The primary objective of this study was to investigate the involvement of reactive oxygen species (ROS) in the induction of Maillard reaction in the presence of medium-chain triacylglycerols (MCTs). Light irradiation was found to induce Maillard reaction in a glucose-lysine system and led to the generation of ROS. The glucose-lysine Millard reaction was clearly inhibited by the presence of MCTs. Enhancement of this inhibitory effect by the addition of superoxide dismutase (SOD) confirmed that ROS are generated by light irradiation and that the generated superoxide anions (O2-) and hydroxy radicals (•OH) promote induction of the Maillard reaction. It is, hereby, proposed that the primary mechanism by which ROS contribute to the induction of the Maillard reaction involves the generation of •OH through the reaction of O2- and H2O2 (Haber-Weiss reaction).
Medium-chain triacylglycerols (MCTs), comprising only medium-chain fatty acids (containing 8 and 10 carbon atoms), were first used in the 1950s as a dietary therapy for malabsorption syndrome caused by rapid absorption. MCTs have since been extensively studied with numerous excellent reports published. The vast majority of these studies, however, were conducted from a clinical nutritional or biochemical perspective while very few were carried out from the standpoint of food science (Kritchevsky and Tepper, 1965; Leveile et al., 1967; Lavau and Hashim, 1978; Gelebter et al., 1983; Seaton at al., 1986; Chanez et al., 1991; Ecelbarger et al., 1991; Kris-Etherton and Yu, 1997; Papamandjaris et al., 1998). The Maillard reaction is a chemical reaction between amino acids and reducing sugars that results in browning of food. It occurs during bread baking and substantially imparts the external appearance of baked confectioneries (Chi-Hao et al., 2011; Boekel van MAJS, 2006; Tareke et al., 2002).
Building on previous work done and confirmation that MCTs inhibit the induction of the Maillard reaction during bread baking (Toyosaki, 2018), the relationship between the constituent fatty acids of the MCTs and Maillard reaction using a variety of fatty acids was investigated and it was found that induction of the Maillard reaction varies depending on the composition of the fatty acids present (differences in degree of unsaturation and carbon chain length). As these results are related to the induction of the Maillard reaction during bread baking and given the complexity of the reactions involved, it was concluded that the observed differences could not be attributed solely to differences in fatty acid composition. In this study, therefore, a glucose-lysine model was used as a simplified reaction system, to investigate the induction of Maillard reaction by light irradiation Even though it has long been known that Maillard reaction is induced by increased temperature, this study has been able to demonstrate, possibly for the first time, that Maillard reaction can also be induced by light irradiation. The main objective of this study, therefore, was to investigate in detail the relationship between fatty acids (differences in degree of unsaturation and chain length) and the induction of Maillard reaction by light irradiation using a glucose-lysine model system. The main findings are, hereby presented.
Medium-Chain Triacylglycerols (MCTs), and Long-Chain Triacylglycerols (LCTs) were a kind gift from Nisshin OilliO Group Ltd., Kanagawa, Japan. Tryptophan, glucose, N’, N’-dimethylformamide and lysine were purchased from Nakarai Tesque, Inc. Kyoto, Japan. Fatty acids (C8:0, C10:0, C18:2, C20:4ï¼›more than 95% purity) were purchased from Wako Pure Chemical Ind. Ltd., Japan. Ferric pyrophosphate (soluble) was purchased from Yoneyama Chemical Ind. Co., Ltd., Osaka, Japan. Nitro Blue Tetrazolium (NBT) and Diformazan were purchased from Tokyo Chemical Ind. Co., Ltd., Tokyo, Japan. Superoxide dismutase (SOD; from bovine erythrocytes), hydrogen peroxide (H2O2), and 2,5-dimethylfuran were purchased from Sigma-Aldrich Inc. USA. Other reagents were of special grade and were obtained from Nacalai Tesque, Inc. Kyoto, Japan.
