Full Length Research Paper

ABSTRACT
Hawthorn (Crataegus Mexicana) is a traditional fruit in Mexican gastronomy and is used to treat many ailments. Previous studies have shown that acetone extracts derived from hawthorn peel (HPE) possess a strong antioxidant activity in chemical and biological model systems in vitro, attributable to their polyphenolic content. The main objective of this study was to investigate the ability of HPE to protect erythrocytes against oxidative damage, in vitro. The protection rendered by the HPE in erythrocytes was studied in terms of protection to oxidative damage by the inhibition of thiobarbituric acid reactive substances (TBARS) assays, morphological changes by light microscopy, and electrophoretic analysis of banding pattern of the red blood cells (RBCs). FeSO4 was chosen to induce lipid peroxidation in human RBCs membranes and cytoskeleton proteins. The total polyphenol content in HPE was found to be 0.68 mg/g (SD 0.001) as the equivalent of gallic acid per gram. Trapping of DPPH was calculated by IC50 in 15.26 mg/L (SD 0.20). Better inhibition of TBARS formation by HPE was 16.78 mg/L (SD 0.33). HPE retards the morphological alteration in the erythrocytes-eryptosis. The electrophoretic pattern showed that some protein bands were not altered during a long period of incubation in HPE. Furthermore, it was found that HPE offers significant protection to human membrane erythrocyte up to for 28 days from the oxidative damage. In conclusion, our results indicate that HPE is capable of protecting erythrocytes against oxidative damage and morphologic changes by acting as a strong antioxidant.
Key words: Antioxidants, Crataegus mexicana, free radicals, Hawthorn, red blood cells, thiobarbituric acid reactive substances (TBARS).
INTRODUCTION
The pathophysiology of many blood diseases is associated with an increase of free radicals derived from reactive oxygen species (ROS) in the blood cells. Oxidative stress in red blood cells (RBCs) induces damage by injuring the protein cytoskeleton (Berlett and Stadtman, 1997; Dean et al., 1997), and the phospholipid cell membrane (Kowalczyk et al., 2012) causing directly and indirectly morphologic and microrheologic changes (Hebbel et al., 1990), thus premature eryptosis (Nagababu et al, 2008; Kempe et al., 2006; Lang et al., 2006). These changes and the reduced life span of the cells are involved in the pathogenesis of a number of blood diseases, including different types of anemia, altered vascular disorders, coagulopathies or clog veins alterations, also it is important in the life span of RBCs’ bags (Baek et al., 2012). ROS are also involved in a number of other chronic human diseases (Kowalczyk et al., 2012).
In the bi-lipid cell membrane of the erythrocytes oxidation occurs by ROS, and malondialdehyde (MDA) formation can be measured and determined due the altered redox status of the cell (Singh and Rajini, 2008). The higher production of ROS is capable of causing oxidative damage in proteins by oxidation of amino acid residue side chain, formation of protein-protein cross-link-ages, and oxidation of the protein backbone resulting in protein fragmentation (Berlett and Stadtman, 1997). Because of RBCs do not have nucleus or mitochondria organelles, their defense and reparation mechanisms against ROS became vulnerable (Pandey and Rizvi, 2011). However, RBCs have an effective anti-oxidant system that includes high levels of the enzyme catalase (CAT) (Cañas et al., 1988; Aebi et al., 1968), lower levels of superoxide dismutase (SOD) (Lynch et al., 1976), and the reduced glutathione peroxidase (GSH) (Van Asbeck et al., 1985), that make this cells mobile free radical scavengers.
A large number of fruits containing high amounts of nutrients are capable of inhibiting the oxidative stress and ROS formation; this is possible mainly by the action of antioxidant molecules, such as vitamin C, polyphenols, flavonoids and other antioxidant enzyme systems (Edwards et al., 2012; Cai et al., 2004). The clinical benefits after the intake of fruits have shown that they can protect and prevent the complications of chronic degenerative diseases, such as atherosclerosis, chronic inflammation, diabetes mellitus, cataract, coronary diseases and some types of cancer (Edwards et al., 2012; Willis and Wians, 2003; Middleton et al, 2000). Antioxidant activity of ethanolic plant extracts on cells has been demonstrated (Salawu et al., 2011; Nugraheni et al., 2011), including the inhibitory effects in RBCs hemolysis by the action of antioxidant extracts of cocoa plant (Zhu et al., 2005, 2002), potato peel (Singh and Rajini, 2008), and flaxseed oil (Liu et al., 2012). The activity has been related to flavonoids (Lam et al., 2007; Asgary et al., 2005; Sorata et al., 1984) to protect the erythrocytes membrane against deformation.
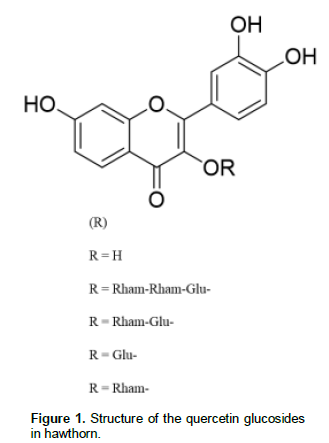
The hawthorn Crataegus mexicana, is a traditional fruit which is a medicinal herb and in the Mexican gastronomy since ancient times. The wild type also named “tejocote” is widespread in almost all around Mexico. This variety of hawthorn has been the subject of little clinical studies for its medicinal properties and benefits (Arrieta et al., 2010; Andrade-Cetto and Heinrich, 2005) and only one study has been conducted to determine its phytochemical composition in their flowers (García-Mateos et al., 2012) showing the presence of polyphenols such as quercetin 3-O-glucoside, quercetin 3-O-rhamnoside, quercetin 3-O-rhamnosyl-(1→2)-[rhamnosyl-(1→6)]-glucoside, and quer-cetin 3-O-rhamnosyl-(1→6)-glucoside (rutin) (Figure 1).
In a previous study, we have demonstrated the anti-oxidant capacity of the hawthorn through lipid peroxi-dation inhibition in in vitro rat brain cells (Méndez-Iturbide et al., 2013). In this context to gain insight into the potential interaction of erythrocytes with the antioxidants found in Mexican fruits the present work aimed to assess the antioxidant activity of the acetone extract derived from the hawthorn peel, to inhibit lipid peroxidation and morphology changes in the red blood cell membrane during the life span of the erythrocyte in vitro experiments. The purpose of the study was to test the hypothesis that lipid peroxidation on the erythrocyt cell membrane is inhibited by HPE from C. mexicana.
MATERIALS AND METHODS
For TBARS analysis, the following reagents were purchased from Sigma-Aldrich Co (St Louis, MO, USA): Polyphenol (quercetin), ferrous sulphate (FeSO4), NaCl, NaOH, KCl, NaHPO4, KH2PO4, trichloroacetic acid, ethylenediaminetetraacetic acid (EDTA), and 2-thiobarbituric acid (TBA).
For Lowry protein concentration, the following reagents were acquired: Bovine Serum Albumin (BSA) [A-7906], Folin–Ciocalteu reagent, phosphate buffer saline (PBS) (9.5 mM, pH 7.4), Na2CO3, 2%; NaOH, 0.1 N; tetrahydrate potassium sodium tartrate 2% [KNaC4H4O6.4H2O] and CuSO4, 1% (Sigma-Aldrich Co. St Louis, MO, USA).
For scavenging effect on free radicals, 2,2-diphenyl-1-picrylhydrazyl (DPPH) was purchased from Sigma-Aldrich Co. (St Louis, MO, USA).
To measure total phenolic compounds, Folin-Ciocalteu reagent, 0.3% HCl, sodium carbonate (2% Na2CO3), and gallic acid, were purchased from Sigma-Aldrich Co. (St Louis, MO, USA).
Cell staining was achieved by preparing 10% neutral buffered formalin, and Haematoxylin-Eosin (HE) stain solutions (Sigma-Aldrich Co. St. Louis, MO, USA), ethanol, xylene, and mounting resin were purchased from J.T. Baker Chemical Co. (Phillipsburg, NJ, USA).
