Full Length Research Paper
ABSTRACT
This study examined the effect of post-natal lead exposure on the hippocampus of developing Wistar rats. Nine pregnant Wistar rats were randomly distributed into three groups of three rats each, consisting of a control Group 1 which received distilled water and experimental Groups 2 and 3 orally administered with 60 and 90 mg/kg bwt of lead acetate, respectively. The pups of the experimental Groups 2 and 3 were exposed to lead acetate via lactation from dams that were administered lead acetate from post-natal day (PND) 1 - PND 21. On PND 22, all the pups were weighed, and then euthanized. The brains of the pups were excised, weighed; lead deposit was quantified and fixed in Bouin’s fluid. The results revealed a significant (p<0.05) decrease in body weight, insignificant (p>0.05) decrease in brain weight, significant increase(p<0.05) in brain somatic index (BSI), significant increase(p<0.05) in lead deposition, weak staining for Nissl substance and distortion in cytoarchitecture of the hippocampus of Wistar rat’s pups exposed to lead acetate compared to the control. Post-natal exposure to lead acetate via lactation caused an increase in lead deposition, decrease in body weight and distortion in the cytoarchitecture of the hippocampus of developing Wistar rats.
Key words: Lead acetate, hippocampus, post-natal, lactation.
INTRODUCTION
Human child development is regulated by the interactions of both endogenous and exogenous factors (Gorini et al., 2014). One of the exogenous factors affecting early development of neurobehaviour in children is the exposure to heavy metals (Burger et al., 2011). Heavy metal poisoning has become a major health problem by nature of their environmental persistence (Adekomi, 2017), especially since the industrial revolution. Almost all organ systems are involved in heavy metal toxicity; however, the most predisposed systems include the nervous, renal, haematopoietic, and cardiovascular system (Arif et al., 2015).
Lead (Pb), one of these highly toxic heavy metals, has been detected in every facet of environmental and biological systems (Clark et al., 2009), particularly in industrialized cities. Lead is one of the oldest harmful agents known to mankind as its toxicity in both human and experimental animals (Abdulrazzaq et al., 2016) has been observed and documented since the historic times of the Greeks, Romans, Arabs and even the Egyptians.
Lead exists in three forms: metallic lead, inorganic lead and organic lead (ATSDR, 2017).
Human activities such as mining, manufacturing and burning fossil fuels can result in lead exposure. Lead is used in production of car batteries, hair dye, farm equipment, gasoline, paints, ceramics, cosmetics, water pipes, airplanes (La-Llave-León et al., 2016). In humans, other routes of exposure include the ingestion of lead-contaminated food or drinking water containing lead leaching from older corroding pipes, inhalation in industrial settings, and dermal contact (ATSDR, 2017). Children can be exposed to peeling lead-based paint or weathered powdered paint when engaging in hobbies. Children suffering from pica (the compulsive, habitual consumption of nonfood items) are particularly vulnerable (Sanders et al., 2009).
Lead is readily absorbed through the gastrointestinal tract and distributes into soft body tissues such as kidney, bone marrow, liver and brain, but accumulates in the blood and bone (Sansar et al., 2012; ATSDR, 2017). Studies have reported increase in accumulation of lead in the brain after exposure. However there is uncertainty on the mechanism by which lead (Pb) gets into the brain. Rahman et al. (2012) and Du et al. (2015) reported significantly higher level of lead accumulation in the brain of rat pups exposed to Pb postnatally, though this contrasted the work of Lindquist et al. (2015) who reported a non-significant accumulation of lead after perinatal exposure to lead. The activity of the hippocampus is distorted particularly in children by exposure to lead. Lead damages the cells within the hippocampus hence causing memory and learning deficit. Structural damages such as irregular nuclei and denaturation of myelin were also reported in rats. Lead usually interferes with the neurotransmitter glutamate which is important for many functions, like learning and memory. It operates by blocking N methyl –D- aspartate receptors (NMDAR) which is thought to be the main target of lead toxicity (Ab Latif et al., 2015).
However, the magnitude of the toxic response depends on several factors, including the dose, the age of the person exposed, the life stage of a woman (childhood, lactating, menopausal), occupational exposure, duration of exposure, health and lifestyle, and nutritional status of the person exposed. It is also believed that the greater absorptive capacity of the gastrointestinal tracts of children puts them at a higher risk than adults (Ab Latif et al., 2015). Breast milk has been suggested as a significant potential source of lead exposure to nursing infants, maternal blood and bone lead levels are both important determinants of lead in breast milk (Ettinger et al., 2014).
