Full Length Research Paper

ABSTRACT
Changes in nutrients elements uptake, physiological characteristics, as well as, correlations analysis for these traits to each other are good factors to study the effects of drought on sweet pepper (Capsicum annuum L. cv., commercial hybrid) and disclose physiological reaction to several specific levels of stress from water deficit. Water scarcity processing was carried out by irrigating the sweet pepper plants with 20, 40 and 60% of water deficit to reach soil pot capacity, plus to 100% of water capacity (control). The results showed that the concentrations of nutrients of the studied sweet pepper grown in pots under different water treatments varied greatly depending on the levels of water stress except C. Resulting data of the growing season’s experiments revealed that, the four water stress levels had a highly significant effect on all the four traits under this study. The content of proline in plant had risen significantly by drought stress treatments (80, 60 and 40% of pot capacity). All water stress treatments led to high significant changes in the accumulation of vitamins and enzymes, that is, peroxidase, ascorbate peroxidase, catalase and superoxide dismutase. Also, the associations of correlation between all physiological traits had been reported in results.
Key words: Capsicum annuum, water deficit, pigments, proline, antioxidant enzymes pigments
INTRODUCTION
Water stress is considered as one of the serious limitations of agricultural production especially in water scarcity regions (Showemimo and Olarewaju, 2007) including Kingdom of Saudia Arabia. The high temeperature and scarcity of water in the Kingdom of Saudi Arabia have led to increased competitions for water resources among various sectors (domestic, agricultural and industry uses). Consequently, the most critical problem to address in this concern of water scarcity is the economic and efficient irrigation systems for increased production of sweet pepper and reduced import from outside countries.
Sweet pepper (Capsicum annuum L.), is one of the important prominent solanaceous crop plants widely cultivated in tropical and subtropical plantations. The growth impairment, low quantity and quality of production are caused by several factors. On top of these factors is water scarcity. Plants utilize a range of physiological and biochemical techniques to overcome the negative effect of water deficit. Many studies in different areas around the world on many species of solanaceae and/or pepper and the effect of water deficit have been carried out (Showemimo and Olarewaju, 2007; Gaveh et al., 2011; Karam et al., 2011) demonstrating that solanaceae crops can be grown at moderate water stress levels with no significant impact on their yield. There is a complicated interrelationship between production, physiological processes and biomass yield to various levels of water deficiency, particularly in semi-arid solanaceous crops (Kirnak et al., 2001).
Optimizing promising features, high crop yields and water conservation are some of the benefits of boosting water management, as effective strategy in farming activities ought to be identified in order to use constrained water efficiently in semi-arid areas. Abdel-Ghany et al. (2004) suggested that the defined cultivars with high or medium yield in addition to favorable specific adaptation characteristics are beneficial for the stress of drought and could be integrated to improve its effectiveness. Many authors reported that one remarkable approach is to control water efficiency and enhance its performance to ensure optimum use of water through designing an effective irrigation schedule (Molden, 1997; Molden et al., 2001). A complete and accurate understanding of the mechanisms of water scarcity on plant species is essential for improved productivity and breeding programs attempts in semi-arid areas. Kurniawati (2014) observed that crop production, foliage, medium moisture, relative water content, total chlorophyll decreased throughout drought,whereas proline and polyamines elevated and exceeded high levels within 14 to 21 days of treatment. Many researches have been developed in the previous thirty years, covering subjects from crop techniques to water status control under moisture stress, the physicochemical feasibility inherent plant reaction and its effect through drought stress (Chaves, 1991; Cornic and Massacci, 1996; Kurniawati, 2014).
Simple correlation coefficient for all possible combinations among all studied traits is a statistical methodology used to calculate some relationships among many different variables and give the relative importance for these association. It could also be useful as indicators of the more important variables under consideration. This mthodology was carried out according to Snedecore and Cochran (1980).
This research was conducted to investigate the changes in some nutrient elements (C, K, P, and N) uptake and physiological characteristics, that is, chlorophylls, carotenoids, antioxidant enzymes and proline activity under effect of water stress in sweet pepper, as well as, to know the relationship among all studied traits to each other using simple correlations coefficient analysis.
MATERIALS AND METHODS
Seeds of pepper (commercial hybrid) were implanted in a greenhouse nursery till the plants formed four leaves and attained a shooting height of 15-20 cm. After that, plants were transferred to bigger containers (30 cm height and 30 cm top diameter) after being filled with (1:1) sandy loam in the open field.
