Full Length Research Paper

ABSTRACT
This study aims at evaluating the agronomic efficiency of the Bacillus subtilis UFMT-Pant001 strain as a plant growth promoter in soybean at three greenhouses from distinct Brazilian regions. The treatments were formulated based on this strain at a concentration of 1x109 UFC.mL-1 in volumes of 100, 150, 200, 250, 300, 350 or 400 mL for each 50 kg of seeds. Bradyrhizobium japonicum was added in all experiments, and the absolute control contained only this bacterium. The positive control employs a commercial product based on Bacillus subtilis. Both inoculums were adjusted at the recommended dose of 100 mL/50 kg of seeds. In the experiments, the soybean cultivars 7110 RR, Desafio RR, TMG-132 and CG 7665 RR were used. A positive influence on the increment in plant height, biomass, number of internodes, nodules, grains, pods, roots, nutrients and shoot dry mass per plant inoculated with UFMT-Pant001 strain was observed. Although the results are promising to the B. subtilis UFMT-Pant001 strain as a potential plant growth promoter for soybean, more tests, including with other cultivars, are needed, especially in the field.
Key words: Bacillus subtilis, bacterial-based inoculant, soybean, plant growth-promoting rhizobacteria.
INTRODUCTION
Soybean (Glycine max (L.) Merrill) is the main oilseed produced globally. Brazil is the largest soybean exporter and the second largest producer in the world (CONAB, 2020), broadly distributing soybean and producing it in almost all Brazilian states. Important states to soybean production include the states of Mato Grosso, Paraná, Rio Grande do Sul, Mato Grosso do Sul, Goiás and Tocantins, reflecting its national importance.
Soybean is of great importance as a food source, but still there is a need for studies and research on approaches to increase its production, including genetic improvements to the cultivars, soil and crop management, seed quality, and increased use of beneficial micro-organisms (increase the availability of mineral nutrients, the production of growth hormones such as gibberellins and auxins, and the suppression of deleterious micro-organisms), among other efforts that are involved in crop development. In recent years, interest in the use of micro-organisms in agricultural practices has increased significantly. Microorganisms are potential substitutes for chemical products to promotion of plant growth and the biological control of pests and plant diseases, among other applications that potentially favoring reduced costs and preserving the environment (Braga et al., 2018; Dias et al., 2019; Chagas Junior et al., 2021).
These micro-organisms, known as plant growth-promoting rhizobacteria (PGPR), provide several benefits to plants, such as increases in seed germination rates, root growth, grain number, leaf growth, leaf area, nutrient content, and drought tolerance and delays in leaf senescence, and in addition, they also can serve as biocontrol of diseases through direct action against a pathogen and indirect action that makes a plant more resistant (Melo, 2015; Contreras-Cornejo et al., 2016; Zeilinger et al., 2016; Lima et al., 2019).
Among these micro-organisms, the bacterium Bacillus subtilis is a natural inhabitant of the soil, produces enzymes and phyto-hormones that provide benefits to plants, and is described as a promoter of plant growth. It produces the antibiotics bulbiformin, mycosubtilin, bacillomycin, bacilizin, and fungicidin and can also inhibit phytopathogens (Saravanakumar et al., 2016; Shafi et al., 2017).
Micro-organisms can confer protection to plants, either by their presence in host plants or by their application as biocontrol agents, which can result in the elimination of agricultural pests. Therefore, there has been growing interest in studying the occurrence, the potential for colonization, and the use of endophytic bacteria and other endophytes to promote plant growth and the biological control of plant diseases. Thus, the objective of the present study was to evaluate the efficacy of the bacterial-based inoculant of the B. subtilis strain UFMT-Pant001 as a plant growth promoter in soybean plants in a greenhouse.
MATERIALS AND METHODS
Location of experiments
Three independent experiments were conducted in a greenhouse installed in the following regions: Experiment 1 at the School of Engineering of Ilha Solteira at São Paulo State University (UNESP) in Ilha Solteira, SP; Experiment 2 at the School of Agronomy and Animal Science (UFMT) in Cuiabá, MT; and Experiment 3 at the State University of Mato Grosso (UNEMAT), Campus II, in the municipality of Alta Floresta, MT.
