried. The seeds were subsequently conditioned as previously described by Babiker et al. (2000). Briefly the seeds (40-60) were sprinkled on glass fiber filter paper discs (8 mm) placed in Petri-dishes lined with glass fiber filter papers and sufficiently moistened with sterilized distilled water. The Petr-dishes sealed with parafilm, were incubated at 30ºC in the dark for 10 to 14 days prior to use for germination assays. Twelve cowpea genotypes were obtained from the International Institute of Tropical Agriculture (IITA) Ibadan Nigeria. GR24, a synthetic Striga germination stimulant, was a gift from Professor B. Zwanenburg Radbound University, Nijmegen the Netherlands. Cowpea seeds were surface disinfected by immersion in 1% sodium hypochlorite (NaOCl) for 5 min. The seeds were subsequently washed with sterilized distilled water, air dried in a laminar flow cabinet and stored at ambient temperature, till used. Experiments were repeated at least twice. Data presented were from typical experiments. In all experiments treatments were arranged in a complete randomized design with 5 replicates unless mentioned otherwise an aqueous and a GR24 (0.1 mg L-1) treated controls were included in each experiment for comparison.
Effects of root powder on germination and radicle length of S. hermonthica
A total of 12 cowpea genotypes, grown in pots in a screen house for 10 days, were harvested and the roots were, carefully, washed to remove soil particles. The roots, dried under ambient conditions, were ground into fine powder using a kitchen grinder and subsequently stored. The dried roots powder (5 to 15 mg equivalent to field rates of 25.5 to 76.4 kg ha-1) was assayed for germination inducing activity and effects on radicle length of the two Striga strains using the sandwich method as described by Fujii et al. (2004). Briefly, agar, nutrient-less (3 g) was added to 1000 ml of distilled water and autoclaved for 20 min at 15 bars and 121°C. The agar, cooled in a water bath set at 40°C, was pipetted into multiwell plates (5 ml/well) and allowed to solidify prior to careful spreading of the test powder sample, by hand, on top. Another 5 ml of agar were placed on top, allowed to solidify, and subsequently overlaid by discs containing conditioned S. hermonthica, sorghum or pearl millet strain. The multiwell plates, sealed with parafilm and wrapped with aluminum foil, were incubated at 30°C in the dark for 48 h prior to examination for germination and radicle length, using a stereomicroscope equipped with a calibrated ocular micrometer. Controls without test samples were included for comparison. Germination, expressed as percentage, was taken on a scale where 0-49.9, 50-59.9, 60-69.9, 70.0-79.9 and 80.0-100% indicated poor, moderate, satisfactory, good and excellent germination, respectively. Radicles measuring 0.1-4.9×10-2, 5-9.9×10-2, 10-14.9×10-2 and ≥15×10-2 mm were considered very short, short, medium and long, respectively.
Effects of incubation time on production of germination and radicle length effectors
The genotypes B301 and T100K-901-6, expected, based on their prostrate and spreading growth habit to be less competitive under field conditions, were selected. The seeds of the two genotypes were surface sterilized and pre-germinated, at 30°C in the dark, in sterilized filter papers rolls. The seedlings, grown hydroponically, each in a 10 ml glass tube containing 40% Long Ashton solution, were maintained in a growth chamber at 30°C with a 12-h photoperiod using fluorescent lights (photosynthetic active radiation of 220 µmol m-2 s-1) for 4 days and subsequently transferred to tap water in glass tubes (100 ml capacity). The glass tubes, wrapped in aluminum foil, to exclude light, were re-incubated for 4 days prior to sampling made daily over a period of 10 days. Prior to sampling the water volume, in each tube, was allowed to drop to 50 ml or adjusted to 50 ml. The samples, 2 ml each, were subsequently tested for germination inducing activity and effects on radicle length. Aliquots (5, 10 and 20 µl) were applied, each, to an 8 mm glass fiber disc containing conditioned Striga seeds, placed in Petri dishes. The seeds were re-incubated and germination and radicle length were determined as described previously.