Irrigation began in the end of February and the harvest was done during July. Agricultural procedures and crop protection were carried out in compliance with the regional known procedures. Randomized Complete Block Design (RCBD) was utilized with triple replicates. To minimize variation, provide unbiased evaluation and achieve a good experiment, we made use of randomization. Therefore, the arrangement of all pots had been changed (haphazardly) every 7-9 days interval.
Irrigation treatments
The water stress condition had been applied on many solanaceae plants (sweet pepper) which has been subjugated to investigation. The evaporation potential was determined utilizing pan evaporation as defined by Doorenbos (1975). Treatments included four levels of water proceedings, viz; control treatment, normal irregation (W1), and the other three treatments were water deficit to reach soil pot capacity by 20% ( W2), 40% (W3) and 60% (W4).
Physiological parameters
The content of C, K, P and N elements were measured based on Richards (1969) method. Chlorophyll and carotenoids were estimated with an Ultrospec 2100 pro spectrophotometer (Amersham Biosciences), and then calculated according to Arnon (1949). Proline was estimated based on Bates et al. (1973) method.
Superoxide dismutase activity (SOD EC 1.15.1.1) was estimated according to Beyer and Fridovich (1987). Peroxidase (POX) activity was measured according to Sakharov and Ardila (1999). Catalase (CAT EC 1.11.1.6) activity was estimated spectrophotometrically according to the method of Aebi (1974). Ascorbate peroxidase (APX EC 1.11.1.11) was determined according to Wang et al. (1991).
Statistical analysis
Analysis of variance (ANOVA), LSD, and simple correlation coefficient were carried out according to Snedecore and Cochran (1980) method using MSTAT software (Nissen et al., 1985).
RESULTS AND DISCUSSION
The analyzed physiological and biochemical characteristics, that is, C, K, P, N, chlorophyll a, b, and total chlorophyll, carotenoids, proline, POX, ascorpate POX, CAT and SOD of sweet pepper variety under four levels of water deficit are presented in Table 1. The ANOVA results revealed that the four water stress levels had high significant effect on all previous investigated traits except the content of C element in plant.
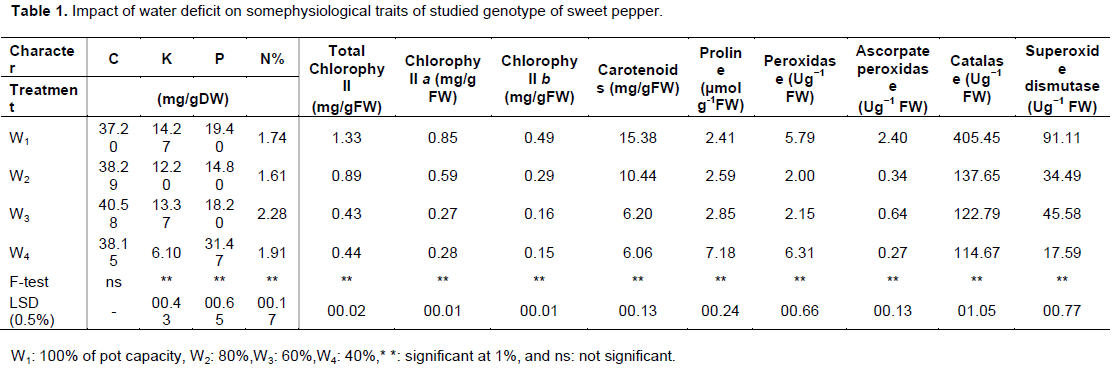
Regarding the content of Potassium in the plant, the highest value (14.27) was recorded in the high level of irrigation 100% (W1), while lowest (6.10) was recorded from (W4) when the level of pot capacity decresed to 40%. In the opposite of the trend of other elements, the largest Phosphorous content (31.47) was obtained by the lowest treatment in the levels of pot capacity (W4), while the other values varied gradually for the four treatments. Generally, the results indicated that the Phosphorous increased with decreasing stress levels from 100% normal irrigation to 40% pot capacity. Percentage Nitrogen content was affected significantly at 0.01 by water deficit where the level of pot capacity (60%) gave the highest value (2.28), while the lowest value was 1.61 with least significant difference of 0.17.