Bacillus subtilis UFMT-Pant001-Identification
The B. subtilis strain UFMT-Pant001 was identified as described by Machado et al. (2010). The strain was grown and sub-cultured in Luria-Bertani (LB) medium, and maintained in the microbial collection of the Laboratory of Microbiology, School of Medicine, Federal University of Mato Grosso.
From previously observational studies with different Bacillus isolates, the strain UFMT-Pant001 was screened as able to promote a better growth of soybean plants in greenhouse (unpublished data).
Experiment 1
Two independent experiments were conducted in a greenhouse located on Campus II of the School of Engineering of Ilha Solteira (UNESP), in the agricultural year 2017. The soil of the experimental area is classified as Eutrophic Red Argisol (Embrapa, 2011). Fertilization was performed in pots with 2 g of formula 8-28-16 per pot.
The experiments were conducted in a completely randomized design containing six treatments. Thus, four doses of B. subtilis UFMT-Pant001 were used (100, 200, 300 and 400 mL for 50 kg of seed), one treatment with a commercial product (B. subtilis with 100 mL of bw/50 kg of seed, positive control) and an absolute control treatment without B. subtilis inoculation, with 4 replicates. The inoculation was performed with the product containing B. subtilis UFMT-Pant001 at the time of planting. All treatments were inoculated with a Bradyrhizobium japonicum-based product at a dose of 100 mL per 50 kg of seeds, as recommended by the manufacturer.
The cultivars used were 7110 RR and Desafio RR. Sowing was performed in 15-L pots, in which six seeds were sown per pot at a depth of 2 cm and covered with 2 cm of soil, and at fifteen days after emergence, thinning was performed, leaving two plants. Two evaluations were performed during the crop cycle. The first was performed 45 days after sowing (DAS), and the second was performed at harvest.
In the first evaluation, the root system was separated from the aerial part of the plants, and the roots were properly washed in running water and then measured with a ruler graduated in mm. The roots and shoots were separately stored in labeled paper bags, which were placed in a forced air oven at a temperature of 65°C to constant weight.
Then, the material was weighed on a precision scale (0.001 g), and the mean values obtained were used to determine the root dry mass (RDM) and shoot dry mass (SDM) of the soybean plants in g plant-1. The dry mass (DM) was calculated using the data obtained for the SDM and SDM. With the biomass data, the relative efficiency (RE) of each treatment was determined and calculated according to the formula RE = (SDM inoculated with the isolates/SDM without inoculant) x 100.
The second evaluation was performed at the soybean harvest. The production components were determined: number of internodes and number of pods per plant, which were obtain using the ratio between the total number of pods obtained and the number of plants evaluated; number of grains per plant, which was obtained using the ratio between the total number of seeds obtained and the number of plants evaluated; mass of 100 grains, which was obtained by weighing 100 grains on a precision scale; and grain weight per plant.
Using R software (R Development Core Team, 2015), the data were subjected to Levene’s normality and homoscedasticity tests, both at a significance level of 5%. Subsequently, the normal data were subjected to analysis of variance by the F test, and when significant, the means were analyzed by the Duncan test at a significance level of 5%.
Experiment 2
The experiment was set up and conducted in a greenhouse at the School of Agronomy and Animal Science (FAAZ) in the municipality of Cuiabá in the agricultural year 2016. The soil was chemically corrected using 90 kg.ha-1 of P2O5 and 80 kg.ha-1 of K2O. The potassium fertilization was divided into two applications, with intervals of 15 days.
The experiments were conducted in a completely randomized design using 3 kg pots composed of three doses of B. subtilis UFMT-Pant001 (150, 250 and 350 mL for 50 kg of seed) and an absolute control without inoculation with B. subtilis, with 4 replicates. All treatments were inoculated with a B. japonicum-based product at a dose of 100 mL per 50 kg of seeds, as recommended by the manufacturer.
Five seeds were sown per pot, and ten DAS, only three plants were left per pot. Crop management and the application of pesticides (acaricides, insecticides, and fungicides) were performed according to the technology adopted for the soybean crop (Henning, 2009). For the control of some pests, Fastac Duo® was used, following the manufacturer's dose recommendations. Plant diseases were not detected.
The pots were disassembled, and the nodules were removed from the root, washed, dried on paper towels, and weighed. Similar to the process for the root system, the shoots of the plants were also dried at 65°C to constant weight.