Effects of pH on germination inducing activity of root exudates:
Root exudates from cowpea genotypes B301 and Tk100-901-6 (30 ml each) grown hydroponically for 10 days, as above, were extracted with ethylacetate (15 ml×3) as described by Yoneyama et al. (2010). The ethylacetate extracts, allowed to stand overnight at 4°C on anhydrous sodium sulphate, were evaporated to dryness at 40°C using a rotary evaporator. The residues were re-dissolved each in 2 ml of acetone. Aliquots of the acetone solution (100 µl) were added each to1.9 ml of 0.1 M Na2HPO4/NaH2PO4 buffers for pH 7 or 0.1M NaCl/HCl buffer for pH 4 and pH 10. Aliquots (5, 10 and 15 µl) from each of the respective buffered solutions were assayed for germination inducing activity, as above, using conditioned Striga sorghum and pearl millet strains. Acetone at the concentration used has no adverse effects on S. hermonthica seed germination.
Statistical analyses
Data collected from all experiments were subjected to statistical analyses using Statistix 8 statistical software, version 2.0 (UK). Means were separated for significance using the Least Significant Difference (LSD at P≥0.05). Figures were drawn, when appropriate, using Sigma plot version 11 and Microsoft Office Excel 2013.
RESULTS
Effects of root powder on germination of S. hermonthica
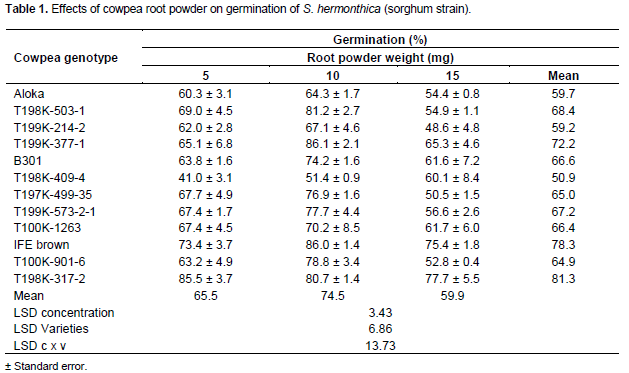
For Striga sorghum strain, GR24 at 0.1 mg L-1 induced 77-90% germination. Cowpea root powder at 5 mg, poor (41%), satisfactory (60.3- 69.0%), good (73.4%) and excellent (85.5%) germination was effected by 1, 9, 1 and 1 samples, respectively (Table 1). At 10 mg moderate, satisfactory, good and excellent germination was elicited by 1, 2, 5 and 4 samples, respectively. At 15 mg poor (48.6%), moderate (50.5-56.6%), satisfactory (60.1-65.3%) and good (75.4 and 77.7%) germination was elicited by 1, 5, 4 and 2 samples, respectively. Of all cowpea root powder from the genotypes T198 K-409-4, T199 K-214-2 and Akola exhibted the lowest (50.9-59.7%) mean germination, while those from T199 K-377-1, IFE brown and T198 K-317-2 displayed the highest (72.2-81.3%) (Table 1).
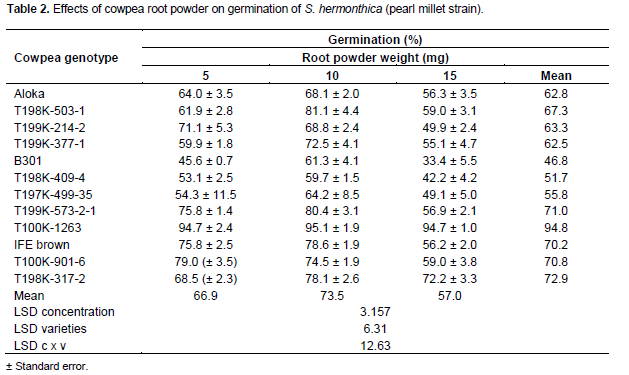
For the Striga pearl millet strain, GR24 at 0.1 mgL-1 induced 55-68% germination. Cowpea root powder at 5 mg, poor (45.6%), moderate (53.1-59.3%), satisfactory (61.9-68.5%), good (71.1-79%) and excellent (94.7%) germination was achieved by 1, 3, 3, 4 and 1 sample, respectively (Table 2). At 10 mg 1 sample induced moderate (59.7%) and 4, each, resulted in satisfactory (64.2-68.4%) and good (72.5-78.6%) germination, while 3 samples achieved excellent (80.4-95 %.) germination. At 15 mg poor (33.4-49.9%), moderate (55.1-59%), good (72.2%) and excellent (94.7%) germination was induced by 4, 6, 1 and 1 samples, respectively (Table 2). Of all cowpea genotypes root powder from B301, T198 K-409-4 and T197-499-35 effected the lowest (46.8-55.8%) mean germination, whereas those from T198 K-3172, T100 K-901-6 and T100K-1263 induced the highest (72.9-94.8%) (Table 2).