The results agree with Kirank et al. (2001) who suggested that concentrations of nutrients in the solaneous leaves decreased under various irrigation levels. Furthermore, Almohisen (2015) reported similar patterns in tomato content of C, K, P and N. This could mainly be because of water deficit on function of two physiological processes. The first one is increasing number of cells through cell division and the second is increasing cell size through cell elongation and turgidity, then the absorption of elements. Therefore, these processing pushed the plants towards different growth behavior. Many investigators have studied the role of many elements in plants and their physiological functions, in addition to a very important role in plant growth and nutrition (Almohisen, 2015; Azam et al., 2016; Buschmann et al., 2000; Delfine et al., 2001; El-Ghany et al., 2012).
The analysis of variance for green and organic pigments accumulation data of the growing season’s experiments revealed that, the four treatments of drought had a highly significant effect on these four previous characters under this study. Results showed that, the detraction of pot capacity from (100%) to (40%) caused a reduction of total chlorophyll from (1.33) to (0.44), respectively with least significant difference (0.02), taking into consideration the value of 60% pot capacity of (0.43). The same actual trend as total chlorophyll was observed where the W1 (100% normal irrigation) produced the highest figure (0.85), while the water stress treatment, either 60 or 40% gave the smallest accumulation (0.28) and (0.27) of chlorophyll a, respectively, without any significant difference between the two lowest values. As regards chlorophyll b, the highest accumulation (0.49) was obtained from adding water to maximization (100%) of pot capacity; however, the addition of water to 40% and/or 60% of pot capacity gave the lowest value (0.15 mg/g FW) and (0.16), respectively, with non-significant difference between the lowest amounts. Whereas the least sigificant differences among data of chlorophyll a and/or chlorophyll b was (0.01), water deficit treatments affected the total accumulation of carotenoids significantly (P< 0.01). Carotenoids dramatically decreased responsive to water deficit from (15.38) in W1 (100% normal irrigation) to (6.06) in W4 (60%).
Kirnak et al. (2001) reported that the overall amount of chlorophyll in drought regimes were reduced by 55% relative to treatment. The chlorophyll indicators were used as subordinate predictors of chlorophyll and concentrations of N (Liu et al., 2006). Okunlola et al. (2017) showed that moderate and severe drought induced significant reduction in carotenoid, chlorophyll a, b and total chlorophyll content of the study plants at the vegetative stage.
Regarding the proline content, results showed that, the water deficit reaching soil capacity of pot proceedings by 20, 40 and 60% had high significant impact on increased elevation of proline compared to control (normal irrigation). The results revealed that, proline content was 2.41, 2.59, 2.85 and 7.18 in W1, W2, W3 and W4, respectively. With least significant difference (0.24) where the highest value obtained by W4 followed by W3, then with W2 and W1 (without insignificant difference between W1 and W2). In terms of water strain, proline elevation typically occurs in cytosol, that plays an important role in cytoplasmic osmotic adjustment (Anjum et al., 2012). El Sayed (1992) reported that, the proline dehydrogenase and oxidase activities declined as the water deficit level escalated. In much more serverely strained roots and leaves, activity of proline dehydrogenase were inhibited by around 85%. Alaei et al. (2012) recommended that, using biological fertilizers and fertilizers which contain amino acids could increase the proline density in wheat under drought conditions.
Data in Table 1 showed the highly significant changes in vitamins and enzymes accumulation, that is, POX, ascorpate POX, CAT and SOD as impact of water deficit. Both low level of pot capacity (40%) and the highest one (100%) gave the highest values of Peroxidase accumulation (6.31) and (5.79), respectively, without any significant difference. Moreover, the lowest values (2.00 and 2.15) were obtained from 80 and 60% of pot capacity, respectively, with no significant difference among them. The highest value of ascorpate peroxidase accumulation (2.40) was obtained from the high level of irrigation (100 pot capacity), while the lowest value of (0.27) was recorded when sweet pepper was irrigated with the low level of pot capacity (W4), taking into consideration that the second treatment (80% pot capacity) gave (0.34) with lowest significant difference (0.13). Exposing sweet pepper plants to water deficit during its life causes a big reduction in catalase accumulation up to about 28% which was (405.45) at normal irrigation (W1), while it highly decreased at 40% of pot capacity (W4) which gave (114.67). On the other hand, superoxide dismutase accumulation in sweet pepper plants values varied highly significantly (0.77 L.S.D.), where the elevation of SOD reached 91.11, 45.58, 34.49 and 17.59 in W1, W3, W2 and W4, respectively.