The results were subjected to ANOVA, and when the “F” test was significant, the means of the treatments were compared by the Tukey test at the 5% significance level.
Experiment 3
The experiment was conducted at the Mato Grosso State University (UNEMAT), on Campus II, located in the municipality of Alta Floresta, northern region of the state of Mato Grosso, in the 2016/17 harvest.
The culture was performed in pots with a capacity of 8 dm-3. Soil liming was performed 30 DAS, with 95% relative total neutralization power (RTNP) dolomitic limestone, aiming to increase the soil base saturation to 60%. Base fertilization, which was 200 mg.dm-3 of P and 150 mg.dm-3 of K, using simple superphosphate (18% P2O5) and potassium chloride (60% K2O) as sources, respectively, was performed before sowing.
Six seeds were sown, and then, thinning occurred, leaving two plants per pot. Overall, with three evaluations, the experiment consisted of 72 pots.
The products used to inoculate the seeds were fungicide + inoculant + coinoculant, which were applied in the appropriate order to ensure greater efficiency and better development of the symbiotic organisms present.
The fungicide contained carboxin + tiram, each at a concentration of 200 g L-1, at a dose of 200 mL of commercial product for each 100 kg of seeds. On the day following the application of the fungicide, the seeds were inoculated with a solid peat inoculant for soybean, which contained the bacterium B. japonicum, at a dose of 80 g for 50 kg of seed. Then, the bacteria were inoculated with B. subtilis UFMT-Pant001. Six treatments were conducted, one with each of four different doses of B. subtilis UFMT-Pant001 (100, 200, 300, and 400 mL for each 50 kg of seed), a positive control with the commercial product (B. subtilis with 100 mL of bw/50 kg of seed), and an absolute control without B. subtilis inoculation. The experimental design was completely randomized, with 6 treatments and four replicates.
The soybean cultivar CG 7765 RR was used. Weeds that emerged in the pots were manually controlled. Systemic and contact insecticides were used to control pests harmful to the crop, such as bugs, beetles, whiteflies and caterpillars. The applications were performed across the entire area to ensure all unwanted pests in the crop were addressed. To control diseases, systemic and contact fungicides were used and applied for prevention and control purposes.
Three evaluations were performed in the following crop phases: the first evaluation at 20 to 25 DAS (Stage V3 to V5), the second evaluation at 40 to 45 DAS (Stage R1), and the third evaluation at the end of the cycle (Harvest).
The characteristics analyzed were SDM, RDM, total DM (TDM), shoot and root length, and number and DM of nodules, which were dried in an oven at 65 °C to constant weight and then weighed on an analytical balance. The nitrogen and phosphorus levels were measured in the laboratory after the DM of the plant was ground in a Willer mill and placed in a paper bag (Malavolta et al., 1997). The percentages of N and P (relative to the negative control), the N and P use efficiency (ratio between the DM and the N and P levels), and the RE were determined by the ratio between the SDM of the treatment with B. subtilis and the SDM of the treatment without B. subtilis (absolute control). The production components evaluated were the number of pods per plant, 100-grain weight, and grain yield converted into kg ha-1 at 13% moisture.
According to the data obtained, the results were statistically analyzed by a simple ANOVA, and the estimates were compared by the F test. The means of the treatments were compared by the Duncan test at the 5% significance level with assistance of software.
RESULTS
Experiment 1
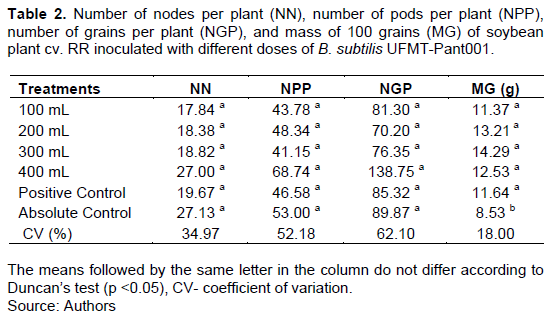
For cultivar 7110 RR, the SDM, TDM, and RE were higher for the treatment with the 400-mL B. subtilis dose than for the other treatments (Table 1). There was no significant difference in plant height (PH), root length, or RDM (Table 1).There was only a significant difference for the 100-grain weight, which was higher in the treatments with the different doses of B. subtilis UFMT-Pant001 and the positive control than in the absolute control (Table 2).