For SDS-PAGE, the following reagents were purchased: Sodium chloride, isobutyl alcohol, methanol, and acetic acid were acquired from J.T. Baker Chemical Company (Phillipsburg, NJ, USA). SDS, Tris, glycine, coomassie brilliant blue R-250, silver stain kit, TEMED, ammonium persulfate, N,N-1-methyl-bis-acrylamide, acryl-amide and low molecular weight markers were from Bio-Rad (Richmond, CA, USA).
All other reagents including 2d-deionized water (18.3 MW, System Milli Q-II. Millipore, USA) were high quality analytical grade and pyrogenous free.
Source of samples
Four kilograms of hawthorn fruits were gathered in the State of Tlaxcala, México, in the month of December 2013. All samples had the same morphological characteristics in size, color, and damage free of the surface. Only skin samples of hawthorn were studied. The fruits were hand washed and the skin was manually peeled off, frozen and lyophilized. A total of 36.0 g of peel was obtained and then mashed (mesh size 20). Samples were maintained frozen at -20°C until use.
Preparation of hawthorn fruits peels extracts
Thirty grams of the resulting powder were homogenized at room temperature. Then, the homogenate was sequentially extracted for 72 h using 300 mL of the following solvents: n-hexane, dichloromethane, ethyl acetate, acetone, ethanol and finally methanol to obtain from the same homogenate one extract per solvent. All the extracts were decanted and evaporated until dry and kept inside a closed amber flask at 5°C until use. Only the acetone HPE was used for this study.
Scavenging effect on DPPH
The scavenging effect of antioxidants in HPE on free radicals was determined by using DPPH (Kim et al., 2005). Briefly, 100 μL of HPE were prepared at a concentration of 20 mg/L in DMSO. Then, 4000, 400, and 40 mg/L final dilutions in cool ethanol were prepared. DPPH was dissolved in ethanol (1 mM). A total of 50 μL of HPE and 150 μL of DPPH (final concentration of 1000, 100, and 10 mg/L of HPE) were mixed and incubated for 30 min at 37°C in an orbital shaker. The absorbance at 517 nm was recorded using an Elisa microplate reader (Bio-Tek EL800, USA). The percent-reduced activity of DPPH was calculated and expressed as the IC50 (Khattak et al., 2008) using six different concentrations of HPE (5.6 - 100 mg/L). Quercetin was used as standard.
Determination of total phenolic compounds
The total phenol content of the acetone HPE was determined using the Folin-Ciocalteu reagent method (Folin and Ciocalteu, 1927) with minor modifications as described previously (Méndez-Iturbide et al., 2013). Briefly, 3 mg of HPE were added to 40 mL of methanol/water (50:50, v/v). The mixture was left to stand for 1 h at 25°C, and the supernatant was then collected and stored (aqueous methanol extract). The residual sample was added to 40 mL of acetone/water (70/30, v/v) and treated as described before. The aqueous methanol and acetone extracts were mixed, evaporated and re-dissolved in 10 mL of ethanol (aqueous ethanol extract). Finally, the residual sample was dissolved in 10 mL of H2O (aqueous extract). One mL of each aqueous and ethanol extract was brought to a volume of 2.5 mL using 0.3% HCl. Subsequently, 50 μL of either this sample or the standard quercetin was added to 1 mL of 2% Na2CO3, followed by the addition of 50 μL of Folin-Ciocalteu reagent diluted in water (1:1). After 30 min incubation at 25 °C, the absorbance at 750 nm was recorded using a spectrophotometer (Genesys 10 UV, Thermo Electron Co. USA). The total phenol content was expressed as milligrams of gallic acid equivalent per gram of HPE (GAE/g) using a standard curve (concentrations 0.2 - 1.0 mg/mL) for a freshly prepared solution of gallic acid.
Subjects selection and clinical examination
This study included eight healthy males. Their average age was 32 years old (SD 11.1). Exclusion criteria included that none of them had any chronic disease such as diabetes mellitus, cardiovascular disease, anemia; they were not under drug treatment or having history of alcoholism and/or smoking, and neither excessive exercise, which induce oxidative stress. The study was explained to all subjects, and their consent signature was taken on a voluntary bases. Also, this study was approved by the Institutional Ethical Committee for Human Research, Faculty of Health Sciences and Nutrition in the State University of Tlaxcala. Body mass index (BMI) was obtained by accurately measuring height and weight of each individual (BMI = weight [kg] / height [m2]).
Blood sample collection and preparation
Seven mL of fasting venous blood sample was collected at 10:00 am from each subject under aseptic conditions, in sterile EDTA-vacutainer tubes (Becton Dickinson Co. NJ, USA). RBCs and serum were separated immediately by centrifugation at 3000 g for 10 min at 25 °C. Hematocrit and total hemoglobin were determined in each individual blood sample. After centrifugation, RBCs were washed and centrifuged 3 times in PBS [0.1 M (0.2 g KCl, 0.2 g KH2PO4, 8 g NaCl, and 2.16 g NaHPO4.7H2O)/L), pH 7.4 (with 10% NaOH] and white cells were pipetted out and discharged. RBCs samples were stored in PBS as the initial hematocrit conditions (1:1) and used for all experiments. All samples were prepared and analyzed the same day of collection, unless otherwise specified.
Ghost cells (GC) preparation
Four mL of separate RBCs in PBS were used to prepare hemoglobin-free erythrocyte membranes according to a hypotonic-hypertonic lysis procedure (Matteucci et al., 1995; Krishnaveni et al., 2013; Jangde, 2011).
Lipid peroxidation induced by FeSO4 in RBCs and GC (TBARS)
Antioxidant activity from the HPE was estimated by using the spectrophotometric method described by Ohkawa et al. (1979). Briefly, extract samples at 10, 30, 50, 100, and 1000 mg/L concentration were prepared in order to inhibit lipid peroxidation induced by Fe2+ in RBCs and GC.
Ghost cells and RBCs were washed and homogenized separately in 10 mL of PBS (pH 7.4) at 5°C (1:10 w/v) (Dominguez et al., 2005). The homogenate was centrifuged for 10 min at 3000 g, and the supernatant was adjusted at 2.66 mg of protein/mL in PBS to be used for total protein determination according to the method of Lowry et al. (1951). BSA was used to elaborate the standard curve. Then 375 µL (1 mg protein content) of RBCs or GCs were previously incubated in an orbital shaker with 50 µL of EDTA 10 µM, and 50 µL of each one of the different extract concentrations of HPE at 37 °C for 30 min for a working dilution concentrations of 3-50 mg/L. Lipid peroxidation was initiated by the addition of 50 µL of FeSO4 (final concentration = 25 µM) (Ng et al., 2000). After 1 h, samples were cooled on ice, and 0.5 mL of TBA (1% 2-thiobarbituric acid in 0.05 N NaOH and 30% trichloroacetic acid, 1:1) was added. Samples were centrifuged 10 min at 3000 g. Lipid peroxidation and their final products in RBCs and ghost cells, were assayed by malondialdehyde (MDA) determination using TBARS assay method (Kibanova et al., 2009). Samples were heated at 94 °C for 30 min in a boiling water bath. After cooling on ice, 200 µL of each sample and a blank sample, were placed in an Elisa microplate reader and the absorbance was recorded at 540 nm. TBARS results were expressed as the equivalent of MDA, compared with 1,1,3,3-tetramethoxipropane as the standard. The percentage of inhibition is defined as the decrease of TBARS formation, due to the extract quelation (Esterbauer and Cheeseman, 1990). HPE was evaluated to determine the minimal concentration (μg/mL) to inhibit lipid peroxidation up to 50% (IC50 μg/mL).
Lipid peroxidation induced by FeSO4 in RBCs to morphologic analysis
HPE was diluted at 5, 10, 100, and 1000 mg/L in PBS with the pro-oxidant FeSO4 (100 µM). Then, 25 µL (~ 125 x 106 cells) of RBCs were added in each dilution and incubated at 25 °C during 5, 30, 120, 180 min, 1, 2, 15, 22, 24 and 28 days. The samples without FeSO4 or HPE were prepared as control blanks. Experiments were carried out in triplicate in RBCs.
