Full Length Research Paper

ABSTRACT
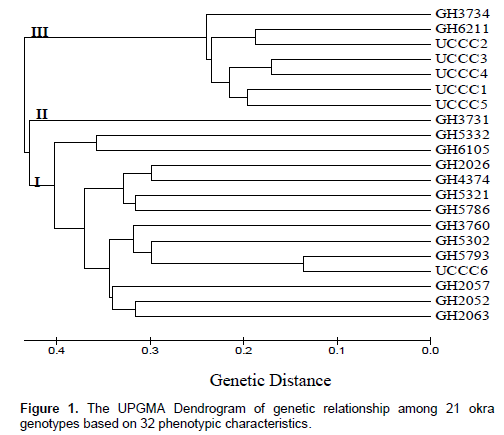
There is low production of okra in Ghana due to lack of improved varieties and biotic constraints. This study was conducted to characterize okra genotypes to predict genetic variation in the crop. Field trial was conducted to determine genetic variability in 21 okra genotypes. The experiment was based on the randomized complete block design (RCBD) involving planting distance of 0.6 × 0.6 m. Thirty-one quantitative and qualitative data were used to generate a dendrogram. Variations in leaf shape, leaf rib colour, petiole colour, petal colour, colour of the darkest ridges and stem colour were distinctive among the okra genotypes. The mean plant height, canopy diameter, leaf length and breadth, petiole length, internode length, number of branches, days to 50% flowering and fruit yield differed significantly (p ≤ 0.05) among the 21 okra genotypes. These were discriminated into three clusters in a dendrogram with GH3731 as the most diverse. UCCC1, UCCC2, UCCC3, UCCC4 and UCCC5 appeared genetically similar with low fruit yield but early maturity. However, GH5332 had a significantly (p ≤ 0.05) the highest fruit yield of 11.88 t ha-1 but late maturing. UCCC5 or similar genotypes with early maturity trait can be hybridized with GH5332 to improve the yield and earliness.
Key words: Breeding, germplasm, genotypes, genetic diversity and hybridization.
INTRODUCTION
MATERIALS AND METHODS
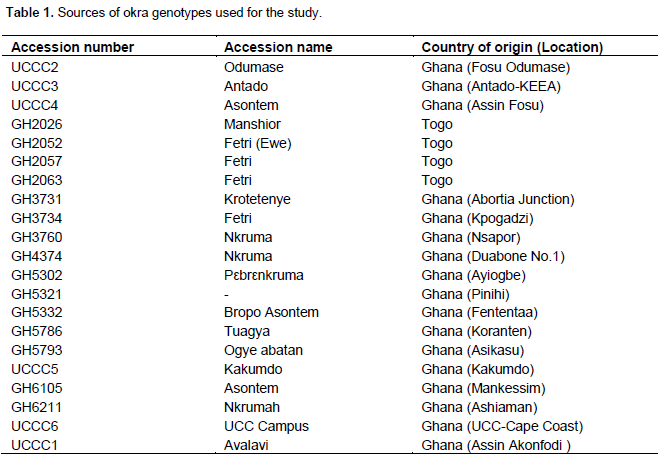

RESULTS AND DISCUSSION
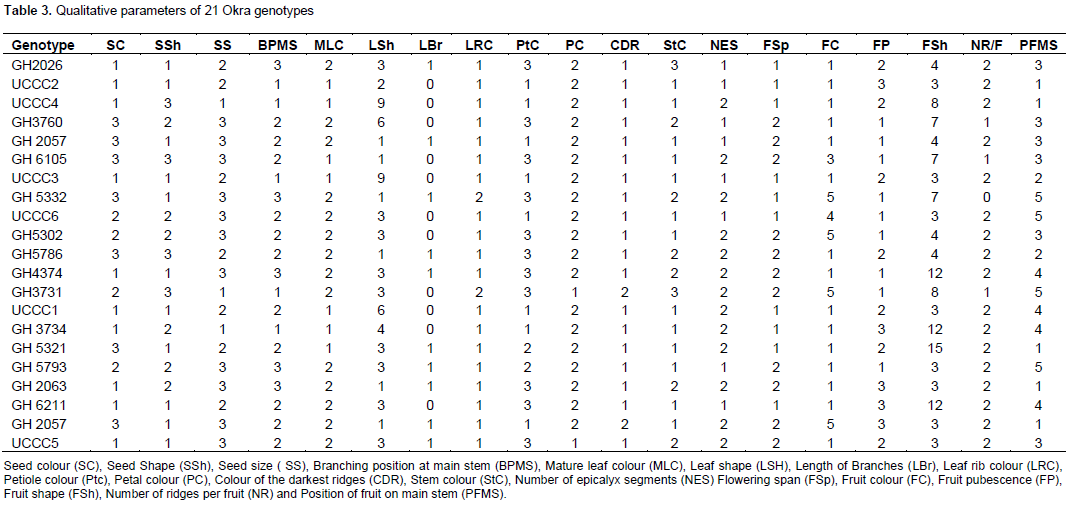
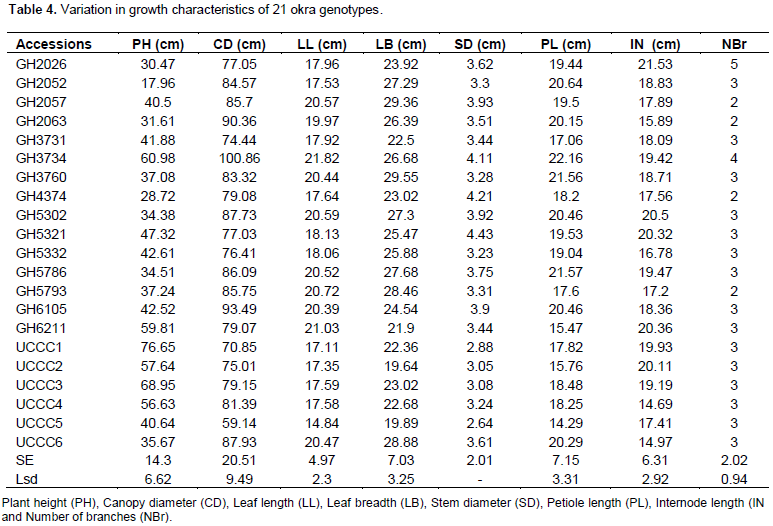