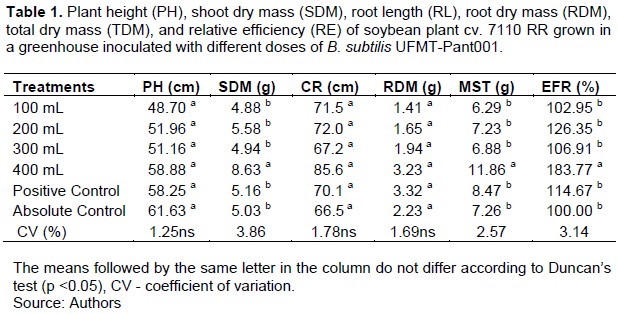
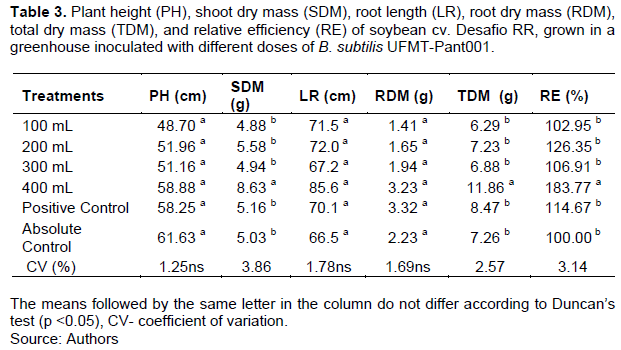
For the cultivar Desafio RR, the SDM of the plants in the first evaluation was influenced by the treatments used, where in comparison to the absolute control, the doses of 200 and 400 mL of B. subtilis and the positive control had higher means (Table 3). Similarly, they also had higher REs than that of the absolute control (Table 3).
Experiment 2
The pH, fresh mass and SDM were all higher in the treatments with the four different doses of B. subtilis UFMT-Pant001 than in the absolute control treatment without inoculation (Table 1). The RDM was higher in the treatment with the 150 mL dose than in the absolute control (Table 4).
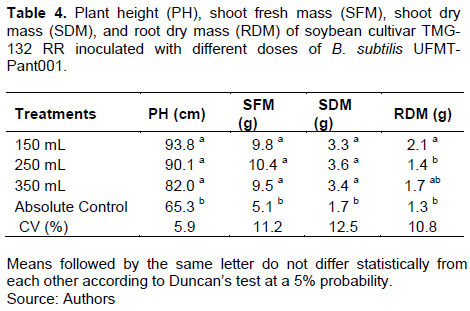
The nodule number (NN) was significantly higher in the treatment with the 350 mL dose than in the absolute control (Table 5). The DMs of the nodules in the 150 and 350 mL treatments were significantly higher than those in the absolute control. The number of pods in the 150 mL treatment was higher than that in the absolute control. The pod mass in all treatments with the different doses were higher than those in the absolute control (Table 5).
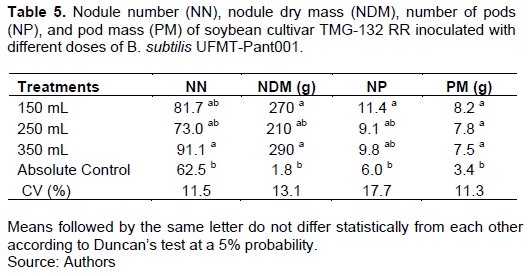
Experiment 3
Of the treatments, the positive control resulted in the longest root length, and this value differed only from that in the treatment with the 100 mL dose of B. subtilis (Table 6). Although there was no significant differences between the root lengths in the treatments with the 300 mL and 400 mL doses and the positive control in comparison to that in the absolute control, these three treatments had root lengths 6.50, 1.85 and 11.86% greater than the absolute control, respectively, indicating the benefit of applying the bacteria at these doses.