Effects of incubation time on production of germination stimulant(s)
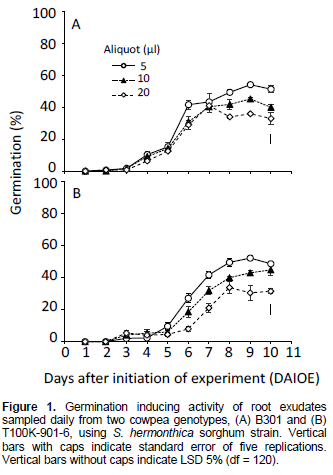
Cowpea root exudates sampled 1 to 3 days after initiation of experiment (DAIOE), irrespective of volume, genotype or Striga strain, induced negligible to little germination (Figure 1). For Striga sorghum strain, GR24 at 0.1 mg L-1 induced 80 to 89% germination. Seeds treated with distilled water displayed no germination. Root exudates from genotype B301 at 5 and 10 µl attained maximum germination, 54 and 45.1%, respectively, at 9 DAIOE (Figure 1A). At 20 µl, exudates volume, germination was maximum (41%) at 7 DAIOE, but declined to 32.9 to 35.8% on further extension of sampling time to 8 to 10 DAIOE. Root exudates from genotype T100K-901-6 at 5 and 10 µl achieved maximum germination 52.1 and 43%, respectively 9 and 10 DAIOE (Figure 1B). At 20 µl maximum germination (33.8%) was realized 8 DAIOE with no further significant change on extension of the sampling period to 10 DAIOE.
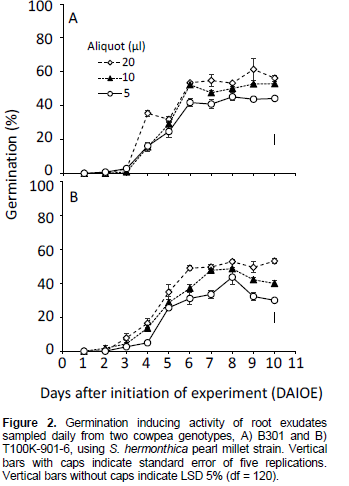
For Striga pearl millet strain GR24 at 0.1 mg L-1 induced 60 to 65% germination. The aqueous control displayed no germination. Root exudates from cowpea B301, sampled 4 DAIOE, at 5 and 10 µl showed low germination. However, at 20 µl a substantial germination (35.4%) was displayed (Figure 2A). Root exudates sampled 6 DAIOE and applied at 5, 10 and 20 µl induced 41.8, 52.3 and 53.4% germination, respectively. A further increase in sampling time had no significant effects on germination inducing activity at exudates volumes of 5 and 10 µl, however at 20 µl a significant increase was displayed and germination was maximal (61.4%) 9 DAIOE (Figure 2A).
Root exudates from cowpea T100K-901-6 sampled 4 DAIOE showed negligible germination at 5 µl (Figure 2 B). Increasing exudates volume to 10 and 20 µl increased germination to 13.6 and 16.4%, respectively. Exudates sampled 6 DAIOE induced 25.6, 28.7 and 34.8% germination at 5, 10 and 20 µl volume, respectively. Germination inducing activity consistently increased with sampling time and was maximal at 8 DAIOE, where germination was 43.6, 48.9 and 52.8% for the lowest, middle and highest exudates volume, respectively (Figure 2B).
Effects of pH on germination inducing activity of cowpea root exudates
For Striga sorghum strain root exudates from cowpea genotype B301 at 5 µl induced 17.1 to 25.4% germination with no significant differences between pH levels (Table 3). Root exudate at 10 and 15 µl induced 16.7-21.4 and 12-14.2% germination, respectively and the attained germination was significantly higher at pH10. Across exudates volumes mean germination, invariably, decreased with volume and was 22.4, 18.7 and 13.2% at 5, 10 and 15 µl root exudates volume, respectively.