Cytological analysis
RBCs samples (2 ?L = ~10 x 106 cells) were taken from different incubation times, and they were extended on a glass slide and immediately fixed with 10% formaldehyde buffer. Cells were stained with HE and a glass cover was mounted for microscope analysis at a magnification of 40x, and 100x using an oil immersion objective. An Olympus CH30RF100 microscope (Olympus Optical Co. LTD, Japan) with a digital camera connected at one PC and an image analysis software system (Motic Images Plus 2.0, Motic China Group Co. LTD) was used to quantify static and kinetic RCBs and the morphology changes.
Histometrical measurements
Histometrical measurements were performed at a magnification of 40x and 100x. The examiner was blinded to which group each specimen belonged to. RCBs were counted in three standard areas of each slide (0.1 mm x 0.1 mm each). In each area three different squares of 70 µm were selected to cell counting. Intensity of colour stain (Anisochromy), size (Anisocytic), shape (Poikilocytic), inclusions and other morphological changes in RBCs were determined as described by Mackenzie (2000) and Brecher and Bessis (1972) as follow:
HE Stain: Normochromic
Hypochromic
Hyperchromic
Size: Normocytic (7 – 8 µm)
Microcytic (less than 7 µm)
Macrocytic (more than 8 µm)
Shape: Acanthocyte (crenated)
Echinocyte
Inclusions: Heinz bodies
Others: Hemolysis
Schistocyte
Dynamic flow RBCs assay
The dynamic flow of RBCs was assayed using a flow channel (20 µm wide, 10 µm high, 5 mm long) cut on a glass microscope slide made in the laboratory and covered with a glass-cover slide to permit direct visualization during the entire flow study (Hillery et al., 1996). The experiments were performed at 25°C. A clean glass slide was put with a 5o inclination on the microscope slide to permit continuous and free flow of RBCs and to avoid shear strength. Two microliters of the above mentioned 2 h-incubated RBCs in PBS or HPE, were dropped on one extreme of the chamber and let to flow freely for 120 min. The number of adherent cells, crenation (acanthocytes and echinocytes), and deformability against time, were counted by direct microscopic visualization and a video image system (20 frames per second) providing multidirectional pictures of the erythrocytes. In order to avoid glass near deformation of RBCs or echinocytes formation (Brecher and Bessis, 1972; Bronkhorst et al., 1995), only cells flowing in the middle of the channel were analyzed.
Polyacrylamide gel electrophoresis
Protein determination and standardization of RBCs concentration was determined as described above by the method of Lowry. After washing with PBS, 25 μL of each incubated sample of RBCs were reconstituted under non-denaturing conditions in 20 μL of sample solubilizing buffer (0.125 M Tris-HCl, 4% SDS, 20% glycerol, pH 6.8, 0.002% bromophenol blue) and sonicated for 3 min prior to running the gels. 1 μL of each individual RBC sample (considering that in 1 μL of blood exist ~ 5 x 106 red blood cells) were subjected to SDS-PAGE electrophoresis at room temperature in a discontinuous gel system according to the method described by Laemmli (1970), using a 3% stacking gel and a 10% separating gel. Molecular-weight standards were used in all gels. Electrophoresis was carried out at 100V constant current until the dye front of the gel reached the bottom of the chamber. A minigel apparatus (Mighty Small II vertical slab unit; Hoefer Scientific Ins., San Francisco CA., USA) was used for this purpose.
Gel staining
Gels were fixed and stained successively with 0.1% Coomassie brilliant blue R250 in methanol-water-acetic acid (1:8:1), and silver stain. To enhance and obtain a good pattern of staining bands, gels were initially stained with the solution of Coomassie blue for 1 h and distained gradually with several changes of distain solution (methanol-water-acetic acid, 1:8:1) for approximately 60 min until the bands showed the appropriate stain.
Identification of membrane proteins
Proteins were identified according to their relative mobility in the gel and stain patterns following the criteria described by Ballas (1977), Mackenzie (2000), and Singh and Rajini (2008). Membrane and cytoskeletal molecules were scored according to the size and stain intensity of the bands as: absent (-), present (+), low intensity and size (-+), and high intensity and size (++). However, because of the subjectivity inherent in these parameters, protein bands were also scored only as present or absent.
Statistical analysis
TBARS analysis
Data in the text, tables and figures were expressed as means with standard error of mean (SEM). One-way analysis of variance (ANOVA) was used to compare the antioxidant capacity of HPE in RBCs, GC, and quercetin/GCs.
RBCs counting
All data to count the RBCs were presented as mean and the standard error of the mean (SEM). The significance between RBCs control group and RBCs experimental group with HPE, was determined using two-way ANOVA repeated measures analysis. HPE was considered as the first variable, and the time as the second variable. A subsequent Bonferroni correction test was performed to determine difference between groups and baseline. A P<0•05 was considered to be statistically significant. In all cases, each experiment was run in triplicate within two control samples. The GraphPad Prism version program (v.5.01 for Windows) was used for this purpose.
RESULTS
Total phenolic compounds, free radical capture of DPPH and IC50
A total of 2.2 g of HPE was obtained from 30 g of hawthorn dried skin. The total polyphenol content in HPE was found to be 0.68 (SD 0.001) mg of gallic acid/g. The amount of DPPH reduced was quantified measuring the decrease in absorbance of DPPH by the HPE. The IC50 of the HPE to scavenging free radicals was 15.26 ± 0.20 mg/L as compared with the IC50 of the pure antioxidant quercetin [5.37 (SD 0.14) mg/L]. It was shown that acetone extract is three times less active than quercetin in scavenging DPPH radical.
MDA determination in RBCs and ghost cells by TBARS
Red blood cells have both saturated and unsaturated lipids and fatty acids on its membrane, which are susceptible to be peroxidized by the ROS. Malondi-aldehyde (MDA) is one of the final products of cell membrane lipoperoxidation. In order to determine the antioxidant capacity of hawthorn fruit peel extracts, FeSO4 was used to induce the lipoperoxidation in RBCs and erythrocyte ghost. The ferrous ion (Fe2+) is capable of forming lipid peroxides that induce the formation of highly reactive substances as hydroxyl radicals, perferryl and ferryl species.
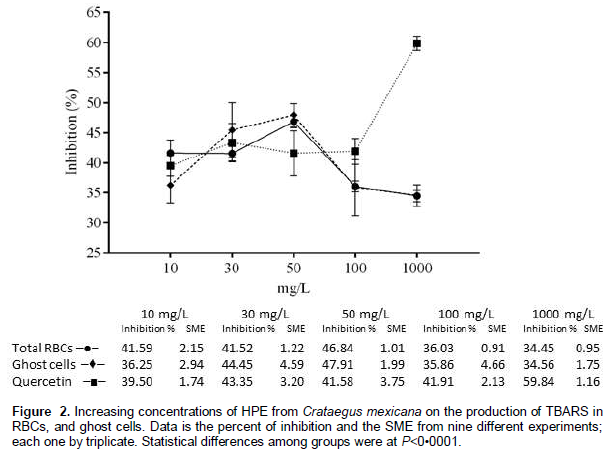
To evaluate the effectiveness of hawthorn extracts to protect the RBCs and ghost cells from the lipoperoxidation, different concentrations of extracts were used as described in material and methods. The acetone HPE had a better performance to inhibit lipid peroxidation in RBCs (46.84%) and in ghost cells (47.91%) in a concentration of 50 mg/L, as compared with GC/quercetin (41.58%) (Figure 2).
RBCs quantitation and morphological changes
The body mass index and RBCs samples from 8 subjects were analyzed to determine their good health status to standardize experimental conditions. Means of BMI was 21.4 kg/m2 (SD 5.2), hematocrit 0.49% (SD 0.05), and hemoglobin 12.5 mg/dL (SD 1.8). All experiments were run immediately after each blood collection.