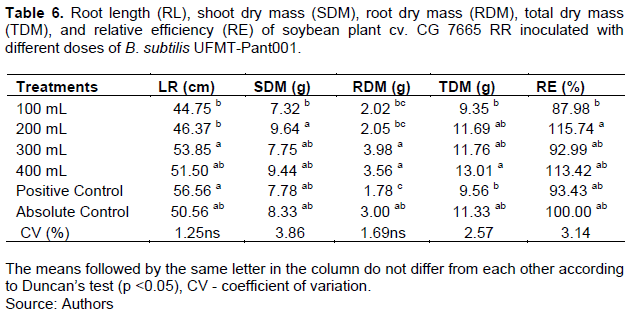
The SDM was higher in the treatment with the 200 mL dose of B. subtilis than in the other treatments; however, this value differed only from that in the treatment with the 100 mL does of B. subtilis (Table 6). The treatments with the 300 and 400 mL of B. subtilis produced the highest RDM, which was higher than those in the treatments with the 200 mL does and the positive control (Table 6). Of the treatments, the 400 mL dose of B. subtilis had the highest mean TDM, which was higher than that in the treatment with the 100 mL dose and the positive control (Table 6). The highest RE in the first evaluation was observed in the treatment with the 200 mL dose of B. subtilis, and this RE value was higher only than that in the treatment with the 100 mL does of B. subtilis (Table 6). Notably, the treatment with the 200 mL does of B. subtilis had a RE value 15.74% above that of the absolute control.
In the second evaluation, of the treatments, the treatment with the 100 mL dose of B. subtilis had the highest root length, SDM, TDM, and RE, and the root length was higher only in relation to that in the treatment with the 400 mL dose of B. subtilis. Additionally, the SDM, TDM, and RE did not differ, except from those in the 200 mL dose treatment. The absolute control treatment had the highest mean RDM, but this value differed only from that in the positive control (Table 7).
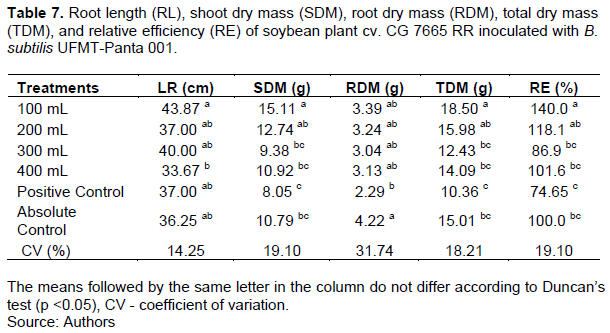
There was no difference between the treatments in terms of the NN (Table 8). Of the treatments, the absolute control had the highest DM of the nodules, differed only from that in the treatment with the 400 mL dose of B. subtilis (Table 8).
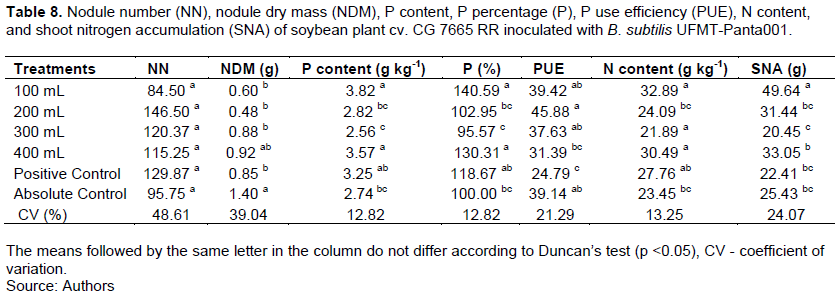
The highest levels and percentage of P were observed for the treatment with the 100 mL dose of B. subtilis, and these values only differed from that in the treatment with the 100 mL dose and the positive control (Table 8). The treatment with the 200 mL dose of B. subtilis showed the highest efficiency of P use, which differed from those in the treatments with the 400 mL dose and the positive control (Table 8). Of the treatments, the treatment with the 100 mL dose had the highest means for the N content and nitrogen accumulation in the shoots. In addition, the N content differed from that in the treatment with the 200 mL dose and the absolute control, and the nitrogen accumulation in the shoots differed from those in all the other treatments (Table 8).
Of the treatments, the treatment with the 200 mL dose had the highest pH, which was higher than that in the treatment with the 400 mL dose and the absolute control (Table 9). The treatment with the 300 mL dose had the highest number of internodes, and this value did not differ from those in the 400 mL dose treatment and the positive control (Table 9).