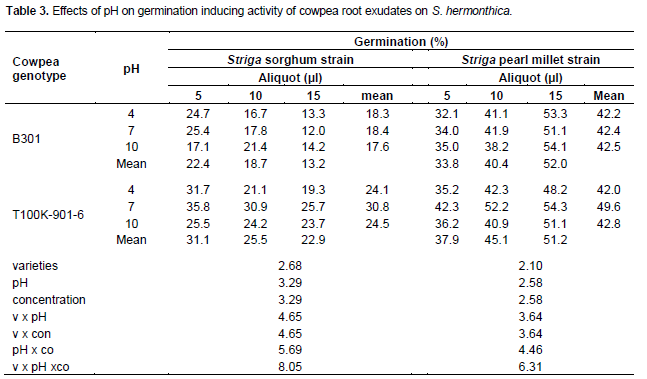
Root exudates of the genotype T100K-901-6, at 5 µl, elicited highest (35.8%) and lowest (25.5%) germination at pH 7 and 10, respectively (Table 3). Increasing exudates volume to 10 µl decreased germination at all pH levels with highest (30.9%) and lowest (21.1%) germination at pH 7 and 10, respectively. At 15 µl maximal (25.7%) and minimal (19.3%) germination were achieved at pH 7 and 4, respectively. Across exudates volume mean germination was highest (31.1%) and lowest (22.9%) at 5 and 15 µl volumes, respectively.
For Striga pearl millet strain root exudates from cowpea B301, at 5 µl induced the lowest germination (32.1%) at pH 4. Germination showed a slight non-significant increase at pH 7, but was significantly higher (35%) at pH 10 (Table 3). Increasing exudates volume to 10 and 15 µl increased germination, however, differences between pH levels were not significant. Across exudates volume mean germination was 33.8, 40.4 and 52% at 5, 10 and 15 µl , respectively (Table 3).
Cowpea genotype T100K-901-6 root exudates at 5 µl induced low germination (35%) at pH 4 (Table 3). Increasing pH to7 increased germination significantly however, a further increase in pH to10 resulted in a non-significant drop. At 10 µl the root exudates showed 42 and 41% germination at pH 4 and pH10, respectively. However, at pH 7 a significantly higher germination (52%) was attained (Table 3). At 15 µl root exudates germination was maximal (54.3%) at pH 7 and minimal (48.2%) at pH 4. Across pH levels mean germination was, significantly, the highest (59.6%) at pH 7. Across exudates volumes mean germination was highest (51.2%) and lowest (37.9%) at the highest (15 µl) and lowest (5 µl) volumes, respectively.
Radicle length
Root powder
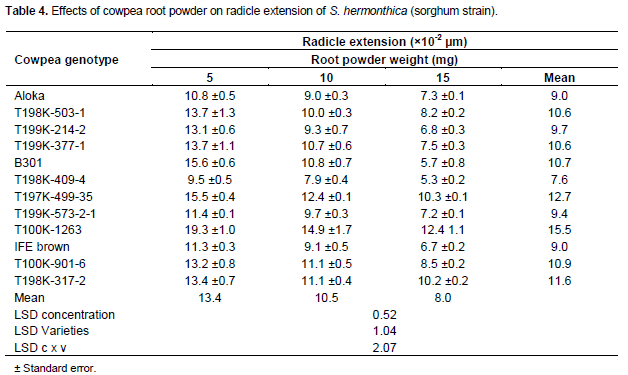
S. hermonthica seedlings, sorghum strain, induced by GR24 at 0.1 mg L-1 displayed long radicles length (17.3-25.7×10-2 mm). Cowpea root powder, invariably resulted in seedlings with shorter radicles than GR24. At 5 mg root powder seedlings with short, medium and long radicles length were effected by 1, 8 and 3 samples, respectively (Table 4). At 10 mg 5 and 7 samples resulted in seedlings with short and medium radicle length, respectively. At 15 mg seedlings with short and medium radicle length were induced by 9 and 3 samples, respectively. Across powder levels 41% of the genotypes induced seedlings with short radicles length (7.6-9.7×10-2mm), 50% educed seedlings with medium radicles length (10.6-11.6×10-2mm) and 8.3% resulted in seedlings with long radicles length (15.5×10-2 mm). Striga seedlings educed by cowpea genotypes T198K-409-4, Aloka and IFE brown displayed the shortest radicle length (7.6-9×10-2 mm)whereas those elicited byT198K-317-2, T197K-499-35 and T100K-1263 exhibited the longest (11.6-15.5×10-2 mm) radicles length.