Furthermore, increasing prevalence of various abnormalities of the female reproductive system has been associated to lead exposure. Blood lead level above 10 μg/dl may adversely affect pregnancy outcome, resulting in increased risk of gestational hypertension, reduced length of gestation, miscarriage, spontaneous abortion, and preterm delivery (La-Llave-León et al., 2016). However, no blood-lead level is safe, because it has been reported that blood lead levels (BLLs) as low as 2 μg/dl are associated with lower measured intelligence, poorer memory, attention and reduced neurocognitive function in children (Sobin et al., 2013). Additionally in mouse and rodent models, early chronic exposure to Pb resulted in abnormal motor, decreased memory and exploratory behavior (Kasten-Jolly et al., 2012). The chief mechanism of lead induced toxicity has been reported to be via oxidative stress in most biological systems (Flora et al., 2012). Since the 1970s, regulation reducing lead in products has greatly reduced exposure in the developed world, but it is still vehemently used in products in many developing countries (Ab Latif et al., 2015; ATSDR, 2017). The continuous use of lead in several products in developing countries necessitates a detailed investigation on its resultant effects on the brain. While peripheral effects in adults often go away when lead exposure ceases, evidence suggests that most of lead's effects on a child's central nervous system are irreversible (Bellinger, 2004; ATSDR, 2017). In this study we examined the effect of post-natal lead exposure on the hippocampus of developing Wistar rats.
MATERIALS AND METHODS
Ethical approval
Ethical approval was obtained from Ahmadu Bello University Ethics Committee on Animal Use and Care with approval number ABUCAUC/2018/047.
Experimental animals
Nine non pregnant and three male Wistar rats weighing between 120 to 150 g were used for this study. They were housed in standard rat cages under suitable environmental conditions, fed with standard pelletized feed and water ad libitum and were maintained in this condition for a period of two weeks (14 days) to acclimatize them prior to experimental study.
Experimental chemical
Analytical grade of lead acetate of 99.9% purity with batch number (CAS 304-55-2) manufactured by Best of chemical limited, New York, USA was purchased.
Mating of rats and confirmation of pregnancy
During acclimatization all female Wistar rats were kept in the same cage to synchronize their estrus cycle (McClintock, 1978). Determination of the estrus cycle was carried out adopting the method of Marcondes et al. (2002) and Adebisi (2008). On the day of proestrus, female rats were caged overnight with sexually matured male rats in ratio of 3 females to one male. A positive mating was confirmed by vaginal plug examination under the light microscope early in the morning of the day after mating (Marcondes et al., 2002).
Experimental design
When the rats were confirmed pregnant, they were separated and kept in separate rat cages containing one dam each and her pups. While in separated rat cages, pregnant Wistar rats were randomly distributed into three experimental groups of three rats each, consisting of a control Group 1 and experimental groups 2 and 3. The dams in each of the three cages in each group gave birth to an average of five pups and an average of fifteen pups was obtained per group. The day of birth was taken as post-natal Day 1 (PND 1). Based on the reported oral LD50 of lead acetate which is 600 mg/kg body weight for Wistar rats (Sujatha et al., 2011), all the pups of the experimental groups 2 and 3 were exposed to lead acetate via lactation from dams exposed to 10 and 15% of the LD 50 of lead via oral gavage respectively for three weeks (from post-natal day (PND) 1 – PND 21). The control Group 1 was given distilled water throughout the experimental period.
Body weight assessment
After administration, a total of ten pups were selected at random from each group. Prior to the day of sacrifice, the ten pups per group were fasted overnight by separating them from their mothers. On the day of sacrifice (PND 22), the pups in each group were weighed using a sensitive weighing balance manufactured by 3 Dresyns with sensitivity of 0.01 g in the Department of Human Anatomy, Ahmadu Bello University Zaria, Nigeria. The resultant weights were recorded.
Anesthesia
On the day of sacrifice (PND 22), five pups each from the control Group 1 and experimental Groups 2 and 3 were subjected to anesthesia (ketamine at 75 mg/kg bwt intraperitoneally) (Wellington et al., 2013). Their brains were quickly harvested by making a mid-sagittal incision through the skull and fixed in Bouin’s fluid. The tissues were routinely processed and stained using routine Haematoxylin and Eosin method and with Cresyl violet stain for Nissl substance.
Brain weight assessment and brain somatic index estimation
Immediately after excision of the brains from the skull of the pups they were quickly weighed using a sensitive weighing balance with sensitivity of 0.01 g manufactured by 3 Dresyns, Barcelona, Spain. After weighing the brain, the resultant weights were recorded. The brain somatic index was calculated from the formula below:
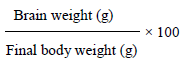
Quantification of lead deposit in the brain
The concentration of lead deposit was carried out by Atomic Absorption Spectrophotometry (Sharafi et al., 2015) using Atomic Absorption Spectrophotometry-lamp standard (AAS-L) model AA 6800 Shimadzu Japan.
Photomicrography
After staining, microscopic slides were viewed under the microscope using X250 magnifications and photomicrographs were taken using MD900 Amscope® digital camera.