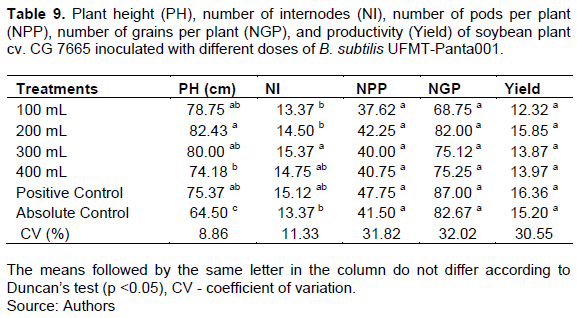
Regarding the number of pods per plant, number of grains per plant, and plant yield, there was no difference between treatments (Table 9).
DISCUSSION
The results show a greater increase in biomass (SDM, RDM, and TDM), nodulation (NN, NDM, and RE, nutrients in the shoots (N content and SNA), number of internodes, number of pods, and productivity of different soybean cultivars in the different experiments in a greenhouse when inoculated with B. subtilis UFMT-Pant001 than in the controls. These increase may be linked to the various mechanisms by which this bacterium acts, such as the production of hydrocyanic acid, phyto-hormones, and enzymes, and the availability of nutrients (P and N), and it may also act in the biological control of phytopathogens, among others.
The increase may be related to the ability of rhizo-bacteria to produce plant growth regulators (RGRs), which are organic substances that influence the physiological processes of plants at low concentrations (Saeid et al., 2018; Saxena et al., 2019), improve plant growth and yield, facilitate the absorption of nutrient minerals, synthesize various phytohormones, and protect plants from various phytopathogens (Kalam et al., 2017; Diaz et al., 2019). The genus Bacillus represents one of the most diverse genera of bacilli commonly used as bio-inoculants to promote plant growth. Akinrinlola et al. (2018) identified 12 strains of Bacillus that promote the growth of corn, wheat and soybean. The strains exhibited multiple characteristics of plant growth promoters, including phosphate solubilization, nitrogen fixation, and indole acetic acid (IAA) production (Contreras-Cornejo et al., 2016).
According to Mohamed et al. (2018), B. subtilis isolated from the rhizosphere of tomato plants was able to solubilize phosphate in a Pikovskaya medium and was characterized by a high phosphate solubilization index, increased ability to release mineral phosphate with decreasing pH values, and resistance to commonly used pesticides, all of which are important characteristics of bacterial isolates suitable for plant growth.
The rhizobacteria of the genus Bacillus are capable of producing gibberellin and auxin (Saxena et al., 2019). Cerqueira et al. (2015) used four isolates of Bacillus spp. in in vitro tests and confirmed the production of IAA, acetylene reduction activity (ARA), and 1-aminocyclopropane-1-carboxylate (ACC)-deaminase by these isolates. Saharan and Nehra (2011) observed that Bacillus species contributed to the improvement of different root parameters, such as rooting, root length and dry matter content, and that inoculation with IAA-producing isolates increased the absorption of some nutrients, promoting sweet potato growth and greater rooting of eucalyptus seedlings.
Kalan et al. (2020) found a significant increase in tomato colonization after inoculation with different Bacillus isolates, and greenhouse experiments with these isolates indicated a general increase in tomato plant growth over 60 days, with TRS-7 and TRS-8 being the best plant growth promoters among the seven isolates tested, with potential use as inoculants to increase tomato productivity.
One way in which the tested B. subtilis UFMT-Pant001 product, inoculated in the different soybean cultivars, may have contributed to the increase in biomass was its positive effect on the availability and solubilization of nutrients such as phosphorus and nitrogen. Araujo et al. (2012) reported that in their study using the cowpea cultivar BRS Guariba, simple inoculation with B. subtilis (PRBS-1) provided the greatest increase in plant growth and N fixation and did not affect nodulation at 40 and 55 DAS.
According to Fonseca et al. (2022), inoculation with B. subtilis in sugarcane improved plant nutrition and chlorophyll concentration, as well as gas exchange parameters (especially liquid photosynthetic rate and water use efficiency), even under drought conditions. In addition, the stress parameters (antioxidant metabolism activity) were reduced in the inoculated plants. These benefits resulted in increased root growth, tillering, stem weight, and sucrose concentration in the stems.