These differences among studied traits responsive to water strain reflects the role of these components as biological regulators in the plant (El-Saidy et al., 2011; Ahanger et al., 2016), and might even be due to the role of plant metabolism system, in addition to the partial retardation of co-vitamins with some interaction in the generation of reactive oxygen species (Ratnakar and Rai, 2013; Ichwan et al. (2017) concluded that, drought stress at 75% FC can readily lower the agronomical and physiological characteristics of red chilli. Sayyari and Ghanbari (2012) reported that, super absorbent polymers, irrigation doses and their interrelationships significantly affected sweet pepper biological and physiological metrics. By increasing irrigation intervals and drought stress, growth parameters, and chlorophyll were reduced and proline content increased. Anjum et al. (2012) declared that water deficit caused numerous changes in the activity of antioxidant enzymes. Futhermore, peroxidase and catalase activity were estimated to have increased or stabilized in the early deficit of water and then lowered as the water deficit progressed. Tahi et al. (2008) noticed that increasing the antioxidants correlated with water deficit tolerance. The SOD is deemed to be the first point of defence from ROS that catalyzed the radical superoxide (O2-) to O2 and H2O2 that are also satiated by various antioxidants.
Correlation analysis
Recorded matrix data in Table 2 show the simple correlation coefficient among some physiological traits of sweet pepper under water stress which are arranged in a correlation matrix.
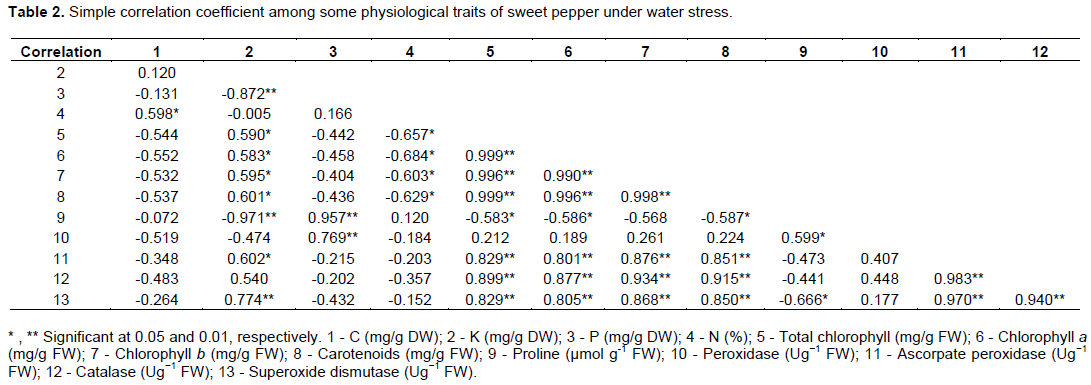
Statistical analysis of correlation coefficient revealed many types of simple correlation coefficients (either positive or negative direction) and (weak or very strong relation).
Positive correlation coefficient among all studied characterististics
Correlation estimates between K and superoxide dismutase was (0.774), to P and proline was (0.957), and peroxidase was (0.769), which were observed as positive and highly significant relations.
Regarding the three green and one organic pigments, they take the same trend, that is, positive and highly significant relations, where total chlorophyll with both chlorophyll a, chlorophyll b, carotenoids, ascorpate peroxidase, catalase and superoxide dismutase gave (0.999), (0.996), (0.999), (0.829), (0.899) and (0.829), respectively. Also, the correlation between chlorophyll a with both chlorophyll b was (0.990), carotenoids was (0.996), ascorpate peroxidase was (0.801), catalase was (0.877) and superoxide dismutase was (0.805), which, were positive and highly significant. In addition, the correlation between chlorophyll b with both carotenoids, ascorpate peroxidase, catalase and superoxide dismutase were highly positive expressive correlations: (0.998), (0.876), (0.934) and (0.868), respectively.
As respects correlation among the organic pigments trait; carotenoids and both ascorpate peroxidase was (0.851), catalase was (0.915) and superoxide dismutase was (0.850), with all these relations being positive and highly significant correlations. For proline, there is only one positive and highly significant correlation with P content traits as earlier mentioned.
Also, the remaining coefficients that were positive and highly significant were (0.983) between ascorpate peroxidase and catalase, (0.970) for ascorpate peroxidase and superoxide dismutase, and (0.940) for catalase with superoxide dismutase. In this respects, we can say that, the positive and highly significant relations represent about 31% of data.
Moreover, positive and only significant correlation estimates were observed between C and N % which gave (0.598).