The leaves of okra serve as the main sites for photosynthesis, an increase or a decrease in their size could affect production of assimilates in the crop. Larger size leaves in any okra genotype may have higher ability to intercept solar radiation to assume higher photosynthetic capacity, which may enhance growth and crop yield. GH3734 had the widest canopy diameter of 100.86 cm compared to the least of UCCC5 (59.14 cm). The highest leaf length of 21.82 cm was produced by GH3734 and the lowest of 14.84 cm was associated with UCCC5. The mean leaf breadth of 29.55 cm was the highest observed for GH3760 compared to the least average leaf breadth of 19.89 cm for UCCC5 (Table 4). According to Ahiakpa (2013), an increased leaf area index and a resultant higher fraction of intercepted radiation and its utilization efficiency may increase crop yield. Significant (p < 0.05) correlations were observed between leaf length and canopy diameter (r = 0.72), breadth and canopy diameter (r = 0.53), leaf length and breadth (r = 0.65), petiole length and canopy diameter (r = 0.49) and stem diameter and canopy diameter (r = 0.70) in the okra germplasm could be determinants for plant vigour and yield indicators. GH2026 had the longest internodes of 21.53 cm per plant compared with the shortest for UCCC4 (14.69 cm per plant).
The mean petiole length of 20.64 cm was the highest for GH2052 compared with that of the lowest average petiole length of 14.29 cm for UCCC5. GH2026 had highest mean internodes (21.53 cm) and number of branches (5), respectively. However, GH2057, GH2063, GH4372 and GH5793 had the least number of 2 branches per plant. Variations in petiole length, leaf size, canopy diameter, number of branches and stem diameter may have implications for crop yield and stability to control lodging.