In comparison to the positive and absolute control treatments, the treatments with different doses result in higher NN and DMN for some soybean cultivars in some experiments (p <0.01), showing these positive effects as a function of the coinoculation of B. subtilis UFMT-Panta001 and B. japonicum in soybean.
Araujo and Guerreiro (2010), in an experiment with cowpea that evaluated the coinoculation of B. subtilis and Bradyrhizobium, showed that in the inoculated treatments, there was an increase in cowpea nodulation with coinoculation, suggesting an influence of coinoculation with B. subtilis and Bradyrhizobium in the promotion of nodulation. Possibly, the metabolites produced by B. subtilis UFMT-Panta001 are not toxic to rhizobia, and coinoculation with both bacteria may increase nodulation, contribute to the increases in the competitiveness of the inoculated bacteria and in the infection sites, and inhibit the growth of pathogenic fungi in the roots, as reported by other authors (Mardanova et al., 2017; Saxena et al., 2019; Costa et al., 2022).
Thus, the increase in the main characteristics evaluated in the different experiments in the greenhouse on the soybean crop inoculated with B. subtilis UFMT-Pant001 can be explained by the probable synthesis or stimulus of phytohormone production, as well as by phosphate solubilization, both of which act on the unavailable P in the soil and the P supplemented by fertilization. The use of beneficial and environmentally safe microbial inoculants is an alternative to using synthetic agrochemicals. Thus, the B. subtilis UFMT-Pant001 product tested in this study showed potential for promoting the growth of soybean plants.
CONCLUSIONS
In the different experiments in the greenhouse, in comparison to the absolute control, the B. subtilis UFMT-Pant001 strain, which was used as an inoculant, increased soybean biomass? In terms of the different doses of B. subtilis UFMT-Pant001, doses of 200 to 400 mL for 50 kg of seeds at a concentration of 1x109 UFC.mL-1 had positive effects on soybean biomass and on similar other evaluated characteristics. The use of B. subtilis UFMT-Pant0001 also contributed to the increase in the number and weight of nodules.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Akinrinlola RJ, Yuen GY, Drijber RA, Adesemoye AO (2018). Evaluation of Bacillus strains for plant growth promotion and predictability of efficacy by in vitro physiological traits. International Journal of Microbiology 2018:e20190642. |
|
|
Araujo FF de, Bragante RJ, Bragante CE (2012). Controle genético, químico e biológico de meloidoginose na cultura da soja. Pesquisa Agropecuária Tropical, 42(2):220-224. |
|
|
Braga GM, Chagas LFB, Amaral LRO, Miller LO, Chagas AF (2018) Efficiency of inoculation by Bacillus subtilis on soybean biomass and productivity. Revista Brasileira de Ciências Agrárias 13(4):e5571. |
|
|
Cerqueira WF, MJS, MJS, MIKS, SAFJ (2015). Influência de bacté-rias do gênero Bacillus sobre o crescimento de feijão comum (Phaseolus vulgaris l.). Enciclopédia Biosfera 11:82-93. |
|
|
Chagas Junior AF, Chagas LFB, Martins ALL, Colonia BSO, Souza MC, Braga Junior GM (2021). Efficiency of Bacillus subtilis bs10 as a plant growth promoting inoculant in soybean crop under field conditions. Research, Society and Development 10(14):e441101422141. |
|
|
CONAB- National Supply Company (2020). Acompanhamento da safra brasileira de grãos (Safra 2019/20). 7:1-68. |
|
|
Contreras-Cornejo HA, Macías-Rodríguez L, del-Val E, Larsen J (2016). Ecological functions of Trichoderma spp. and their secondary metabolites in the rhizosphere: interactions with plants. FEMS Microbiology Ecology 92(4):fiw036. |
|
|