Statistical analysis
Data obtained from the study were expressed as Mean±SE (Standard error). One way analysis of variance (ANOVA) was used to compare the mean differences between and within the groups followed by LSD post-hoc test. P-value less than 0.05 was considered statistically significant. Data were analyzed using Statistical Package for Social Sciences (IBM SPSS 20).
RESULTS
Body weight assessment of Wistar rat pups
There was a significant (p<0.05) decrease in body weight of Wistar rat pups in Groups 2 and 3 when compared with the control (Table 1).
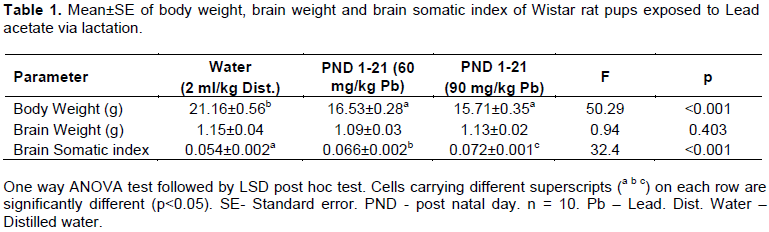
Brain weight and brain somatic index of Wistar rat pups
The brain weight of Wistar rats pups in Groups 2 and 3 were insignificantly (p>0.05) lower than that of the control (Table 1). The brain somatic index significantly (p<0.05) increased in Wistar rats pups of Groups 2 and 3 when compared with the control (Table 1). Between the groups, the brain somatic index also significantly (p<0.05) increased in pups of Group 3 when compared to Group 2 (Table 1).
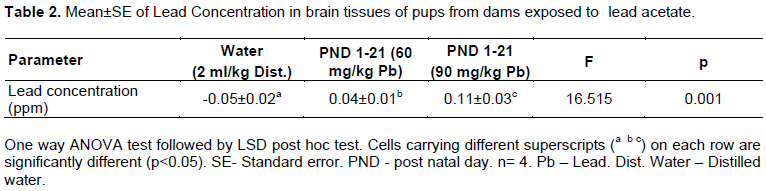
Lead concentration
Lead accumulation in brain tissues in Wistar rat pups of Groups 2 and 3 was significantly (p<0.05) higher than the values obtained in the control (Table 2). The lead concentration in the brains of the pups from Group 3 was also significantly (p<0.05) higher than the values obtained for pups from Group 2 during the experiment (Table 2).
Histological observations
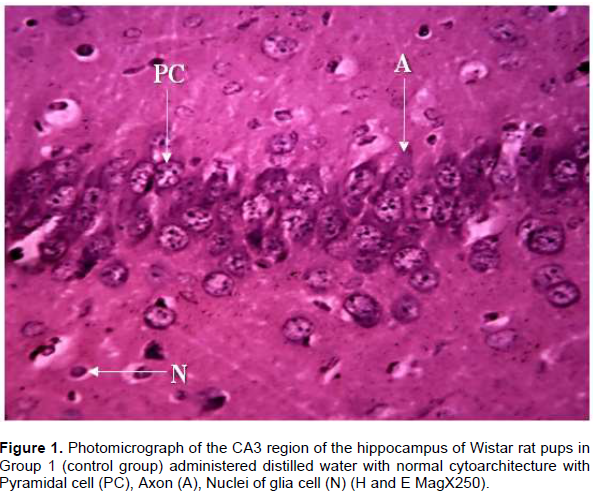
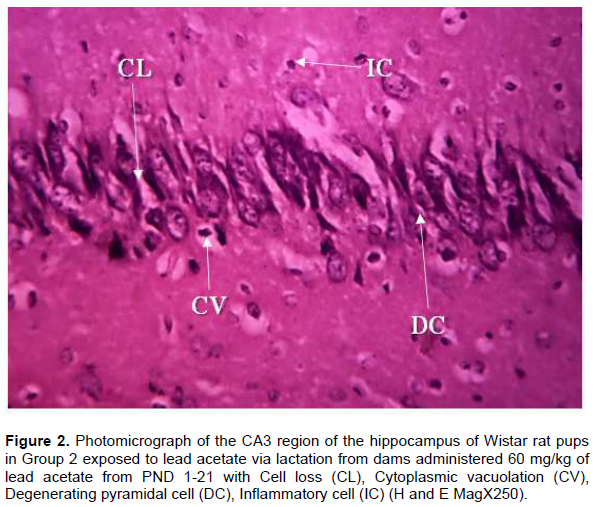