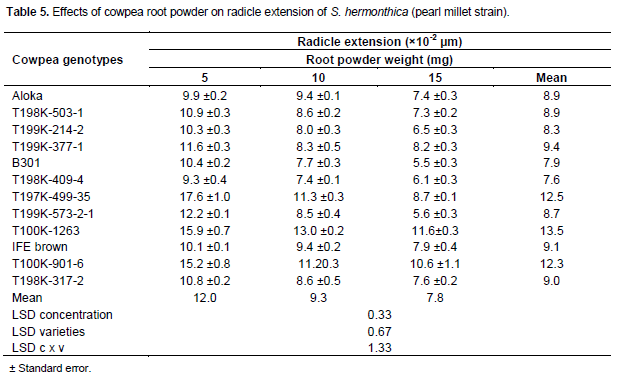
For S. hermonthica, pearl millet strain, seedlings induced to germinate with GR24 at 0.1 mg L-1 displayed short to long (9.4-19 ×10-2mm) radicles length. At 5 mg cowpea root powder seedlings with short, medium and long radicles length were induced by 2, 7 and 3 samples, respectively (Table 5). At 10 mg seedlings with short and medium radicles were length induced by 9 and 3 samples, respectively. At 15 mg seedling with short and medium radicles were induced length by 10 and 2 samples, respectively. Across powder samples 75% of the genotypes effected seedlings with short radicles (7.6- 9.4×10-2 mm) length and 25% resulted in seedlings with medium size (12.3-13.5×10-2 µm) radicles length. Striga seedlings induced by the genotypes T198K-409-4, B301 and T199K-241-2 displayed the shortest (7.6-8.3 × 10-2mm) radicle length, while those induced by the genotypes T100K-901-6, T197K-499-35 and T100K1263 exhibited the longest (12.3-13.5 ×10-2 mm) radicles length (Table 5).
Root exudates
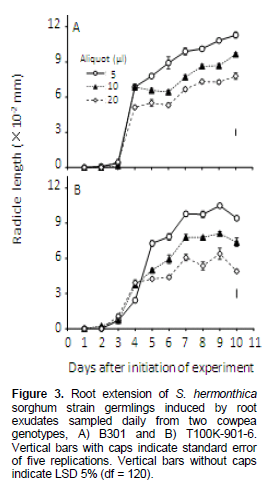
Striga seedlings from seeds induced to germinate by cowpea root exudates sampled 1-3 DAIOE, irrespective of genotype, volume or Striga strain, displayed negligible to short radicle length (Figures 3 and 4 ). For Striga sorghum strain, seedlings induced by GR24 displayed radicle length of 28-31.2 × 10-2mm. Cowpea root exudates sampled at 4 DAIOE or more radicle length progressively, increased with time and decreased with volume of root exudates (Figure 3). Exudates from genotype B301collected 4-7 DAIOE at 510 and 20 µl induced seedlings with short radicle length (5.1-9.9 × 10-2 mm). On further length of sampling time to 8 to 10 DAIOE the root exudates at 5 µl resulted in seedlings with medium (10.1-11.3 × 10-2 mm) radicle length. However, at 10 and 20 µl only seedlings with short (7.3-9.7 × 10-2 mm) radicle length were displayed. For root exudates from cowpea genotype T100K-901-6 sampled length 6 DAIOE or more radicle length was maximal at 9 DAIOE. Radicle length affected by root exudates sampled 9 DAIOE at 5, 10 and 20 µl was 10.5 × 10-2, 8.1 × 10-2 and 6.4 × 10-2 mm, respectively.
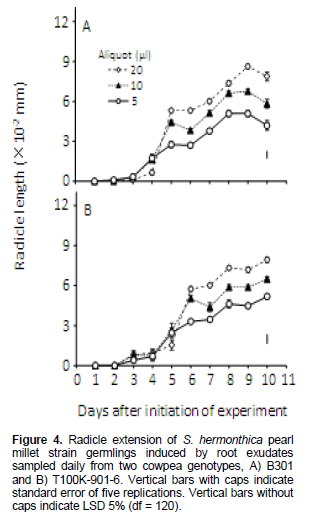
For Striga pearl millet strain, seedlings induced by GR24 displayed radicle length of 16.1-18.8 × 10-2 mm. Radicle length of seedlings induced by cowpea genotype B301 root exudates sampled length6 DAIOE or more progressively increased with time and exudates volume, reached a maximum at 9 DAIOE where it was 5.1× 10-2, 6.8× 10-2 and 8.6 × 10-2 mm for exudates volumes of 5, 10 and 20 µl, respectively (Figure 4A).