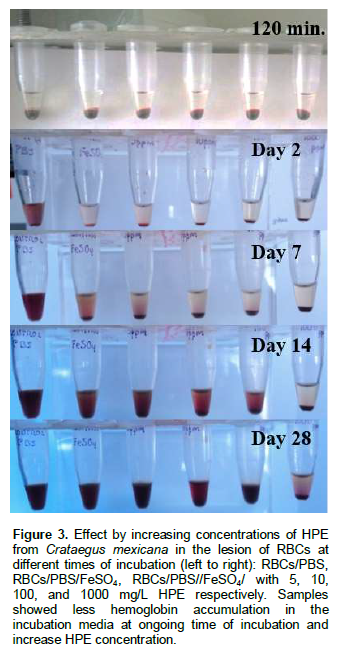
RBCs were incubated in HPE with the prooxidant FeSO4 at different concentrations and analyzed for morphological changes, eryptosis and hemolysis. The extent of hemolysis of RBCs in the different HPE concentrations showed a tim dependent concentration in the erythrocyte damages during the 28 days of incubation. RBCs with higher HPE concentration were less damaged as compared with a lower concentration of the extract as visually demonstrated by the presence of less hemoglobin accumulation in the incubation media (no quantified). This suggests a low-level of hemolysis and damage on the membranes of the erythrocytes, keeping the colour and less cell adhesion in the erythrocyte-clot formation (Figure 3). These observations suggest an antioxidant effect of HPE in RBCs, and are similar to those found by Baek et al. (2012) that described the attenuation of cell lesion using haptoglobin therapy in guinea pigs red blood cells.
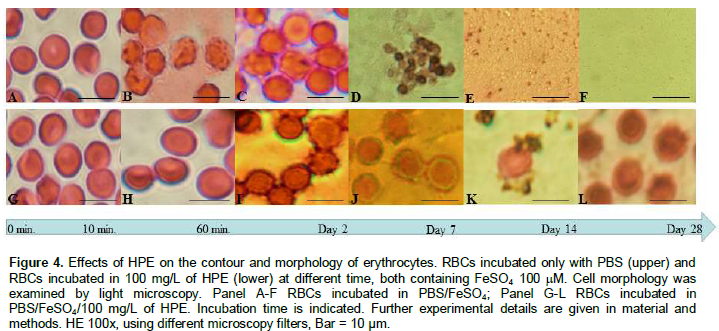
RBCs exposed to the different concentrations of HPE retained in greater or lesser extent their normal morphology and phenotypic characteristics during all the time of the experimental conditions. A concentration of 100 mg/L showed a better and repeatability pattern under the microscopic analysis. The hemolysis process was faster in untreated RBCs (Figure 4A to F) and started at day 7 of incubation, as compared with the RBCs treated with HPE (Figure 4G to L), where the hemolysis process started later. All morphological cell changes are described independently and also the sum of these changes.
Normocytic and normochromic erythrocytes
Microscopic counting of RBCs was similar in both cases, showing normocytic cells at the beginning of the experiment incubated a non-incubated with the HPE. The counting of cells was easier in treated RBCs than in untreated RBCs, because the cytological smear in untreated RBCs showed large amounts of aggregated cells that was made difficult to separate each cell from the other.
Smear from treated RBCs with HPE was homogeneous and the cells showed a better separation among them; this could be due to the antioxidant activity since the first moment that the cells were incubated in the extract. Treated RBCs were longer hydrated, that suggests no loss of water inside cells by keeping the cell membrane intact for longer periods of time. Evident morphologic changes and hemolysis of RBCs began at 180 min either treated and untreated erythrocytes with HPE, and hemolysis started at 48 h in untreated RBCs (Figure 5a). By contrast, the beginning of hemolysis in RBCs treated with HPE started later than in untreated cells, maintaining the morphology for 28 days (Figure 4K to L). Total RBCs quantitation showed the effect of HPE (P<0•01), during time of incubation in min and days (P<0•001). A comparable dose-response relationship was observed in RBCs at different time points of incubation.
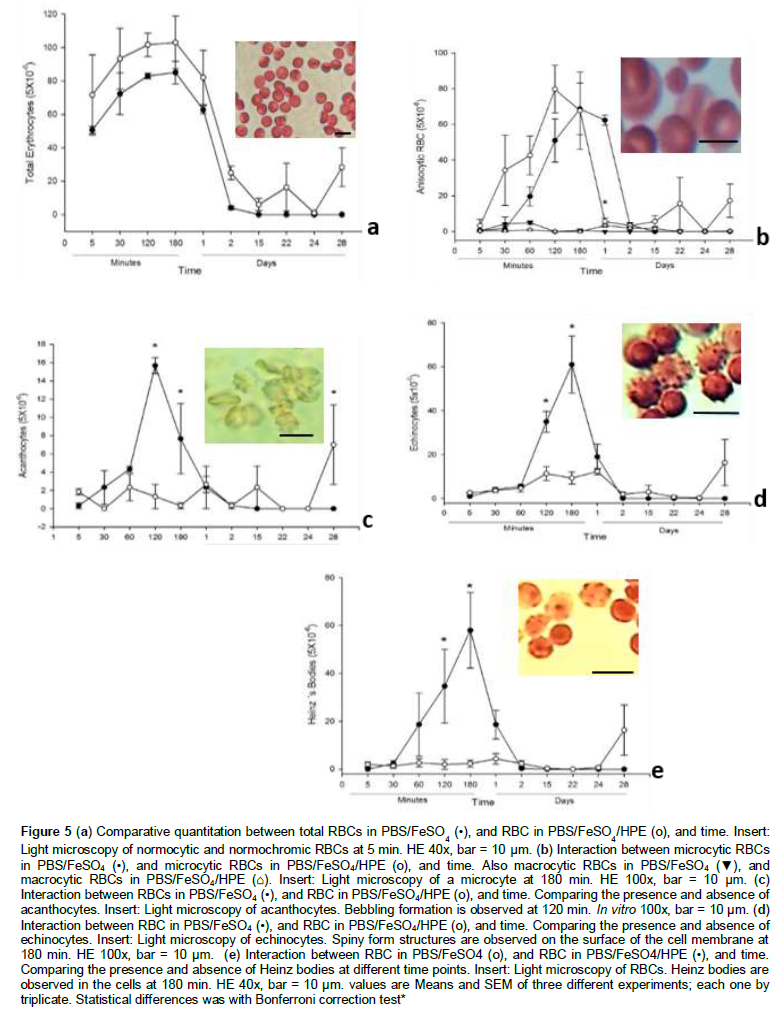
Anisocytic RBCs
An increased number of microcytic erythrocytes was observed during the first 120 min in both incubated PBS/FeSO4 cells and in PBS/FeSO4/HPE cells. Following this microcytic formation, a decrease in these cell number was observed until they became lysed. However, the cells treated with the HPE remains up to for 28 days. This suggests that the crenated cell formation and the eryptosis process is retarded with the HPE extract (Figure 5b). No effect in the formation of microcytic cells was observed by the treatment (P<0•05), however the time of incubation (P<0•0001) and interaction (P<0•005) induce microcytic formation.
Crenated, acanthocytes
The oxidation of the main structural proteins spectrin and actin that form the submembrane cytoskeletal meshwork of RBCs, induced morphological and viscoelastic changes in the cell membrane that were observed as blebs formation on the surface of the cells (Figure 4B to C, and 5c). RBCs with the HPE showed an early inhibition in the formation of these continuous bebbling oxidative phenomenon, thus also inhibited the eryptosis and hemolysis process in the life span of the erythrocyte (Figure 4I to L, and 5c) as compared with the RBCs from the control group. These data suggest that the acetone extract is acting to reduce membrane cytoskeletal deformation caused by the FeSO4 oxidative effect on the lipoperoxidation of phospholipid on the membrane bilayer, and besides inhibit the oxidation of amino acidresidue side chains, formation of protein-protein cross-link, and oxidation of the cytoskeletal backbone of proteins in the erythrocyte. These morphological changes observed during the first 24 h reached the maximun peak at 120 min of the untreated red blood cells. A less formation of crenated cells is related to the HPE (P<0•01), time (P<0•0001), and interaction factors (P<0•0001). These morphological changes have been described as Type I and II (Brecher and Bessis, 1972) in the eryptosis process of the RBCs. As we already mentioned, HPE-treated RBCs did not increase these changes in a time-dependent manner.