On the same positive direction of relationships, the only significant correlations were (0.590), (0.583), (0.595), (0.601) and (0.602) between K with green and organic pigments, both total chlorophyll, chlorophyll a, chlorophyll b, carotenoids and ascorpate peroxidase, respectively. In the same trend, coefficient of correlation (0.599) was obtained from the relation between proline and Peroxidase.
As regards weak or moderate positive relationship among characters, there is no expressive correlation (at level of 5 and 1%) for correlation coefficient between C with K which gave corelation value (0.120); or K with catalase (0.540). In the same trend, the correlation coefficient between P and N % was not significant and gives the value (0.166). The correlation between N % and proline was (0.120). Focusing on the results of correlation matrix in Table 2, we found that the association between peroxidase with all studied traits was not significant either at 5 or 1% level, except two cases (mentioned before and three negative cases that will be mentioned later), where both total chlorophyll, chlorophyll a, chlorophyll b, carotenoids, ascorpate peroxidase, catalase and superoxide dismutase show correlation values of (0.212), (0.189), (0.261), (0.224), (0.407), (0.448) and (0.177), respectively.
Negative correlation coefficient among all studied characterististics
Conversely, negative correlation coefficient between all studied traits and each other had been observed to be either at probability of 0.01 or 0.05. Results showed just two cases were negative and highly significant correlations between content of K with both P and proline which gave (-0.872) and (-0.971), respectively.
On the same positive direction of relationships, the only significant correlations were (0.590), (0.583), (0.595), (0.601) and (0.602) between K with green and organic pigments, both total chlorophyll, chlorophyll a, chlorophyll b, carotenoids and ascorpate peroxidase, respectively. In the same trend, coefficient of correlation (0.599) was obtained from the relation between proline and Peroxidase.
As regards weak or moderate positive relationship among characters, there is no expressive correlation (at level of 5 and 1%) for correlation coefficient between C with K which gave corelation value (0.120); or K with catalase (0.540). In the same trend, the correlation coefficient between P and N % was not significant and gives the value (0.166). The correlation between N % and proline was (0.120). Focusing on the results of correlation matrix in Table 2, we found that the association between peroxidase with all studied traits was not significant either at 5 or 1% level, except two cases (mentioned before and three negative cases that will be mentioned later), where both total chlorophyll, chlorophyll a, chlorophyll b, carotenoids, ascorpate peroxidase, catalase and superoxide dismutase show correlation values of (0.212), (0.189), (0.261), (0.224), (0.407), (0.448) and (0.177), respectively.
Negative correlation coefficient among all studied characterististics
Conversely, negative correlation coefficient between all studied traits and each other had been observed to be either at probability of 0.01 or 0.05. Results showed just two cases were negative and highly significant correlations between content of K with both P and proline which gave (-0.872) and (-0.971), respectively.
While the correlation associations were negative and significant at 5% only with the content of N % and both total chlorophyll, chlorophyll a, chlorophyll b and carotenoids, which gave (-0.657), (-0.684), (-0.603) and (-0.629), respectively; in the same trend (negatively significant probability at 0.05), the correlation between proline (µmol g-1 FW) and both total chlorophyll, chlorophyll a, carotenoids and superoxide dismutase were (-0.583), (-0.586), (-0.587) and (-0.666), respectively.
On the contrary, there is negative and non-significant correlation either at probability of 0.01 or 0.05 where the values of correlation, that is, (-0.131), (-0.544), (-0.552), (-0.532), (-0.537), (-0.072), (-0.519), (-0.348), (-0.483) and (-0.264) were obtained from relations between the content of C and both P, total chlorophyll, chlorophyll a, chlorophyll b, carotenoids, proline, peroxidase, ascorpate peroxidase, catalase and superoxide dismutase, respectively.
In the same way, the content of K had been negative and non-significantly correlated with N % by (-0.005) and peroxidase by (-0.474). In addition to that mentioned before, trait of P accumulation had negative and non-significant correlation with both total chlorophyll, chlorophyll a, chlorophyll b, carotenoids, ascorpate peroxidase, catalase and superoxide dismutase with the folowing values: (-0.442), (-0.458), (-0.404), (-0.436), (-0.215), (-0.202), and (-0.432), respectively.