Radicle length in seedlings from seeds induced to germinate by roots exudates from cowpea genotype TK100-901-6 followed the same trends as those of seedlings educed by root exudates of genotype B301 (Figure 4). Radicle length was maximal at 10 DAIOE where it was 5.2 × 10-2, 6.5 × 10-2 and 7.9× 10-2 mm at 5 , 10 and 20 µl exudates volume, respectively.
DISCUSSION
The general features of the germination inducing activity of cowpea root powder, irrespective of genotype or Striga strain, were an increase and a subsequent decrease with sample weight (Tables 1 and 2). However, the magnitude of the germination varied with cowpea genotype and Striga strain. Variations in germination inducing activity with genotype, amount of root powder and Striga strains, in line with several reports could be attributed to variations in qualitative and quantitative composition of the active substances (Sato et al., 2005; Yoneyama et al., 2010), simultaneous production of germination stimulants and inhibitors (Muller et al., 1992) and to inherent differences in response of the two S. hermonthica strains to germination stimulants (Parker and Riches, 1993). The high frequency of occurrence of high germination inducers of the pearl S. hermonthica millet strain among cowpea genotypes (41.6%) compared to the low frequency (24.9%) for its sorghum congener (Tables 1 and 2) indicates better chances of success on random selection of genotypes as rotational or companion crops for combating the parasite on pearl millet. However, for sorghum, selection based on prior knowledge is critical.
The low initial germination inducing activity of root exudates observed 1 to 3 DAIOE, irrespective of cowpea genotype or Striga strain may be attributed to a lag phase in stimulant production (Figure 1). A similar initial lag phase in stimulant production was reported for hydro-ponically grown cotton (Sato et al., 2005) and was attributed to an acclimatization process. In general the highest germination inducing activity was attained by root exudates sampled 8 DAIOE and was maintained without a significant change throughout the experiment (Figure 1).
The contrasting response of S. hermonthica pearl millet and sorghum strains to germination stimulants from cowpea root exudates (Figures 1 and 2 and Table 3) is in line with their differential response to root exudates of their respective true hosts (Parker and Riches, 1993). It is noteworthy that sorghum produces, mainly, strigol-type SLs, while cowpea produces, mainly, orobancol-type (Awad et al., 2006; Uneo et al., 2011; Yoneyama et al., 2013). However, the presence of unidentified novel SLs and/or other germination stimulants cannot be ruled out based on bioassay assessments. Germination assays were reported to be at least 100-fold more sensitive than mass spectrometry (Yoneyama et al., 2010).
The high germination induced by the root powder relative to the root exudates (Tables 1 and 2 and Figures 1 and 2) could be due to differences in conditions under which cowpea plants were grown. The root powder was obtained from plants grown in potted soil while the root exudates were obtained from hydroponically grown plants. However, more subtle interactions involving release of the germination stimulants and/or inhibitors from root powder and the balance between production, transport and exudation of the active compounds from intact plants cannot be ruled out. It is noteworthy that plants roots are considered to be the main sites for SLs biosynthesis and that the cut root technique developed by Berner et al. (1996) is claimed to be more reliable for screening plants for SLs activity than root exudates.
The inconsistent effects of solution pH on germination inducing activity of root exudates of cowpea genotype B301 compared to the consistent effects of pH on germination inducing activity of root exudates of the genotype T100K-901-6 which was invariably significantly higher at pH 7, in line with the results obtained with root powder (Tables 1 and 2), suggest qualitative and /or quantitative differences in composition of the root exudates. Differences in stability and lipophilicity were reported to influence SLs germination inducing activity on Striga spp. (Yoneyama et al., 2010). The likelihood of qualitative and quantitative differences in composition of the root exudates is further substantiated by the high germination response of Striga pearl millet strain and its increase with exudates volume compared to the relatively low germination response of the sorghum strain and its decrease with increasing exudates volume (Table 3 and Figures 1 and 2).