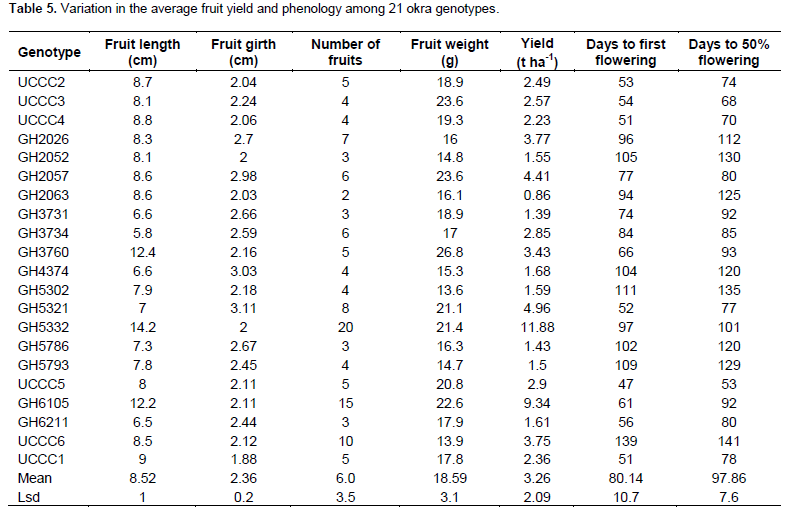
According to Ariyo and Odulaja (1991), variability in okra germplasm is more prominent in days to flowering, plant height and various fruit characteristics and these traits could be important in differentiating varieties of A. esculentus. Similarly, in the current study, fruit length, girth and weight as well as the days to first flowering and days to 50% flowering differ significantly (p < 0.05) among the 21 genotypes of okra (Table 5). However, UCCC1, UCCC2, UCCC3, UCCC4, and UCCC5 were very similar in early flowering and days to 50% maturity as well as average fruit number, weight and size. Generally, all okra genotypes with high vegetative growth delayed flowering and maturity. UCCC5 was first to flower at 47 days and attained 50% flowering at 53 days, respectively after sowing seeds, which were significantly early compared to others. On the contrary UCCC6 was very late to first flower at 139 days and 50% flowering at 141 days, which were significantly (p < 0.05) different from all the other okra genotypes.
In this study, the significantly (p < 0.05) high yielding okra genotype, GH5332, produced 20 fruits per plant, with the highest fruit weight of 11.88 t ha-1, which compared well with the size of the fruits (mean fruit length of 14.2 cm, girth 20 mm, and weight of fruit per plant of 21.6 g) among the okra genotypes. However, GH5332 is late maturing with 50% flowering at 101 days. Indeed, Esthiet and Brisibe (2015) reported that fruit length, pod number and pod weight are the most important determinants of production or yield in okra. It has been suggested that the number of days and plant height at flowering are controlled by the same genetic variables (Choudhary et al., 2006; Hussain et al., 2006).
It is critical to consider early maturity in the phase of erratic rainfall as essential trait to complement yield for hybridization to produce climate-smart okra genotypes. Therefore, UCCC5 with very early flowering and fruiting traits can be hybridized with the high yielding but late maturing genotypes of GH5332 to improve the crop. A successful cross between unrelated varieties may result into an array of elite genotypes from which advantageous agronomic line may be selected (Ali et al., 2014).