Crenated, echinocytes
As the time increased the morphology changes in RBCs became more evident. The oxidative process and the crenation in RBCs induced the formation of cytoplasmic prolongations due the membrane deformation by cytoskeletal protein and lipid oxidation. These changes became non-reversible and were determinant in the eryptosis process of RBCs (Figure 4C, and 5d). The HPE from C. mexicana inhibited echinocytes formation following the time point in the acanthocytes formation, retarding cell deformation and the hemolysis process (Figure 4I – K, and 5d). A time dependent action in echinocyte formation (P<0•0001) and its interaction with the HPE (P<0•01) was observed. This morphology change in the cell membrane of the erythrocyte has been denominated as Type III (Brecher and Bessis, 1972) with many different forms in membrane deformation, and is considered as a non-reversible morphological alteration during the life span of the erythrocyte and therefore in the eryptosis process. The biological process in acanthocytes formation by radical-mediated damage in proteins and membrane, is similar in echynocites when they became FeSO4 oxidized.
Heinz bodies
The oxidation of hemoglobin produced in their primary structure disulphide cross-links between adjacent globin chains which become distorted and eventually lead to visible precipitates in the inner membrane of RBCs. HPE was able to inhibit the formation of these bodies from the beginning of in vitro experiments up to the day 24. RBCs without the acetone extract initiated the formation of Heinz bodies at min 30, increasing the formation in the next min and days. The increase in hemoglobin precipitation was associated by the distortion of the cell at the final phase of acanthocytes formation during the eryptosis process. This data suggest that HPE inhibited the oxidative process of hemoglobin for a longer period of time (P<0•0001), this interaction permits the extension of the life span of the cell (P<0•0001). Afterwards, the hemolysis process continued and began the formation of schistocytes. This changes last longer than the control group in which the Heinz body formation and hemolysis took less time (Figure 4B, 4I, and 5e).
Schistocyte
The schistocytes formation was observed on the first day of incubation, this is due to the deformation, bebbling formation and separation of liposomes from the surface of the cell membrane to be completely degraded (Figure 4D to E). However, schistocytes were observed in the RBCs incubated in HPE up to for 28 days, probably because the cells keeps their membrane for longer time as is deduced observing all the graphics in the different experiments (Figure 4K).
Dynamic flow of RBCs assay
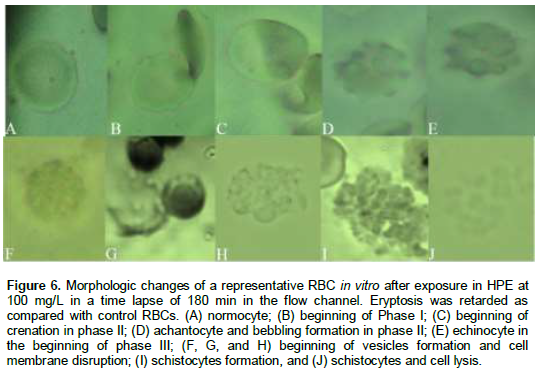
To critically evaluate the potential contribution of HPE to inhibit any cellular morphologic change by protein and lipid membrane oxidation, RBCs flow rate, adhesion, and morphologic changes were quantified from video recording of nine individual cells in the flow channel over a period of 180 min. RBCs incubated in HPE demonstrated a longer period of time of flowing through the flow channel up to for 180 min as compared with RBCs in PBS whose cells were dehydrated in less than 6 min. Morphologic changes such as acanthocytes and echinocytes formation were observed, and an increase of adhesion among cells was demonstrated in RBCs incubated only in PBS during the first 60 min as compared with RBCs incubated with the HPE (Figure 6A to J) without changes in the morphology up to 120 min. It is particularly notable in these experiments and suggest that RBCs maintain their morphologic and the micro-rheologic characteristics in the micromolar concentrations of HPE. This could be because there is not lipid or protein oxidation in the cell membrane, thus no dehydration and crenation, and therefore inhibition of cell deformity and viscosity and better flow of RBCs. However, life span of RBCs with or without the extract is going to continue its eryptosis process, thus the cell will lose their biological properties and finally will be cell lysed.
Polyacrylamide gel electrophoresis
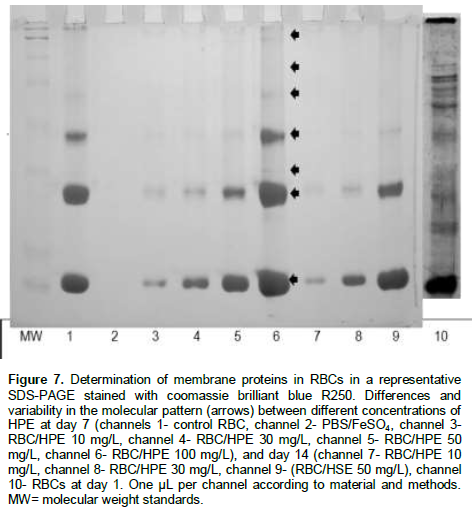
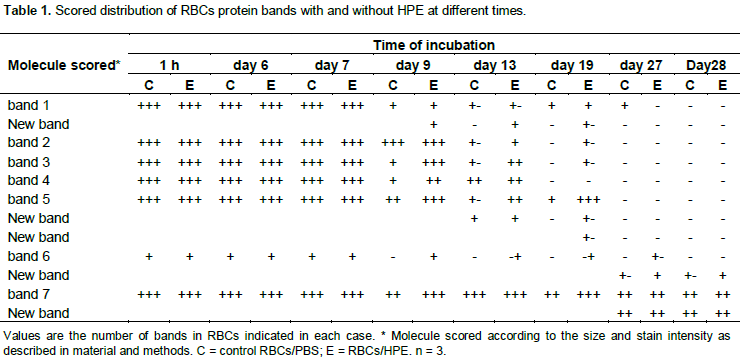
The banding pattern of some cytoskeletal proteins of RBCs were stained and identified in SDS-PAGE. Band staining intensity was observed during almost all time points of incubation of RBCs with the different concentrations of the hawthorn extract.
The qualitative analysis showed that some bands keep the stain intensity in relation to a major concentration of the HPE. In these experiment, seven main bands were observed at day 1 of the experiments (Figure 7). The colour intensity and size of some bands were maintained over a longer period of time with increasing the extract concentration and suggest an antioxidant protective effect of the HPE on the cytoskeletal proteins of the RBCs. This data is related to what we observed in the course of incubation, where the higher is the concentration, the lesser is the lesion in RBCs. This supports the fact that the cytoskeleton is protected against the oxidative process induced by the FeSO4, therefore the cells will keep their morphology and rheology characteristics over a longer period of time.
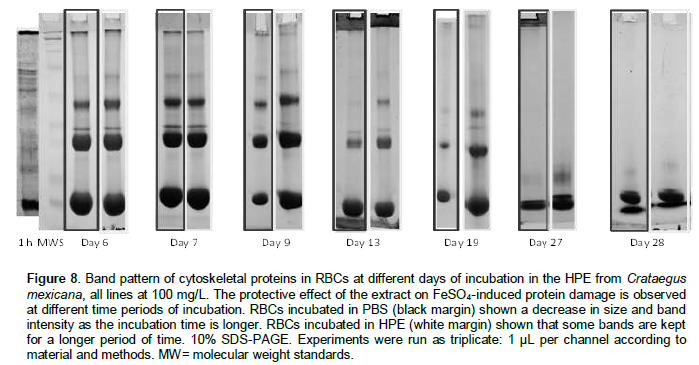
The banding pattern was compared between RBCs in PBS, and RBCs in HPE in an attempt to determine time-dependent antioxidant protective effect on the cytoskeleton proteins. Meanwhile the banding pattern of some protein bands were observed in some cases up to the day 28 when the RBCs are incubated with HPE, the proteins in the control group were degraded, and some of them in a time point of incubation were absent (Figure 8). Because the band pattern was constant in each different concentration, it has been only shown the band pattern with HPE in a concentration at 100 mg/L. These observations were scored (Table 1).