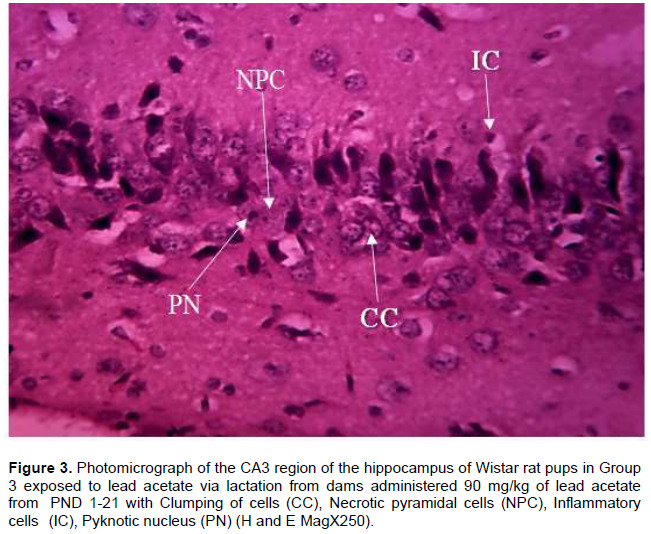
Histological observations of the CA3 region of the hippocampus of Wistar rat pups in Group 1 (control group) administered distilled water revealed normal cyto-architecture and characteristic appearance of the pyramidal cell with well-preserved cytoplasm, moderately dispersed neuroglia cells, prominent nuclei and nucleoli and axon radiating from the cell body (Figure 1). The CA3 region of the hippocampus of Wistar rat pups in Group 2 revealed distortion in cytoarchitecture of the hippocampus with presence of cell loss, inflammatory cells, cytoplasmic vacuolation and degeneration of pyramidal cells (Figure 2). In Group 3 Wistar rat pups, the cytoarchitecture of CA3 region of the hippocampus was distorted with the presence of necrotic pyramidal cell, more inflammatory cells, pyknotic nucleus and clumping of cells (Figure 3).
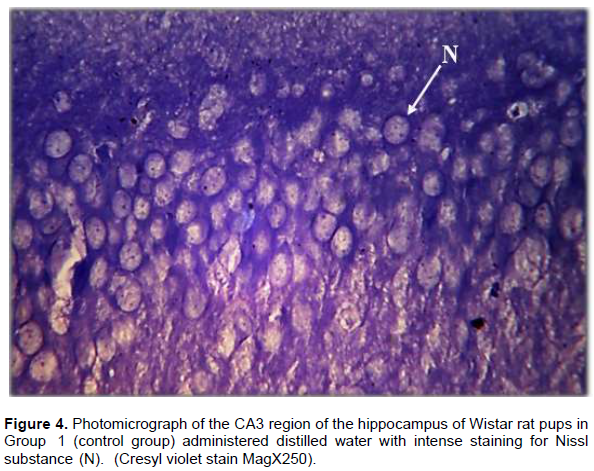
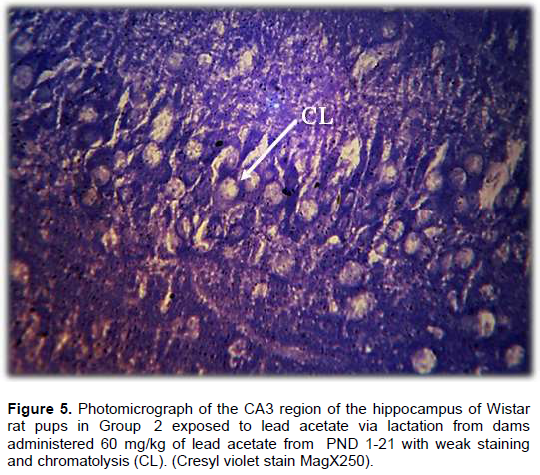
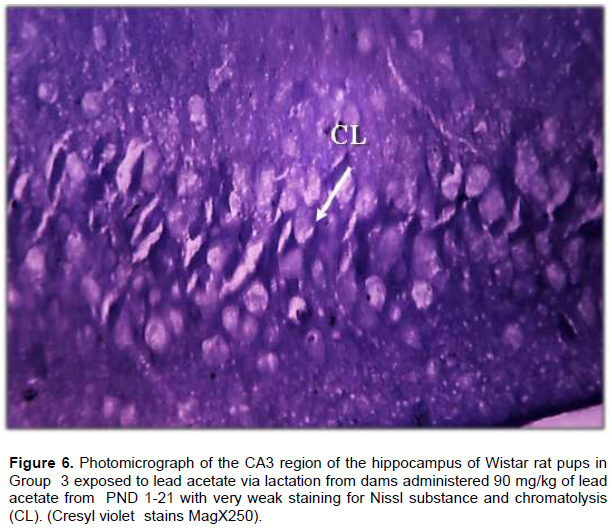
Result from Nissl staining of the CA3 region of the hippocampus showed intense staining of the basophilic ribosomes in the perikaryon of the pyramidal cells (Nissl substance) after cresyl violet staining (Figure 4). In Wistar rat pups of Group 2, the pyramidal cells showed weak staining for Nissl substance when compared with the control but more intense staining when compared with Group 3 (Figure 5). The CA3 region of the hippocampus of Wistar rat pups in Group 3 revealed very weak staining for Nissl substance when compared with the control (Figure 6).