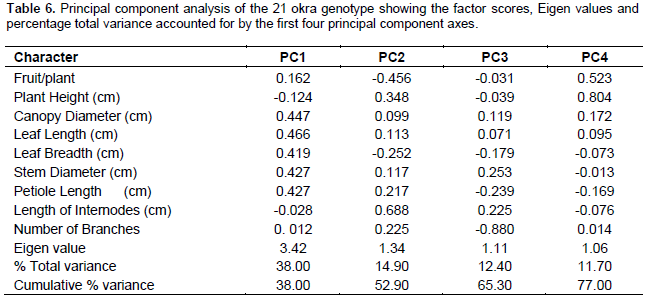
The variation in the quantitative characteristics which accounted for the total variance includes number of fruits per plant, mean plant height, canopy diameter, leaf length, breadth, stem diameter, petiole length, internode length, and number of branches. The proportion contributed by each quantitative variable to determine the total variation within each Principal Component (PC) axis is shown in Table 6. The variations in the quantitative characters contributed significantly (Eigen vector ≥ 0.2) to the variation within each of the four PC-axes as 38.00, 14.90, 12.40 and 11.70% for PC1, PC2, PC3 and PC4, respectively. The cumulative proportion of variation explained by the first four PC-axes, 77.00% (Table 6) compared well with observations made by Campos et al. (2005) and Ogunbayo et al. (2005) that the PC-axes contributed 76.62 and 64.5% variations, respectively. Similarly, Ahiakpa et al. (2013) reported that the first four PC-axis contributed 82.97% of the variations in okra. The remaining six axes in the current study accounted for only 23.00% of the total variation. Indeed, canopy diameter, leaf length, breadth, stem diameter and petiole length contributed to the variation in PC1. Plant height, length of internodes and number of branches accounted for the variations observed in PC2 and fruit per plant as well as plant height contributed to the variations in PC4. These variations may suggest the existence of genetic diversity in okra that can be harnessed to improve the crop. Similar observation was made by Yonas et al. (2014).
CONCLUSIONS
RECOMMENDATION
CONFLICT OF INTERESTS
ACKNOWLEDGEMENT
REFERENCES
|
AdeOluwa OO, Kehinde OB (2011). Genetic variability studies in West African okra (Abelmoschus caillei). Agric. Biol. J. N. Am. 2(10):1326-1335. |
|
|
Ahiakpa JK, Kaledzi PD, Adi EB, Peprah S, Dapaah HK (2013). Genetic diversity, correlation and path analyses of okra (Abelmoschus spp. (L.) Moench) germplasm collected in Ghana. Int. J. Dev. Sust. 2(2):1396-1415. |
|
|
Ali AS, Shah H, Gul R, Ahmad H, Nangyal H, Sherwani KS (2014). Morpho-Agronomic Characterization of Okra (Abelmuscus esculentus L.). World App. Sc. J. 31(3):336-340. |
|
|
Aliyu B, Fawole I (2001). Inheritance of pubescence in crosses between V. unguiculata and V. rhomboidea. Nig. J. Genet. 15:9-14. |
|
|
Aremu CO, Adebayo MA, Ariyo OJ, Adewale BB (2007). Classification of Genetic diversity and choice of parents for hybridization in cowpea Vigna unguiculata (L.) Walp for humid savanna ecology. Afr. J. Biotechnol. 6(20): 2333-2339. |
|
|
Ariyo OJ, Akinova ME, Fatokun PE (1987). Plant character correlation and path analysis of pod yield in okra. Euphytica 36:677-686. |
|
|
Ariyo OJ, Odulaja A (1991). Numerical analysis of variation among accessions of okra. (A. esculentus [L.] Moench). Malvaceae. Ann. Bot. 67:527-531. |
|
|
Asare-Bediako E, Addo-Quaye AA, Bi-Kusi A (2014a). Comparative efficacy of phytopesticides in the management of Podagrica spp and mosaic disease on okra (Abelmoschus esculentus L.). Am. J. Expt. Agric. 4(8):879-889. |
|
|
Bisht IS, Mahajan RK, Rana RS (1995). Genetic diversity in South Asian okra (Abelmoschus esculentus) germplasm collection. Ann. Appl. Biol. 126:239-550. |
|
|
Campos ET, Espinosa MAG, Warburton ML, Monter AV (2005). Characterisation of mandarin (Citrus spp) using morphological and AFLP markers", Interciencia 30(11):1-14. |
|
|
Chattopadhyay A, Dutta S, Chatterjee S (2011). Seed yield and quality of okra as influenced by sowing dates. Afr. J. Biotechnol. 10:5461-5467. |
|
|