Costa N de JF, Santos e Silva MSB dos, Candido e Silva EK, Oliveira ACS de, Rodrigues AAC (2022). Tratamento térmico e biológico de sementes de alface no controle de fungos fitopatogênicos. Diversitas Journal 7(2):596-606. |
|
|
de Araujo FF, Guerreiro RT (2010). Bioprospection of Bacillus isolates promoters of corn growth in natural and sterile soil. Ciencia e Agrotecnologia 34(4):837-844. |
|
|
Diaz PAE, Baron NC, Rigobelo EC (2019). Bacillus spp. as plant growth-promoting bacteria in cotton under greenhouse conditions. Australian Journal of Crop Science 13(12):2003-2014. |
|
|
Empresa Brasileira de Pesquisa Agropecuária (EMBRAPA) (2011). Manual de Métodos de análise de solo. EMBRAPA-CNPS, Ed., Brazilian Government, Brasilia, DF, Brazil. |
|
|
Fonseca M de C da, Bossolani JW, de Oliveira SL, Moretti LG, Portugal JR, Scudeletti D, de Oliveira EF, Crusciol CAC (2022). Bacillus subtilis inoculation improves nutrient uptake and physiological activity in sugarcane under drought stress. Microorganisms 10(4):809. |
|
|
Henning AA (2009). Manejo de doenças da soja (Glycine max L. Merrill). Informativo ABRATES 19:9-12. |
|
|
Kalam S, Basu A, Podile AR (2020). Functional and molecular characterization of plant growth promoting Bacillus isolates from tomato rhizosphere. Heliyon 6(8):e04734. |
|
|
Kalam S, Das SN, Basu A, Podile AR (2017). Population densities of indigenous acidobacteria change in the presence of plant growth promoting rhizobacteria (PGPR) in rhizosphere. Journal of Basic Microbiology 57(5):376-385. |
|
|
Lima UA, AE, BW, SW (2019). Biotecnologia industrial - processos fermentativos e enzimáticos. Editora Blucher 3:616. |
|
|
Machado AP, Vivi KV, Tavares JR, Gueiros Filho FJ, Fischman O (2010). Antibiosis and dark-pigments secretion by the phytopathogenic and environmental fungal species after interaction in vitro with a Bacillus subtilis isolate. Brazilian Archives of Biology and Technology 53(5):997-1004. |
|
|
Malavolta E, Vitti GC, Oliveira SA (1997). Avaliação do estado nutricional das plantas: princípios e aplicações. Piracicaba: Potafós: 319 p. |
|
|
Mardanova AM, Fanisovna Hadieva G, Tafkilevich Lutfullin M, Valer'evna Khilyas I, Farvazovna Minnullina L, Gadelevna Gilyazeva A (2017). Bacillus subtilis strains with antifungal activity against the phytopathogenic fungi. Agricultural Sciences 8(1):1-20. |
|
|
Melo IS (2015). Rizobactérias Promotoras de Crescimento de Plantas: Descrição e Po-Tencial de Uso Na Agricultura. In: EMBRAPA Meio Ambiente, Ed., Ecologia Microbiana, Jaguariúna pp. 86-116. |
|
|
Mohamed EAH, Farag AG, Youssef SA (2018). Phosphate solubilization by Bacillus subtilis and Serratia marcescens isolated from tomato plant rhizosphere. Journal of Environmental Protection 9(3):266-277. |
|
|
Saeid A, Prochownik E, Dobrowolska-Iwanek J (2018). Phosphorus solubilization by Bacillus species. Molecules 23(11):2897. |
|
|
Saharan BS, Nehra V (2011). Plant growth promoting rhizobacteria: a critical review. Life Sciences and Medicine Research, 2021:1-30. |
|
|
Saravanakumar K, Yu C, Dou K, Wang M, Li Y, Chen J (2016). Synergistic effect of Trichoderma-derived antifungal metabolites and cell wall degrading enzymes on enhanced biocontrol of Fusarium oxysporum f. sp. cucumerinum. Biological Control 94:37-46. |
|
|
Saxena AK, Kumar M, Chakdar H, Anuroopa N, Bagyaraj DJ (2020). Bacillus species in soil as a natural resource for plant health and nutrition. Journal of Applied Microbiology 128(6):1583-1594. |
|
|
Shafi J, Tian H, Ji M (2017). Bacillus Species as versatile weapons for plant pathogens: a review. Biotechnology and Biotechnological Equipment 31(3):446-459. |
|
|
Zeilinger S, Gruber S, Bansal R, Mukherjee PK (2016). Secondary metabolism in Trichoderma - chemistry meets genomics. Fungal Biology Reviews 30(2):74-90. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