DISCUSSION
Lead metal causes toxicity in living cells either by ionic mechanism or that of oxidative stress (Flora et al., 2012). Compared to adults, children are more predisposed to its toxicity due to the greater absorptive capacity of their gastrointestinal tracts (Arif et al., 2015). The present study was designed to assess the effects of maternal exposure to different doses of lead acetate during lactation on the Hippocampus of developing Wistar rats. In this study a significant decrease in body weight was observed in Wistar rats pups exposed to lead from PND 1-21 when compared with the control. This suggests that exposure to lead may cause serious loss of body weight in developing infants. This decrease may be as a result of decline in the consumption of food (anorexia) which is induced by heavy metal consumption. Another possible explanation for the loss of body weight may be decreased muscle mass and cachexia due to the oxidative stress induced by lead (Zaheer et al., 2013). Aprioku and Siminialayi (2013) reported significant and time-dependent reductions in body weight after perinatal exposure to 4 or 8 mg/kg lead acetate respectively. The result from the present study is dissimilar to the findings of Lewis and Pitts (2004) who reported an insignificant decrease in body weight of Wistar rat pups after 3-week post-natal exposure to lead acetate at (50, or 250 ppm) when compared with the control. The difference observed in the result of Lewis and Pitts (2004) with that of the present study may be due to the differences in the doses used in the studies. The findings from the present study also showed an insignificant decrease in brain weight of Wistar rats pups treated with lead acetate from PND 1-21 when compared with the control. The result from this study is dissimilar to the work of Alyaa et al. (2015) who reported a significant reduction in fetal brain weight of Sprague–Dawley rat pups exposed to lead from dams taking drinking water with 0.5 and 1% lead nitrate for two weeks before conception, and during pregnancy when compared to the control group. The reason for dissimilarity in the result of the present study and that of Alyaa et al. (2015) could be due to the difference in lead compound administered to the rats, the period and duration of administration as well as the species of the animals used for the experimental study. These finding is also dissimilar to the findings of Corpas et al. (2002) who reported that testes weights significantly decreased in experimental pups exposed to lead perinatally when compared to the control. In this study an insignificant increase in the brain weight of Wistar rat pups in Group 3 in comparison with Wistar rat pups in Group 2 was observed. This is dissimilar to the findings of Saeed (2016) who reported a significant increase in the weight of the Liver, Kidney, Heart and Spleen with increasing concentration of lead in male and female albino rats treated with sub lethal doses of lead acetate (0.4, 0.8, and 1.2 mg/kg body weight, respectively) in drinking water for 12 weeks. The dissimilarity in the result from this study and that of Saeed (2016) could be due to the differences in the duration of exposure to lead. A significant increase in brain somatic index was also observed in the lead exposed group when compared with the control. From this result, it can be deduced that the brain weight of Wistar rat pups exposed to lead acetate via lactation did not reduce as rapidly as their body weight after exposure to lead. That is, the decrease in body weight observed could be as a result of the decrease in weight of other organs of the body and not necessarily a consequence of decrease in brain weight. This result also suggests that the decrease in body weight observed in Wistar rat pups could be as a result of lead causing irritation of the gastrointestinal tract and disruption of the activity of the lateral hypothalamus, hence reducing food consumption. This is similar to the findings of Kang et al. (2004) who reported significantly higher brain somatic index in high dose lead treated group when compared with the control. The mechanism by which Pb gets into the brain is not clear. One potential mechanism is the disruption of the blood-brain barrier (BBB) by Pb. However, findings on the effects of Pb on the integrity of BBB are inconsistent. In the present study it was observed that lead exposure resulted in significant accumulation of lead in the Pb-exposed Wistar rat pups when compared to the control. Increased accumulation of Pb in Wistar rat pups exposed to lead acetate may have been as a result of lead crossing the BBB by passing through the endothelial cells at the blood brain barrier because it can substitute for calcium ions and be taken up by calcium-ATPase pumps (Terry, 2009; Rai et al., 2010), since the blood brain barrier in the developing brain is not fully developed. It is also possible that Pb might have been transported into the brain by the divalent metal ion transporter 1 (DMT1), which is localized in astrocytes and neurons in