Based on this, the only negative correlation coefficient between the element N % and both peroxidase (-0.184), ascorpate peroxidase (-0.203), catalase (-0.357) and superoxide dismutase (-0.152) that were not significant was either at 5 or 1% level of probability. Also, the relation between proline and both chlorophyll b, ascorpate peroxidase and catalase gave value of (0.568), (-0.473) and (-0.441), respectively, making the negative and non-significant relations at either 5 or 1% probability level represent more than 33% of the data.
Al-Saady et al. (2012) reported that the existence of NaCl in soil had a massive impact on the content of proline and chlorophyll. According to Yunus (2019), correlation analysis revealed that traits under stress condition had no significant correlation with the traits under non-stress environment, indicating high stress intensity and adds that the drought tolerant genotypes identified in this research could be utilized in future breeding program for further improvement in curly pepper. Many investigators had studied the importance of correlation (El-Ghany et al., 2012; Fufa, 2013; Ichwan et al., 2017; Khan, 1985; Kole and Saha, 2013; Nemeskéri and Helyes, 2019; Sadek et al., 2006; Singh and Chowdhury, 1983; Waitt and Levin, 1998). The role and impact of proline, vitamins and enzymes and its relation with others under drought and salt stress conditions had been studied by many researchers (Abdelaal et al., 2020; Al-Saady et al., 2012; Beltagi, 2008; Gadalla, 2009; Misra and Gupta, 2005; Oertli, 1987; Ratnakar and Rai, 2013; Sun et al., 2011; Veeranagamallaiah et al., 2007).
CONCLUSION
Considering all the parameters of the preliminary study, the preceding findings as a whole and with respect to the sweet pepper hybrid variety tested, it can be concluded that there are various physiological alterations caused by water deficit proceedings for all treatments studied except one (N constant). The findings suggest that irrigation procedures (in pots) with 20% moisture stress to attain pot capacity (W2) are more effective than 40% (W3) and 60% (W4) from the viewpoint of more effective water usage. This would help to reduce drought damage, maintain healthy plants in order to ensure superior performance, and ability to adapt under water deficit and save water use by 20%.
CONCLUSION
The author has not declared any conflict of interests.
REFERENCES
|
Abdel-Ghany H, Nawar A, Ibrahim M, El-Shamarka A, Selim M, Fahmi A (2004). Using tissue culture to select for drought tolerance in bread wheat. Paper presented at the Proceedings of the 4th International Crop Science Congress Brisbane, Australia. |
|
|
Abdelaal KA, EL-Maghraby LM, Elansary H, Hafez YM, Ibrahim EI, El-Banna M, Elkelish A (2020). Treatment of sweet pepper with stress tolerance-inducing compounds alleviates salinity stress oxidative damage by mediating the physio-biochemical activities and antioxidant systems. Agronomy 10(1):26. |
|
|
Aebi H (1974). Catalase. In Methods of Enzymatic Analysis: Elsevier pp. 673-684 |
|
|
Ahanger MA, Akram NA, Ashraf M, Alyemeni MN, Wijaya L, Ahmad P (2016). Signal transduction and biotechnology in response to environmental stresses. Biologia Plantarum 61(3):401-416. |
|
|
Al-Saady N, Khan A, Rajesh L, Esechie H (2012). Effect of salt stress on germination, proline metabolism and chlorophyll content of fenugreek (Trigonella foenum gracium L.). Journal of Plant Sciences 7(5):176. |
|
|
Alaei Y, Khanghah AM, Jafari M, Khaneghah A (2012). Evaluation on leaf proline amount in three bread wheat cultivars in presence of two fertilizers containing amino acids in drought stress. World Applied Sciences Journal 18(9):1190-1192. |
|
|
Almohisen I (2015). Effect of Water Stress on Growth and Physiology of Tomato (Lycopersicon esculentum Mill.). Hortscience Journal of Suez Canal University 4(1):1-5. |
|
|
Anjum SA, Saleem MF, Wang L, Bilal MF, Saeed A (2012). Protective role of glycinebetaine in maize against drought-induced lipid peroxidation by enhancing capacity of antioxidative system. Australian Journal of Crop Science 6(4):576. |
|
|
Arnon DI (1949). Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant physiology 24(1): 1-15. |