Singh and Rajini (2008), in a similar experiment showed the antioxidant protective activity against ferrous- ascorbate using potato peel extract in different concentrations, but they did not determine time dependent changes during long periods of time. This makes clear that incubating RBCs in extracts containing polyphenols improve the antioxidant effect on the cytoskeletal proteins. Many pro-oxidants such as H2O2, Fe2+-ascorbate and AAPH are capable to induce alterations in the erythrocyte membrane and a decrease in cytoskeletal proteins that lead a series of changes in the morphology and rheology properties of the cells (Singh and Rajini, 2008; Battistelli et al., 2005; Srour et al., 2000). In these experiments, the use of the pro-oxidant FeSO4 is responsible of the oxidative attack inducing this changes and subsequent non-enzymatic protein fragmentation and membrane deformation, but the HPE of C. mexicana is capable of protecting the RBCs of the oxidative process reducing the morphologic changes and retarding the hemolysis in the eryptosis process.
DISCUSSION
The most abundant cells in the blood are the RBCs with a 45% of its total volume, with an average life span of 120 days, and upon which main function is transportation of O2 and CO2. During their life span RBCs are exposed to many stressful situations through the capillaries, and suffering osmotic, energetic, and oxidative stress. Therefore, the high concentration of oxygen and haemoglobin promote the oxidative process and impaired RBCs functions and damage in the cell. The oxidative stress process is catalysed by Fe2+, via the Fenton reaction to produce hydroxyl-radical; a process that has been well documented (Cheng and Li, 2007). The oxidation of the polypeptide backbone is O2.- dependent; is caused by the ?OH-dependent abstraction of the α-hydrogen atom of an amino acid residue, this results in the formation of a carbon-centered radical that react with O2 in a series of molecular reactions, forming alkoxyl radicals and aldehydes, the last one may participate in Schiff-base formation and Michael addition (Dean et al., 1997). Many of the steps in this pathway that are mediated by interactions with HO2? can also be catalysed also by Fe2+. The intermediate radicals may undergo side chain reactions with other amino acid residues in the same or a different protein molecule to generate a new carbon-centered radical and the cleavage of the peptide bond of proteins. In addition, carbonyl groups may be introduced into proteins by reactions with aldehydes (MDA) produced during lipid peroxidation (Figure 9).
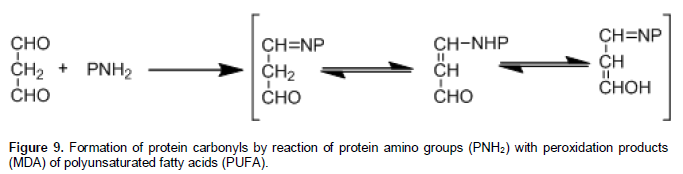
This carbonyl groups and MDA are associated with aging, oxidative stress, many chronic diseases, and human red blood cells (Berlett and Stadtman, 1997). Therefore, these oxidized products in the cell proteins of RBCs, can contribute in many morphologic changes and reduced life span of the erythrocyte. An increase in oxidation raises hydrophobicity, while a limited oxidation increases hydrophilicity. This change in turn produces inner and outer rheology alterations on the cell bilayer membrane, and such responses will further heighten the eryptosis process in RBCs.
Hawthorn is a source of natural polyphenol antioxidants (García-Mateos et al., 2012). Acetone extract from hawthorn C. mexicana inhibited oxidative stress induced by the pro-oxidant FeSO4 in RBCs. It was demonstrated and confirmed by TBARS analysis, evidences in cell morphology, and protein electrophoretic pattern using different concentrations. Our results by TBARS analysis showed a relationship between low doses of HPE to inhibit MDA formation induced by FeSO4 in the RBCs. These results are similar to those demonstrated in previous studies using homogenate from rat brain cells.
FeSO4-induced lesion of the cell membrane; hemolysis and morphologic changes in the RBCs were inhibited in a concentration-dependent manner in the presence of HPE, and the life span of the erythrocytes were extended in the RBCs samples that contained the acetone extract. The morphologic changes associated to the eryptosis process were retarded during the time of the experiments. It was observed that the crenated cells, acan-thocytes, and echinocyte were present for longer periods of time, before the hemolysis process and/or schistocyte formation took place. This protection supplied by the hawthorn extract was observed during the first 24 h of the experiment, as compared with the control group, where the eryptosis and life span of RBCs are reduced in a significant period of time. Zhu et al. (2002), determined the antioxidant protective effect of the cocoa feeding in rats, to inhibit erythrocyte hemolysis up to for 240 min in a dose dependent manner, suggesting that some poly-phenols in HPE have the capacity of quenching free radicals in the cell membrane of the erythrocyte. Our study also showed a longer period of time on the inhibition of FeSO4-induced hemolysis and morphologic changes in the erythrocytes, in a concentration-dependent manner up to for 28 days. This results support the fact that polyphenols in hawthorn fruits improve the antioxidant protective effects in RBCs, and may be working in an interactive way on their lipid bilayer of the cells.
Changes in the number, relative mobility and staining intensity of the protein bands observed in the electrop-horetic gels, indicate that cytoskeletal proteins are susceptible to oxidation due the pro-oxidant FeSO4 and during life span of the erythrocyte under the experimental condition. Furthermore, RBCs samples with the different hawthorn extract concentrations showed that many proteins bands are maintained during different times of the experiment and, also with a higher concentration of the extract the band pattern staining of some proteins was more intense. This may indicate that the extract contribute in some way in the inhibition of the protein oxidation for a longer and extended period of time, probably by protecting the –SH protein groups against oxidation. Also, it can inhibit the ability of proteases to degrade the oxidized forms of other proteins, and therefore to increase cellular resistance to oxidative stress (Asgary, et al., 2005). The delay time for protein oxidation and polymerization has a relationship on the stability and morphology of the erythrocytes. The lipid bilayer membrane and protein stability should be related to antioxidants concentration present in the HPE of C. mexicana, reducing cell dehydration, K loss and other osmotic cell changes (Wandersee et al., 2005). This protection in the cytoskeletal proteins is related to better form and shape of the erythrocyte as compared with those RBCs without the extract of C. mexicana. The observed reduction in the number and intensity of the band pattern could be related to increased proteolysis or instability of the absent bands, but is also unlikely in view of the finding that results were similar in all the experiments in RBCs comparing with or without the acetone extract. The pathophysiology of this changes, remains to be determined using specific and purified antioxidants.
The deformability of the red blood cells is an important factor in the rheology of the blood (Bronkhorst et al., 1995; Dupire et al., 2012), and it is related to many important cell properties, flow rate and blood diseases. The deformability is the result of several mechanical and geometrical properties of the RBCs, and is determinant in the stability of the membrane lipids and cytoskeletal proteins of the cell. Consequent disruption of the membrane-cytoskeleton by any oxidative process may be responsible for the bebbling formation in RBCs (Spangler, 2011; Liu et al., 2012). Thus, hydrodynamic stress and oxidation of proteins and phospholipids can also induce morphological and rheological changes of the cell. HPE of C. mexicana, was able to inhibit oxidative stress maintaining the structural and morphologic integrity of the cell, with less changes and damage in the morphology of the erythrocyte under the experimental conditions. HPE reduce the eryptosis process, retarding the formation of crenated cells and increasing the flow rate of the erythrocyte, contributing to the cell motion and avoid energetically a costly oxidation and deformation, to keep the cell elasticity and shape. This could be possible by the presence of antioxidants metabolites in the acetone HPE. This capacity of the hawthorn may be used to avoid erythrocyte adhesion and vasoocclusion in the blood stream in many blood diseases, and to inhibit peroxidation of biomembranes that can induce damage during ischemia/reperfusion, inflammation and aging.
In spite of this, is well known that echinocytes are formed after 3 to 4 days in whole blood kept at 4o C and within 24 h in blood incubated at 37o C (Brecher and Bessis, 1972), but with the acetone extract of C. mexicana the crenated cells, acanthocytes and echinocytes, were formed later compared with the control group and the shape of the cell membrane was kept for 28 days, showing inhibition during the time of the experiment of these morphological changes. Thus, may be, the oxidative process in the cells is inhibited by the antioxidants in the extracts, that maintain the morphology of the cell and its life span for a longer time, reducing the eryptosis process.