the normal brain (Wang et al., 2011). This is similar with the findings of Rahman et al. (2012) who reported a significant accumulation of lead in the brain of Wistar rat pups that were exposed to 0.2% lead acetate via their dams’ drinking water from PND 1 to 21. Du et al. (2015) also reported significantly higher level of lead accumulation in Sprague–Dawley rats exposed to Pb from parturition to PND 30. The result of this study is dissimilar to the work of Lindquist et al. (2015) who reported a non-significant accumulation of lead after perinatal exposure to lead. The primary target of low level lead exposure is the central nervous system. The developing brain appears to be especially vulnerable to lead neurotoxicity. Lead neurotoxicity causes impairment of cognitive abilities and a significant delay in behavioural development, especially in children. Microscopic examination of lead treated groups in this study revealed various degrees of histological changes in the CA3 region of the hippocampus which includes presence of necrotic pyramidal cell, cell loss, cytoplasmic vacuolation, presence of degenerating pyramidal cells, inflammatory cells, clumping of cells, and pyknotic nucleus when compared with the control group. This suggests the toxic effects of post-natal lead exposure. These changes were dose dependent, with higher doses causing more damaging effects. Necrosis of pyramidal cells may have been as a result of oxidative stress due to the depletion of the antioxidant system by post-natal lead exposure as chronic lead exposure has been reported to cause necrosis of hepatocytes due to oxidative stress on these cells by glutathione depletion (El Sayed et al., 2015). The presence of pyknotic nucleus and degenerating cell was in agreement with the reports of Omayma et al. (2011) and Abbas (2015) who stated that lead administration of different dose levels caused pyknosis of nucleus and degeneration of neurons. The increase in inflammatory cells seen in our result as a post-natal exposure to lead may increase the production and release of inflammatory cytokines, enhance generation of reactive oxygen species (ROS), impede antioxidant activity, and may be the reason for the distortion of the cytoarchitecture of the CA3 region of the hippocampus. The cytoplasmic vacuolation observed may have resulted from disturbance in the oxidative phosphorylation in the mitochondria with suppression of ATP production and failure of the ATP dependent sodium pump at the cell membrane. This resulted in accumulation of sodium intracellularly and consequent entry of water into the different cellular compartments which results in cellular swelling (Haouas et al., 2014). Nissl substances are seen as dark spots in an optical microscope. They are dispersed throughout the neuronal perikaryon. At the ultrastructural level, they correspond to 1-4 μm regions abundant with granular endoplasmic reticulum, ribosomes and polysomes (Fedorenko and Uzdensky, 2010). Cresyl violet stain for Nissl substance of lead treated groups in this study showed weak staining for Nissl substance within the perikaryon of the pyramidal cell of the hippocampus when compared with the control. These changes were dose dependent, with higher doses causing more damaging effects. This is analogous with the work of Adekomi et al. (2017) who reported dispersed perinuclear Nissl deposits in cerebral cortex of male Wistar rat exposed to 5 mg/kg Pb-nitrate orally for 21 days. Syed (2015) and Hamza et al. (2017) also reported loss of Nissl substance from Purkinje cells of the cerebellar cortex in Wistar rats exposed to varying doses of lead. The mossy fibres of the granule cell in the dentate gyrus conduct neurotransmitter from the perikaryon of the granule cells to the dendrite of the pyramidal cells. Degeneration of the Nissl bodies decrease synthesis of neurotransmitter which in turn affects the transmission of impulses to the pyramidal cells of the CA3 region of the hippocampus. This leads to anterograde and retrograde amnesia. A limitation of the present study is the constraint in performing neurobehavioural test for memory and biochemical analysis for oxidative stress markers. Another limitation of the study is the small number of Wistar rat dams used per group.
CONCLUSION
It can be inferred that exposure of Wistar rat pups to increasing doses of lead acetate via lactation induced a decrease in body weight and an increase in brain somatic index. Post-natal lead exposure also caused accumulation of lead in the brain of the Wistar rat pups and distortion in the cytoarchitecture of the CA3 region of the hippocampus which may lead to learning and memory deficit.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
The authors appreciate the support given by the staff of the Department of Human Anatomy, Ahmadu Bello, University, Zaria-Nigeria and staff of National Research Institute for Chemical Technology (NARICT), Zaria, Nigeria and also thank Prof. B. Mohammed of the Department of Veterinary Pathology for assisting in the interpretation of the slides.