Radicle length in Striga seedlings induced by cowpea root powder, irrespective of Striga strain or cowpea genotype, was by far shorter than those induced by the concurrent GR24 control treatments. GR24 at 0.1 mg L-1 elicited seedlings with radicle length of 17.3-25.7 × 10-2 mm and 9.4-19 × 10-2mm in S. hermonthica sorghum strain and its pearl millet congener, respectively. The influence of root powder on radicle length varied with, cowpea genotype, amount of powder and Striga strain. However, minimum and maximum inhibition of radicle length, in both strains, was achieved by powder from the genotypesT100K-1263 and T198K-409-4, respectively (Tables 4 and 5).
The proportions of short radicles, irrespective of Striga strain or cowpea genotype, increased with increasing amount of powder (Tables 4 and 5). For S. hermonthica sorghum strain the proportion of samples eliciting seedlings with short radicles was 8.3, 41.7 and 75% at powder amounts of 5, 10 and 15 mg/well, respectively. The corresponding figures for the pearl millet strain were 16.7, 75 and 83.3% thus suggesting that the pearl millet strain is more prone to radicle shorting than its sorghum congener. Shortening of Striga radicles due to curtailment of cell extension and/or precocious initiation of haustoria away from host roots, may, as observed with Desmodium spp. for S. hermonthica and a variety of resistant sorghum genotypes for S. asiatica, lessen the frequency of attachment and subsequently parasitism (Riopel and Timko, 1995; Khan et al., 2002, 2008).
Radicle length in seedlings from seeds induced to germinate with root exudates often mirrored imaged the germination inducing activity of the exudates (Figures 1 to 4). Synonymous with germination, radicle length pro-gressively increased with sampling time and decreased with exudates volume for Striga sorghum strain, but, conversely, both germination and radicle length increased with sampling time and exudates volume for the pearl millet congener (Figures 3 and 4).
The decrease in germination inducing activity and reduction in radicle length in Striga sorghum strain with increasing exudates volume corroborate the results obtained in this study with the root powder (Table 1 and Figures 1 and 2) and suggest interactions involving allelochemicals as pre and post-germination and radicle growth inhibitors. A similar shorting in S. hermonthica radicle, on exposure to root exudates of Desmodium uncinatum (Jacq), attributable to allelochemicals including di-C-glycosylflavone, was reported by Khan et al. (2008). The contrasting effects of cowpea root exudates on germination and radicle length of the pearl millet S. hermonthica strain are of interest and suggest high sensitivity to the germination stimulants and/or low sensitivity to inhibitors in the roots exudates. However, the possibility of involvement of different stimulants in the germination and radicle length of the two S. hermonthica strains cannot be ruled out and is consistent with the notable host specificity of the two strains previously reported by Wilson-Jones (1955). Recent studies showed that a plant species may produce several strigolactones with differing germination inducing activity and that the stereochemistry of a stimulant plays a crucial role in germination and in host specificity (Matusova et al., 2005).
The results of the present study revealed that germination inducing activity and effects on radicle length of cowpea root powder and exudates are influenced by genotype and the parasite strains. Furthermore, the results suggest that selection of cowpea genotypes for trap cropping should be based on initial laboratory screening for stimulant production. However, for intercropping growth vigour and habit, among other factors, have to be taken into consideration.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests
ACKNOWLEDGMENTS
The authors are indebted to JST/JICA SATREPS (Japan Science and Technology Agency, Tokyo Japan and International Cooperation Agency, Tokyo, Japan) for financial support.