|
|
Azam M, Norman M, Abbasi N, Ramzan A, Imran M, Hayat A, Akram A (2016). Effect of foliar application of micro-nutrient and soil condition on growth and yield of sweet pepper (Capsicum annuum L.). Science, Technology and Development 35:75-81. |
|
|
Bates LS, Waldren RP, Teare ID (1973). Rapid determination of free proline for water-stress studies. Plant and Soil 39(1):205-207. |
|
|
Beltagi MS (2008). Exogenous ascorbic acid (vitamin C) induced anabolic changes for salt tolerance in chick pea (Cicer arietinum L.) plants. African Journal of Plant Science 2(10):118-123. |
|
|
Beyer WF, Fridovich I (1987). Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Analytical Biochemistry 161(2):559-566. |
|
|
Buschmann PH, Vaidyanathan R, Gassmann W, Schroeder JI (2000). Enhancement of Na+ uptake currents, time-dependent inward-rectifying K+ channel currents, and K+ channel transcripts by K+ starvation in wheat root cells. Plant physiology 122(4):1387-1398. |
|
|
Chaves MM (1991). Effects of Water Deficits on Carbon Assimilation. Journal of Experimental Botany 42(1):1-16. |
|
|
Cornic G, Massacci A (1996). Leaf photosynthesis under drought stress. In Photosynthesis and the Environment Springer pp. 347-366. |
|
|
Delfine S, Loreto, F, Alvino A (2001). Drought-stress effects on physiology, growth and biomass production of rainfed and irrigated bell pepper plants in the Mediterranean region. Journal of the American Society for Horticultural Science, 126(3):297-304. |
|
|
Doorenbos J. (1975). Guidelines for predicting crop water requirements. Food and Agriculture organization. Rome, Irrig. Drainage pap., 24. |
|
|
El-Ghany H, El-Salam MA, Hozyen M, Afifi M (2012). Effect of deficit irrigation on some growth stages of wheat. Journal of Applied Sciences Research pp. 2776-2784. |
|
|
El-Saidy AEA, Farouk S, El-Ghany HMA (2011). Evaluation of Different Seed Priming on Seedling Growth, Yield and Quality Components in Two Sunflower (Helianthus annuus L.) Cultivars. Trends in Applied Sciences Research 6(9):977-991. |
|
|
El Sayed H (1992). Proline metabolism during water stress in sweet pepper (Capsicum annuum L.) plant. Paper Presented at the Phyton: annales rei botanicae. |
|
|
Fufa M (2013). Correlation studies on yield and yield components of fenugreek (Trigonella Foenum-Graecum L.) lines evaluated in South-Eastern Ethiopia. Wudpecker Journal of Agricultural Research 2:280-282. |
|
|
Gadalla S (2009). The roles of ascorbic acid α-tocopherol in minimize of salt-induced whert leaf senescence. Journal of Plant Production 34(11):10645-10661. |
|
|
Gaveh EA, Timpo GM, Agodzo SK, Shin DH (2011). Effect of irrigation, transplant age and season on growth, yield and irrigation water use efficiency of the African eggplant. Horticulture, Environment, and Biotechnology 52(1):13-28. |
|
|
Ichwan B, Suwignyo RA, Hayati R (2017). Response of red chilli varieties under drought stress. Russian Journal of Agricultural and Socio-Economic Sciences 66(6). |
|
|
Karam F, Saliba R, Skaf S, Breidy J, Rouphael Y, Balendonck J (2011). Yield and water use of eggplants (Solanum melongena L.) under full and deficit irrigation regimes. Agricultural Water Management 98(8):1307-1316. |
|
|
Khan IA (1985). Correlation and path coefficient analysis of yield components in mung bean (Phaseolus aureus Roxb.). Botanical Bulletin of Academia Sinica 26:13-20. |
|
|
Kirnak H, Kaya C, Tas I, Higgs D (2001). The influence of water deficit on vegetative growth, physiology, fruit yield and quality in eggplants. Bulgarian Journal of Plant Physiology 27(3-4):34-46. |
|
|
Kole P, Saha A (2013). Correlation coefficients of component characters with seed yield and their direct effects in path analysis in fenugreek grown under six environments. Journal of Horticulture and Forestry 5(1):17-20. |
|
|
Kurniawati S (2014). Drought Stress Tolerance Mechanisms of Eggplant (Solanum spp.): Physiology and Molecular. |
|
|
Liu YJ, Tong YP, Zhu YG, Ding H, Smith EA (2006). Leaf Chlorophyll Readings as an Indicator for Spinach Yield and Nutritional Quality with Different Nitrogen Fertilizer Applications. Journal of Plant Nutrition 29(7):1207-1217. |