REFERENCES
|
Ab Latif W, Anjum A, Jawed AU (2015). Lead toxicity: a review. Interdisciplinary Toxicology 8(2):55-64. |
|
|
Abbas ZA (2015). The possible protective role of vitamin E on the joined neurobehavioural effects of lead toxicity and noise stress in rats. Journal of International Academic Research for Multidisciplinary 3(4):256-268. |
|
|
Abdulrazzaq MA, Mohammed MH, Wahid AH, Yusof MS, Mohammed AR (2016). Prophylactic effect of Nigella sativa against lead acetate induced changes in spermiogram, reproductive hormones and gonadal histology of rats. Veterinary World 9(6):660-671. |
|
|
Adebisi SS (2008). The toxicity of artesunate on bone developments: the Wistar rat animal model of malaria treatment. The Internet Journal of Parasitic Diseases 4(1):1-8. |
|
|
Adekomi DA, Adewole OS, Tijani AA, Adeniyi TD (2017). Lead induces inflammation and neurodegenerative changes in the rat medial prefrontal cortex. Anatomy 11(2):79-86. |
|
|
Adekomi DA, Adewole OS, Tijani AA, Adeniyi TD (2017). Lead induces inflammation and neurodegenerative changes in the rat medial prefrontal cortex. Anatomy 11(2):79-86. |
|
|
Alyaa MM, Ameera SA, Muddanna SR, Narayana K (2015). Gestational lead exposure induces developmental abnormalities and up-regulates apoptosis of fetal cerebellar cells in rats. Drug and Chemical Toxicology 38(1):73-83. |
|
|
Aprioku JS, Siminialayi IM (2013). Maternal lead exposure and pregnancy outcome in Wistar albino rats. Journal of Toxicology and Environmental Health Sciences 5(10):185-193. |
|
|
Arif TJ, Mudsser A, Kehkashan S, Arif A, Inho C, Qazi M, Rizwanul H (2015). Heavy Metals and Human Health: Mechanistic Insight into Toxicity and Counter Defense System of Antioxidants. International Journal of Molecular Sciences 16:29592-29630. |
|
|
ATSDR (2017). Agency for Toxic Substances and Disease Registry. Toxicological Profile for Lead. Update. U.S. Department of Health and Human Services. Public Health Service 1-185. |
|
|
Bellinger DC (2004). Lead. Pediatrics 113(4):1016-1022. |
|
|
Burger J, Gochfeld M, Jeitner C (2011). Locational differences in mercury and selenium levels in 19 species of saltwater fish from New Jersey. Journal of Toxicology and Environmental Health part A 74(13):863-874. |
|
|
Clark CS, Adebamowo CA, Adebamowo EO, Agbede OA, Chen CK, El-Safty A, Menrath W, Rampal KG, Roda SM, Shinde RM, Sridhar MKC, Succop P, Thuppil V, Yu J, Zakaria Y (2009). Lead levels in new enamel household paints from Asia, Africa and South America. Environmental Research 109:930-936. |
|
|
Corpas I, Castillo M, Marquina D, Benito MJ (2002). Lead intoxication in gestational and lactation periods alters the development of male reproductive organs. Ecotoxicology and Environmental Safety 53:259-266. |
|
|
Du Y, Ge MM, Xue W, Yang QQ, Wang S, Xu Y, Hui-Li W (2015). Chronic Lead Exposure and Mixed Factors of Gender×Age×Brain Regions Interactions on Dendrite Growth, Spine Maturity and NDR Kinase. PLoS ONE 10(9):e0138112. |
|
|
El Sayed AM, Ahmed NF, Rania AS, Eman MN (2015). Anatomical and Histological Study of the Effect of Lead on Hepatocytes of Albino Rats. International Journal of Biomedical Materials Research 3(4):34-45. |
|
|
Ettinger AS, Roy A, Amarasiriwardena CJ, Smith D, Lupoli N, Mercado-García N, Lamadrid-Figueroa H, Tellez-Rojo MM, Hu H, Hernández-Avila M (2014). Maternal Blood, Plasma, and Breast Milk Lead: Lactational Transfer and Contribution to Infant Exposure. Environmental Health Perspectives 122(1):87-92. |
|
|
Fedorenko GM, Uzdensky AB (2010). Nissl substance and cellular structures involved in the intraneuronal and neuroglial transport in the crayfish stretch receptor. Microscopy: Science, Technology, Applications and Education. Méndez-Vilas A, Díaz J (Eds.) pp. 299- 306. |
|
|
Flora G, Gupta D, Tiwari A (2012). Toxicity of lead: A review with recent updates. Interdisciplinary Toxicology 5(2):47-58. |
|
|
Gorini F, Chiappa E, Gargani L, Picano E (2014). Potential effects of environmental chemical contamination in congenital heart disease. Pediatric Cardiology 35(4):559-568. |
|
|
Hamza GA, Ibegbu AO, Buraimoh AA (2017). Evaluation of the Effects of Aqueous Garlic Extract on Lead-Induced Changes on Cerebellum of Wistar rats. African Journal of Cellular Pathology 8:9-14. |
|
|
Haouas Z, Sallem A, Zidi I, Hichri H, Mzali I, Mehdi M (2014). Hepatotoxic Effects of lead acetate in rats: histopathological and cytotoxic studies. Journal of Cytology and Histology 5(5):1-6. |
|
|