Choudhary UN, Khanvilkar MH, Desai SD, Prabhudesai SS, Choudhary PR (2006). Performance of different okra hybrids under North Konkan coastal zone of Maharashtra. J. Soils Crops 16:375-378. |
|
|
Cudjoe AR, Kyofa-Boamah M, Nkansah GO, Braun M, Owusu S, Adams E, Monney E, Attasi R, Owusu P, Sarpong S (2005). Commercial Okra Production in Ghana - Good Agricultural Practices/Code of Practice and IPM Strategies: In: Kyofa-Boamah M, Blay E, Braun M, Kuehn A (eds). Handbook of Crop Protection. Recommendations in Ghana, Ministry of Food and Agric. Accra pp. 75-92. |
|
|
Esthiet JA, Brisibe AE (2015). Morphological Characterization and Yield Traits Analysis in Some Selected Varieties of Okra (Abelmoschus Esculentus L. Moench). Adv. Crop. Sci. Technol. 3(5):1-5. |
|
|
Hussain S, Muhammad NS, Shah A, Iqbal Z (2006). Response of okra (Abelmoschus esculentus) cultivars to different sowing times. J. Agric. Biol. Sci. 01:55-59. |
|
|
IPGRI (1991). Okra descriptor, Diversity for Development. Int. Plant Gen. Res Inst. Rome. |
|
|
Nwangburuka CC, Kehinde OB, Ojo DK, Denton OA, Popoola AR (2011). Morphological classification of genetic diversity in cultivated okra, Abelmoschus esculentus (L) Moench using principal component analysis (PCA) and single linkage cluster analysis (SLCA). Afr. J. Biotechnol. 10(54):11165-11172. |
|
|
Ogunbayo SA, Ojo DK, Guei R, Oyelakin OO, Sanni KA (2005). Phylogenetic diversity and relationships among 40 rice accessions using morphological and RAPDs techniques. Afr. J. Biotechnol. 4(11):1234-1244. |
|
|
Ofoefule SI, Chukwu AN, Anayakoha A, Ebebe IM (2001). Application of Abelmoschus esculentus in solid dosage forms 1: use as binder for poorly water soluble drug. Indian J. Pharm. Sci. 63:234-238. |
|
|
Omonhinmin CA, Osawaru ME (2005). Morphological characterization of two species of Abelmoschus: Abelmoschus esculentus and Abelmoschus caillei. Genet. Resour. Newsl. 144:51-55. |
|
|
Owusu-Sekyere JD, Alhassan M, Nyarko BK (2011). Assessment of climate shift and crop yields in the Cape Coast area in the Central Region of Ghana. ARPN. J. Agric. Biol. Sci. 6(2):49-54. |
|
|
Perdosa J (1983). The morphological characters of okra introductions. Horticult. Bras. 1(1):14-23. |
|
|
Rahman K, Waseem M, Kashif MS, Jilani M, Kiran G (2012) Performance of different okra (Abelmoschus esculentus L.) cultivars under the agro-climatic conditions of Defra Ismail Khan. Pak. J. Sci. 64:316-319. |
|
|
Saifullah M, Rabbani MG (2009). Evaluation and Characterization of Okra (Abelmoschus esculentus L. Moench.) Genotypes. SAARC J. Agric. 7(1):92-99. |
|
|
Siesmonsma JS, Kouame C (2004). Vegetables. In: Plant Resources of Tropical Africa 2 (Grubben, G.J.H. & Denton, O.A., Eds.) PROTA Foundation, Wageningen, Netherlands and Backhuys Publishers, Leinden, Netherlands and CTA, Wageningen, Netherlands pp. 20-29. |
|
|
Tamura K, Dudley J, Nei M, Kumar S (2007). MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. (Accessed on 28th March, 2016). Mol. Biol. Evol. 24:1596-1599. |
|
|
Verma VD (1993). Collecting eggplant and okra in Madhya Pradesh and Maharashtra. IBPGR. Newsletter for Asia Pacific and Oceania 13:14-15. |
|
|
Wamanda DT (2007). Inheritance studies in collected local okra (Abelmoschus esculentus L. Moench) cultivars. In: Combining ability analysis and heterosis on diallel cross of okra. Afr. J. Agric. Res. 5:2108-2155. |
|
|
Yonas M, Garedew W, Debela A (2014). Maltivariate analysis among Okra (Abelmoschus esculentus L. Moench) Collections in South Western Ethiopia. J. Plant Sci. 9(2):43-50. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