The attenuation of RBCs lesion was demonstrated in our study, and this is similar to those reported by Baek et al. (2012) in guinea pigs by haptoglobin therapy. However, the hawthorn extract is a natural product than can be consumed regularly by the human being in healthy and disease conditions. Wautier et al. (1981) mentioned the increased adhesion of erythrocytes to endothelial cells in diabetes mellitus and its relation to vascular complications, therefore, the nutraceutical application of hawthorn could inhibit the adhesion by keeping the RBC membrane from being oxidized by free radicals.
These results are in broad agreement with the published reports regarding anti-oxidant capacity of other fruits and vegetables extracts, such as potato peel extracts, cocoa extracts, and flaxseed oil. A large body of evidence implicates oxidative stress in the pathogenesis of red blood cell diseases, affecting their life span, shortening the eryptosis process, and increasing the hemolysis process. This process might also result in a systemic inflammatory response. Therefore, we favor the hypothesis that the observed protective effects on the morphology and life span of RBCs is due to the presence and collective action of the antioxidant molecules present in the acetone HPE. These results are related to the use of hawthorn in Mexico and other countries since ancient times in order to treat blood and other related systemic diseases. However many of this complex dynamics and interactions remain unexplored experimentally.
There is a need to identify more effective and novel compounds of fruits according to food consumption in traditional diets like the Mexican one, as well as how theirs constituents are acting against oxidative stress in cells, tissues and, in human diseases. The broad range of fruits and theirs parts to inhibit and remove ROS and intermediate radicals in many forms encourage continuing the purification of antioxidants in Mexican fruits and determining their mechanisms of action in lipids and proteins in red blood cells.
CONCLUSIONS
This study provides the first in vitro evidence for an active role of C. mexicana as antioxidant on erythrocytes. Hawthorn is a fruit with anti-oxidant potential, reduces morphological changes and extends life span of the red blood cells in vitro. Such protection could be due to the inhibition of membrane lipoperoxidation and oxidative damage in cytoskeletal proteins. The conjunction of antioxidants in this extract, and not just specific purified antioxidants, could be helpful to keep normal the rheology properties of the erythrocyte apparently without any pro-oxidative damage.
The antioxidants present in the acetone extract may contribute to protect the erythrocyte cell membrane against lipid peroxidation induced by oxidative stress, thereby participating in diminishing the morphological and many rheology changes that may happen in the pathogenesis of vasoocclusive diseases, such diabetes mellitus, metabolic syndrome, different types of anemia, and the inhibition of cell storage lesion in RBCs.
More studies are required to correctly identify the bioactive compounds of C. mexicana and how it contribute to the erythrocyte protection, and their mechanisms of action in the protection of the RBCs. This study give the scientific biochemical bases to use hawthorn extracts in a clinical setting, to prevent or treat many diseases and conditions associated to oxidative stress.
CONFLICT OF INTERESTS
The authors did not declare any conflict of interest.
ACKNOWLEDGMENTS
This study was supported by grant 105663 from the Mexican Council of Science and Technology (CONACYT). We also acknowledge CONACYT for providing the financial support (54150) to José Antonio Banderas Tarabay during his Ph.D. studies.
REFERENCES
| Aebi H, Bossi E, Cantz M, Matsubara S, Suter H (1968). A catalasemia in Switzerland. In: Hereditary Disorders of Erythrocyte Metabolism. Beutler E, editor. pp. 41-65. | ||||
|
Andrade-Cetto A, Heinrich M (2005). Mexican plants with hypoglycaemic effect used in the treatment of diabetes. J Ethnopharmacol. 99: 325-348. Crossref |
||||
|
Arrieta JD, Siles-Barrios J, García-Sánchez B, Reyes-Trejo and Sánchez-Mendoza ME (2010). Relaxant effect of the extracts of Crataeugus mexicana on guinea pig tracheal smooth muscle. Pharmacogn. J. 2: 40-46. Crossref |
||||
|
Asgary S, Naderi GH, Askari N (2005). Protective effect of flavonoids against red blood cell hemolysis by free radicals. Exp. Clin. Cardiol. 10: 88-90. PMid:19641665 PMCid:PMC2716227 |
||||
|
Baek JH, D’Agnillo F, Vallelian F, Pereira CP, Williams MC, Jia Y, Schaer DJ, Buehler PW (2012). Hemoglobin-driven pathophysiology is an in vivo consequence of the red blood cell storage lesion that can be attenuated in guinea pigs by haptoglobin therapy. J. Clin. Invest. 122:1444-1458. Crossref |
||||
| Ballas SK (1977). Abnormal erythrocyte membrane protein pattern in severe megaloblastic anemia. Clin. Res. 25: 333A. | ||||
|
Battistelli M, De Sanctis R, De Bellis R, Cucchiarini L, Dachá M, Goobi P (2005). Rhodiola rosea as antioxidant in red blood cells: ultra-estructural and hemolytical behaviour. Eur. J. Histochem. 49:243-254. PMid:16216810 |
||||
|
Berlett BS, Stadtman ER (1997). Protein oxidation in aging, disease, and oxidative stress. J. Biol. Chem. 272: 20313-20316. Crossref |
||||
|
Brecher G, Bessis M (1972). Present status of speculated red cells and their relationship to the discocyte-echinocyte transformation. A critical review. Blood 40:333-343. PMid:5056966 |
||||
|
Bronkhorst PJH, Streekstra GJ, Grimbergen J, Nijhof EJ, Sixma JJ, Brakenhoff GJ (1995). A new method to study shape recovery of red blood cells using multiple optical trapping. Biophys. J. 69: 1666-1673. Crossref |
||||
|
Cai Y, Luo Q, Sun M, Corke H (2004). Antioxidant activity and phenolic compounds of 112 traditional Chinese medicinal plants associated with anticancer. Life Sci. 74:2157-2184. Crossref |
||||
| Ca-as P, Olivares M, Celedón C, Llaguno S, Valenzuela A (1988). Actividad del sistema de defensa antioxidante y agresión oxidativa en eritrocitos de recién nacidos de bajo de peso y distintas edades gestacionales. Rev. Chil. Pediatr. 59:83-88. | ||||
|
Cheng Z, Li Y (2007). What is responsive for the initiating chemistry of iron-mediated lipid peroxidation: An update. Chem. Rev. 107: 748-746. Crossref |
||||
| Dean RT, Fu S, Stocker R, Davies MJ (1997). Biochemistry and pathology of radical-mediated protein oxidation. Biochem. J. 324:1-18. | ||||
|
Domínguez M, Nieto A, Marin JC, Jeffery E, Céspedes CL (2005). Antioxidant activities of extracts from Barkleyanthus salicifolius (Asteraceae) and Penstemon gentianoides (Scrophulariaceae). J. Agric. Food Chem. 53: 5889-5895. Crossref |
||||
|
Dupire J, Socol M, Viallat A (2012). Full dynamics of a red blood cell in shear flow. PNAS 109: 20808-20813. Crossref |
||||
|
Edwards JE, Brown PN, Talent N, Dickinson TA, Shipley PR (2012). A review of the chemistry of the genus Crataegus. Phytochemistry 79: 5-26. Crossref |
||||
|
Esterbauer H, Cheeseman KH (1990). Determination of aldehydic lipid peroxidation products. Methods Enzymol. 186: 407-421. Crossref |
||||