|
|
Misra N, Gupta A K (2005). Effect of salt stress on proline metabolism in two high yielding genotypes of green gram. Plant Science 169(2):331-339. |
|
|
Molden D (1997). Accounting for water use and productivity, SWIM Paper 1, system-wide initiative for water management. International Water Management Institute, Colombo, Sri Lanka, 16 p. |
|
|
Molden D, Sakthivadivel R, Habib Z (2001). Basin-level use and productivity of water: Examples from South Asia (Vol. 49): IWMI. |
|
|
Nemeskéri E, Helyes L (2019). Physiological Responses of Selected Vegetable Crop Species to Water Stress. Agronomy 9(8):447. |
|
|
Nissen O, Eisensmith S, Freed R, Everson E, Smail V, Weber M, Portice G (1985). A microcomputer program for the design, management and analysis research experiments. In: Version. |
|
|
Oertli J (1987). Exogenous application of vitamins as regulators for growth and development of plants-a review. Zeitschrift für Pflanzenernährung und Bodenkunde 150(6):375-391. |
|
|
Okunlola GO, Olatunji OA, Akinwale RO, Tariq A, Adelusi AA (2017). Physiological response of the three most cultivated pepper species (Capsicum spp.) in Africa to drought stress imposed at three stages of growth and development. Scientia Horticulturae 224:198-205. |
|
|
Ratnakar A, Rai A (2013). Influence of NaCl salinity on β-carotene, thiamine, riboflavin and ascorbic acid contents in the leaves of Atriplex hortensis L. var. Pusa Bathua No. 1. Journal of Stress Physiology and Biochemistry 9(4):187-192. |
|
|
Richards LA (1969). Diagnosis and improvement of saline and alkali soils: United States Department Of Agriculture; Washington. |
|
|
Sadek S, Ahmed M, Abd El-Ghaney H (2006). Correlation and path coefficient analysis in five parents inbred lines and their six white maize (Zea mays L.) single crosses developed and grown in Egypt. ournal of Applied Sciences Research 2(3):159-167. |
|
|
Sakharov IY, Ardila GB (1999). Variations of peroxidase activity in cocoa (Theobroma cacao L.) beans during their ripening, fermentation and drying. Food Chemistry 65(1):51-54. |
|
|
Sayyari M, Ghanbari F (2012). Effects of super absorbent polymer A200 on the growth, yield and some physiological responses in sweet pepper (Capsicum annuum L.) under various irrigation regimes. International Journal of Agricultural and Food Research 1(1). |
|
|
Showemimo F, Olarewaju J (2007). Drought tolerance indices in sweet pepper (Capsicum annuum L.). International Journal of Plant Breeding and Genetics 1(1):29-33. |
|
|
Singh B, Chowdhury R (1983). Correlation and path coefficient analysis of seed yield and oil content in mustard (Brassica juncea). Canadian Journal of Genetics and Cytology 25(3):312-317. |
|
|
Snedecor G, Cochran W (1980). Statistical methods. Iowa State University Press, Ames pp.358-360. |
|
|
Sun J, Li T, Xia J, Tian J, Lu Z, Wang R (2011). Influence of salt stress on ecophysiological parameters of Periploca sepium Bunge. Plant, Soil and Environment 57(4):139-144. |
|
|
Tahi H, Wahbi S, El Modafar C, Aganchich A, Serraj R (2008). Changes in antioxidant activities and phenol content in tomato plants subjected to partial root drying and regulated deficit irrigation. Plant Biosystems - An International Journal Dealing with all Aspects of Plant Biology 142(3):550-562. |
|
|
Veeranagamallaiah G, Chandraobulreddy P, Jyothsnakumari G, Sudhakar C (2007). Glutamine synthetase expression and pyrroline-5-carboxylate reductase activity influence proline accumulation in two cultivars of foxtail millet (Setaria italica L.) with differential salt sensitivity. Environmental and Experimental Botany 60(2):239-244. |
|
|
Waitt DE, Levin DA (1998). Genetic and phenotypic correlations in plants: a botanical test of Cheverud's conjecture. Heredity 80(3):310-319. |
|
|
Wang SY, Jiao M, Faust M (1991). Changes in ascorbate, glutathione, and related enzyme activities during thidiazuron-induced bud break of apple. Physiologia Plantarum 82(2):231-236. |
|
|
Yunus A (2019). Screening of capsicum annuum l. Genotypes for drought tolerance based on drought tolerance indices. SABRAO Journal of Breeding and Genetics 51(3). |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