Kang JK, Donggeun S, Kang JK, Sang-Yoon N, Hae-Joon K, Lee E (2004). Effects of Lead Exposure on the Expression of Phospholipid Hydroperoxidase Glutathione Peroxidase mRNA in the Rat Brain. Toxicological Sciences 82:228-236. |
|
|
Kasten-Jolly J, Bolivar VJ, Lawrence DA, Pabello N (2012). Developmental lead effects on behavior and brain gene expression in male and female BALB/cAnNTac mice. Neurotoxicology 33:1005-1020. |
|
|
La-Llave-León O, Pacheco MS, Martínez SE, Rodríguez EE, Francisco X, Castellanos J, Carrillo AS, Quiñones AM, Alanís FV, Vargas GG, Hernández MM, Sustaita JD (2016). The relationship between blood lead levels and occupational exposure in a pregnant population. BMC Public 16:1231. Lewis MW, Pitts DK (2004). Inorganic lead exposure in the rat activates striatal cFOS expression at lower blood levels and inhibits amphetamine-induced cFOS expression at higher blood levels. Journal of Pharmacology and Experimental Therapeutics 310(2):815-820. |
|
|
Lindquist DM, Beckwith T, Cecil KM, Sánchez-Martín FJ, Landero-Figueroa J, Puga A (2015). Prenatal and early postnatal lead exposure in mice: neuroimaging findings. Quantitative Imaging in Medicine and Surgery 4:511-518. |
|
|
Marcondes FK, Bianchi FJ, Tanno AP (2002). Determination of the estrous cycle phases of rats: Some helpful considerations. Brazilian Journal of Biology 62(4a):609-614. |
|
|
McClintock MK (1978). Synchronizing Ovarian and Birth Cycles by Female Pheromones. Chemical Signals in Vertebrates 3:159-178. |
|
|
Omayma KH, Ayman MM, Nadra K (2011). Effect of antioxidants on hippocampal structure in male albino rats exposed to lead toxicity: histological and biochemical study. The Egyptian Journal of Histology 34:808-817. |
|
|
Rahman A, Khan KM, Al-Khaledi G, Khan I, Sreeja A (2012). Early postnatal lead exposure induces tau phosphorylation in the brain of young rats. Acta Biologica Hungarica 63(4):411-425. |
|
|
Rai A, Maurya SK, Khare P, Srivastava A, Bandyopadhyay S (2010). Characterization of developmental neurotoxicity of As, Cd, and Pb mixture: synergistic action of metal mixture in glial and neuronal functions. Toxicology of Science 118:586-601. |
|
|
Saeed AA (2016). Hematobiochemical changes induced by lead intoxication in male and female albino mice. National Journal of Physiology, Pharmacy and Pharmacology 6(1):46-51. |
|
|
Sanders T, Liu Y, Buchner V, Tchounwou PB (2009). Neurotoxic Effects and Biomarkers of Lead Exposure: A Review. Review on Environmental Health 24(1):15-45. |
|
|
Sansar W, Ahboucha S, Bouyata M, Gamrani H (2012). Effects of chronic lead intoxication on rat serotoninergic system and anxiety behavior. Acta Histochemica 114(1):41-45. |
|
|
Sharafi K, Fattahi N, Pirsaheb M, Yarmohamadi H, Fazlzadeh-Davil M (2015). Trace determination of lead in lipsticks and hair dyes using microwaveâ€assisted dispersive liquidâ€liquid microextraction and graphite furnace atomic absorption spectrometry. International Journal of Cosmetic Science 37(5):489-495. |
|
|
Sobin C, Flores MGM, Parisie N, Schaube T, Cervantesa M, Rodrigo XA (2013). Microglial disruption in young mice with early chronic lead exposure. Toxicology Letters 220(1):44-52. |
|
|
Sujatha K, Srilatha C, Anjaneyulu Y, Amaravathi P (2011). Lead acetate induced neurotoxicity in Wistar albino rats: A pathological, immunological, and ultrastructural studies. The Journal of Pharmaceutical and Biological Science 2:459-462. |
|
|
Syed ZN (2015). A Comparative Study of the Histological Changes in Cerebral Cortex, Hippocampus, Cerebellum, Pons & Medulla of the Albino rat due to Lead Toxicity. International Journal of Anatomy and Research 3(2):1173-1178. |
|
|
Terry JAV (2009). Spatial Navigation (Water Maze) Tasks. In: Buccausco JJ. Methods of Behaviour Analysis in Neuroscience. CRC Press, Boca Raton pp. 13.1-13.4. |
|
|
Wang Q, Luo W, Zhang W, Liu M, Song H, Chen J (2011). Involvement of DMT1+IRE in the transport of lead in an in vitro BBB model. Toxicology in Vitro 25:991-998. |
|
|
Wellington D, Mikaelian I, Singer L (2013). Comparison of Ketamine-Xylazine and Ketamine-Dexmedetomidine Anesthesia and Intraperitoneal Tolerance in Rats. Journal of the American Association for Laboratory Animal Science 52:481-487. |
|
|
Zaheer A, Muhammad ZI, Amir AS (2013). Lead-Induced Reduction in Body and Kidney Weight of Wistar Albino Rats Ameliorated by Ginkgo biloba Extract (EGb 761). Biochemical Physiology 2(2):2. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