| Folin C, Ciocalteau V (1927). Tyrosine and trypthopan determination in proteins. J. Biol. Chem. 73: 627-650. | ||||
| García-Mateos R, Aguilar-Santelises L, Soto-Hernández M, Nieto-Angel R, Kite G (2012). Total phenolic compounds, flavonoids and antioxidant activity in the flowers of Crataegus spp. from Mexico. Agrociencia 46: 651-662. | ||||
| Hebbel RP, Leung A, Mohandas N (1990). Oxidation-induced changes in microrhelogic properties of the red blood cell membrane. Blood 76: 1015-1020. | ||||
| Hillery CA, Du MC, Montgomery RR, Scott JP (1996). Increased adhesion of erythrocytes to components of the extracellular matrix. Isolation and characterization of a red blood cell lipid that binds thrombospondin and laminin. Blood 87 (11): 4879-4886. | ||||
| Jangde R (2011). An Overview of resealed erythrocyte for cancer therapy. Asian J. Res. Pharm. Sci. 1: 83-92. | ||||
| Kempe DS, Lang PA, Duranton C, Akel A, Lang KS, Huber SM, Wieder T, Lang F (2006). Enhanced programmed cell death of iron-deficient erythrocytes. FASEB J. 20: 368-370. | ||||
|
Khattak KF, Simpson TJ, Ihasnullah (2008). Effect of gamma irradiation on the extraction yield, total phenolic content and free radical-scavenging activity of Nigella staiva seed. Food Chem. 110: 967-972. Crossref |
||||
|
Kibanova D, Nieto-Camacho A, Cervini-Silva J (2009). Lipid peroxidation induced by expandable clay minerals. Environ. Sci. Technol. 43: 7550-7555. Crossref |
||||
|
Kim HJ, Chen F, Wang X, Chung YH, Jin Z (2005). Evaluation of antioxidant activity of vetiver (Vetiveria zizanioides L) oil and identification of its antioxidant constituents. J. Agric. Food Chem. 53: 7691-7695. Crossref |
||||
|
Kowalczyk E, Kowalski J, Blaszczyk J, GwoździÅ„ski L, Cieéwierz J, Sienkiewicz M (2012). Estimation of cell membrane properties and erythrocyte red-ox balance in patients with metabolic syndrome. Mol.Biol. Rep. 39: 11113-11118. Crossref |
||||
| Krishnaveni G, Krishnamoorthy B, Muthukumaran M (2013). Future Prospectus in Targetting Drug Delivery: A Review on Resealed Erytrocytes - A Promising Carrier. Schol. Acad. J. Pharm. 2: 81-88. | ||||
|
Laemmli UK (1970). Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 277: 680-688. Crossref |
||||
|
Lam RYY, MPhil, Woo AYH, Leung Po-Sing, Cheng CHK (2007). Antioxidant actions of phenolic compounds found in dietary plants on low-density lipoprotein and erythrocytes in vitro. J. Am. Coll. Nutr. 26: 233-242. Crossref |
||||
|
Lang F, Lang KS, Lang PA, Huber SM, Wieder T (2006). Mechanisms and significance of eryptosis. Antioxid. Redox Signal. 8: 1183-1192. Crossref |
||||
|
Liu L, Yang W, Fu J, Yu M, Huang Q, Wang D, Xu J, Chun D, Yao P, Huang F (2012). Effects of flaxseed oil on anti-oxidative system and membrane deformation of human peripheral blood erythrocytes in high glucose level. Lipids Health Dis. 11: 88. Crossref |
||||
| Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951). Protein measurement with the Folin phenol reagent. J. Biol. Chem. 193: 265-275. | ||||
| Lynch RE, Lee GR, Cartwright GE (1976). Inhibition of superoxide dismutase of methemoglobin formation from oxyhemoglobin. J. Biol. Chem. 251: 1015-1018. | ||||
| Mackenzie SB (2000). El eritrocito. En: Hematología Clínica (2nd ed.) México: Ed. El Manual Moderno. pp. 39-60. | ||||
| Matteucci E, Cocci F, Pellegrim L, Gregori G, Giampietro O (1995). Measurement of ATPase in red cells: setting up and validation of a highly reproducible method. Enzyme Protein 48: 105-119. | ||||
|
Méndez-Iturbide D, Banderas-Tarabay JA, Nieto-Camacho A, Rojas-Chávez A, García-Meza MG (2013). Antioxidant capacity of extracts from hawthorn (Crtaegus mexicana) skin. Afr. J. Food Sci. 7:150-158. Crossref |
||||
| Middleton IJ, Kandaswami C, Theoharides TC (2000). The effects of plant flavonoids on mammalian cells: Implications for inflammation, heart disease, and cancer. Pharmacol. Rev. 52: 673-751. | ||||
|
Nagababu E, Gulyani S, Earley CJ, Cutler RG, Mattson MP, Rifkind JM (2008). Iron-deficiency anemia enhances red blood cell oxidative strees. Free. Radic. Res. 42:824–829. Crossref |
||||
|
Ng TB, Liu F, Wang ZT (2000). Antioxidative activity of natural products from plants. Life Sci. 66: 709-723. Crossref |
||||
| Nugraheni M, Santoso U, Suparmo, Wuryastuti H (2011). In vitro antioxidant, antiproliferative and apoptosis effect of Coleus tuberosa L. Afr. J. Food Sci. 5: 232-241. | ||||
|
Ohkawa H, Ohishi N, Yagi K (1979). Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 95: 351-358. Crossref |
||||
|
Pandey KB, Rizvi SI (2011). Biomarkers of oxidative stress in red blood cells. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czech. Repub. 155:131-136. Crossref |
||||
| Salawu SO, Akindahunsi AA, Sanni DM, Decorti G, Cvorovic J, Tramer F, Passamonti S, Mulinacci N (2011). Cellular antioxidant activities and cytotoxic properties of ethanolic extracts of four tropical green leafy vegetables. Afr. J. Food Sci. 5: 267-275. | ||||
|
Singh N, Rajini PS (2008). Antioxidant-mediated protective effect of potato peel extract in erythrocytes against oxidative damage. Chem. Biol. Interact. 173:97-104. Crossref |
||||
|
Sorata Y, Takahama U, Kimura M (1984). Protective effect of quercetin and rutin on photosensitized lysis of human erythrocytes in the presence of hematoporphyrin. Biochim. Biophys. Acta 799: 313-317. Crossref |
||||
| Spangler EJ (2011). Computer simulation of blebbing in erythrocyte membranes. Proccedings of the National Conference on Undergraduate Research (NCUR). Ithaca College, New York. | ||||
|
Srour T, Bilto YY, Juma M, Irhimeh MR (2000). Exposure of human erythrocytes to oxygen radicals causes loss of deformability, increased osmotic fragility, lipid peroxidation and protein degradation. Clin. Hemorrheol. Microcirc. 23:13-21. PMid:11214709 |
||||
|
Van Asbeck BS, Hoidal J, Vercellotti GM, Schwartz BA, Moldow CF, Jacob HS (1985). Protection against lethal hyperoxia by tracheal insufflation of erythrocytes: Role of red cell glutathione. Science 227: 756-759. Crossref |
||||
|
Wandersee NJ, Punzalan RC, Rettig MP, Kennedy MD, Pajewski NM, Sabina RL, Scott JP, Low PS, Hillery CA (2005). Erythrocyte adhesion is modified by alterations in cellular tonicity and volume. Br. J. Haematol. 131: 366-377. Crossref |
||||
|
Wautier JL, Paton RC, Wautier MP, Pintigny D, Abadie E, Passa P, Caen JP (1981). Increased adhesion of erythrocytes to endothelial cells in diabetes mellitus and its relation to vascular complications. N. Engl. J. Med. 305:237. Crossref |
||||
|
Willis MS, Wians FH (2003). The role of nutrition in preventing prostate cancer: a review of the proposed mechanism of action of various dietary substances. Clin. Chim. Acta 330:57-83. Crossref |
||||
| Zhu QY, Holt RR, Lazarus SA, Orozco TJ, Keen CL (2002). Inhibitory effects of cocoa flavanols and procyanidin oligomers on free radical-induced erythrocyte hemolysis. Exp. Biol. Med. 227: 321-329. | ||||
|
Zhu QY, Schramm DD, Gross HB, Holt RR, Kim SH, Yamaguchi T, Kwik-Uribe CL, Keen CL (2005). Influence of cocoa flavanols and procyanidins on free radical-induced human erythrocyte hemolysis. Clin. Dev. Immunol. 12:27-33. Crossref |
||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