REFERENCES
|
Awad AA, Sato D, Kusumoto D, Kamioka H, Takeuchi Y, Yoneyama K (2006). Characterization of strigolactones, germination stimulants for the root parasitic plants Striga and Orobanche, produced by maize and sorghum. Plant Growth Regul. 48:221-227. |
|
|
Babiker AGT (2007). Striga: The spreading scourge in Africa. Regulation of Plant Growth and Development 42:74-87. |
|
|
Babiker AGT, Ma Y, Sugimoto Y, Inanaga S (2000). Conditioning period, CO2 and GR24 Influence ethylene biosynthesis and germination of Striga hermonthica. Physiol. Plant. 109:75-80. |
|
|
Berner K, Ikie FO, Aigbokhan EI (1996). Methods for soil infestation with Striga hermonthica seeds. Agron. J. 88:33-37. |
|
|
Cardoso C, Charnikhova T, Jamil M, Delaux PM, Verstappen F, Amini M, Lauressergues D, Ruyter-Spira C, Bouwmeester H (2014). Differential activity of Striga hermonthica seed germination stimulants and Gigaspora rosea hyphal branching factors in rice and their contribution to underground communication Plos One 9 e1410(9):1-11. |
|
|
Ejeta G, Butler LG, Babiker AGT (1993). New approaches to the control of Striga. Striga Research at Purdue. Agricultural Experimental Station. Purdue Univ. Res. Bull. 991:27. |
|
|
Fujii Y, Shibuya T, Nakatani K, Itani T, Hiradate S, Parvez MM (2004). Assessment method for allelopathic effect from leaf litter leachates. Weed Biol. Manage. 4:19-23. |
|
|
Khan Z, Hassanali A, Overholt W, Khamis TM, Hopper AM, Picket JA, Wadhams LJ, Woodcock C (2002). Control of Witchweed Striga hermonthica by Intercropping with Desmodium spp., and the mechanism defined as allelopathic. J. Chem. Ecol. 28:1871-1885. |
|
|
Khan ZR, Pickett, JA, Hassanali A, Hooper AM, Midgea CAO (2008). Desmodium species and associated biochemical traits for controlling Striga species: Present and future prospects. Weed Res. 43:302-306. |
|
|
Kureh I, Kamara AY, Tarfa BD (2006). Influence of cereal-legume rotation on Striga control and maize grain yield in farmers, fields in Northern Guinea Savanna of Ngeria. J. Agric. Rural Dev. Trop. Subtrop. 107:41-54. |
|
|
Matusova R, Rani K, Verstappen FWA, Franssen MCR, Beale MH, Bouwmeester HJ (2005). The strigolactone germination stimulants of the plant-parasitic Striga and Orobanche are derived from the carotenoid pathway. Plant Physiol. 139:920-934. |
|
|
Muller S, Hauck C, Schildknecht H (1992). Germination stimulants produced byVigna unguiculata Walp cv. Saunders Upright. J. Plant Growth Regul. 11:77-84. |
|
|
Parker C, Reid DC (1979). Host specificity in Striga species-some preliminary observations. In: Supplement to Proceedings, 2nd International Symposium on Parasitic Weeds. Raleigh USA. (eds LJ Musselman EDITOR, AD Worsham EDITOR & R Eplee) North Carolina State University, Raligh. pp. 79-90. |
|
|
Parker C, Riches CR (1993). Parasitic Weeds of the World: Biology and Control. CAB International, Wallingford, UK. |
|
|
Riopel JL, Timpko MP (1995). Haustorial initiation and differentiation. In: Parasitic Plants (eds MC Press Editor & JD Graves Editor). Chapman and Hall London, UK. 332 p. |
|
|
Sato D, Awad AA, Takeuchi Y, Yoneyama K (2005). Confirmation and quantification of strigolactones, germination stimulants for root parasitic plants Striga and Orobanche, produced by cotton. Biosci. Biotechnol. Biochem. 69:98-112. Spallek T, Mutuku, JM, Shirasu K (2013). The genus Striga: A witch profile. Molec. Plant Pathol. 69(1):98-102. |
|
|
Ueno K, Nomura S, Muranaka S, Masaharu M, Takikawa H, Sugimoto Y (2011). Ent-2/-epi-orobanchol and its acetate as germination stimulants for Striga gesnerioides seeds isolated from cowpea and red clover. J. Agric. Food Chem. 59:10485-10490. |
|
|
Welsh AB, Mohamed IK (2011). Genetic diversity of Striga hermonthica populations in Ethiopia. Evaluating the role of Geography and host specificity in shaping population structure. Int. J. Plant Sci. 172:773-782. |
|
|
Wilson-Jones K (1955). Further experiments on witchweed control II. The existence of physiological strains of Striga hermonthica. Empire J. Exp. Agric. 23:206-213. |
|
|
Yoneyama K, Awad AA, Xie X, Yoneyama K, Takeuchi Y (2010). Strigolactones as germination stimulants for root parasitic plants. Plant Cell Physiol. 51:1095-1103. |
|
|
Yoneyama K, Ruyer-Spira C, Bouwmeester H (2013). Induction of germination. In: Parasitic Orobanchaceae Parasitic Mechanisms and Control Strategies (eds DM Joel, J Gressel, & LJ Musselman), Springer Verlag, Berlin Heidelberg, Germany pp. 164-194. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