Preparation of Glucose-lysine systems and light irradiation
Glucose-lysine systems were prepared as 20 mM glucose and lysine, using 50 ml of 0.25 M phosphate buffer (pH 7.4). MCTs, LCTs and various fatty acids were adjusted to 10 mM using methyl alcohol and from this, 10.0 ml was added to the 20 mM glucose-lysine, thereafter light irradiation of 10,000 lux was done for 60 min at 25°C.
Determination of advanced glycation endproducts (AGEs) in light irradiation-induced glucose-lysine systems
AGEs were extracted from glucose-lysine systems using 50 ml of 0.25 M phosphate buffer (pH 7.4). AGEs are brown pigments and the absorbance at 420 nm (brown color development) was done with the Spectrophotometer (SmartSpec PlusBio-Rad Laboratories, Inc. K.K. Tokyo Japan).
Measurement of reactive oxygen species at the time of light irradiation
Reactive oxygen species generated at the time of light irradiation were measured according to the following method. The generation of superoxide anion (02-) was determined by assessment of the reduction of NBT (Toyosaki et al., 1984). NBT (1.5 × 10-4 M, 0.5 ml) solution was added to 10.0 ml of products using glucose-lysine systems with MCTs, LCTs and various fatty acids. After the mixture was light irradiated at 10,000 lux for 60 min, the deposit of diformazan formed from the reduction of NBT by 02- was collected and dissolved in 4.0 ml N’, N’-dimethylformamide. Absorbance at 560 nm was done with the Spectrophotometer (U-3900, Hitachi High-Technologies GLOBAL, Co., Japan) and was measured to estimate the amount of 02- produced in the test sample, from a calibration curve prepared with the use of standard diformazan solution. Generation of hydrogen peroxide (H2O2) was measured by the method of Lijie and Yinsheng (2007).
Statistical analysis
All data were expressed as the mean ± standard deviation. Statistical analysis was performed using the unpaired student’s t-test (KaleidaGraph, Ver. 4.0; Synergy software, PA, USA). Differences in mean values among groups were assessed using the Tukey-Kramer multiple comparison test (Instat Ver. 3.0; GraphPad software, Inc., CA, USA). The level of significance was set at p<0.05 for all statistical tests.
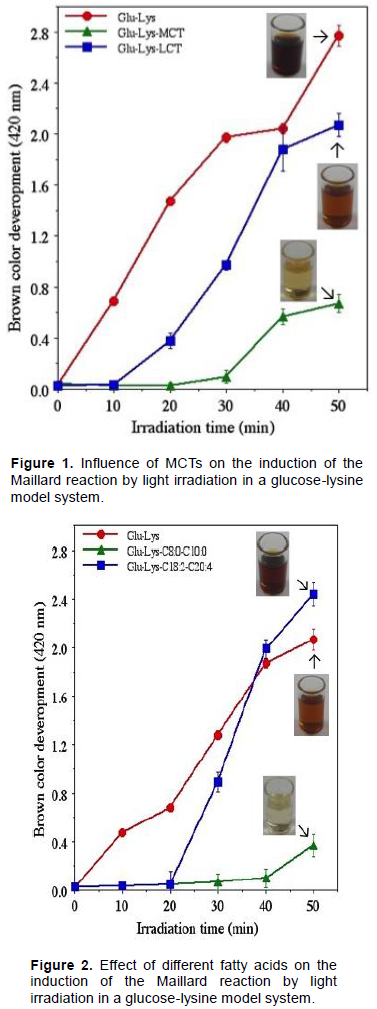
Influence of MCTs on the induction of Maillard reaction by light irradiation in a glucose-lysine model system
To determine whether MCTs have an inhibitory effect on the light irradiation-induced Maillard reaction in a glucose-lysine model system, the impact of the presence of MCTs to that of LCTs was compared (Figure 1). The Maillard reaction occurred under all light irradiation conditions, however, it was inhibited in the presence of both MCTs and LCTs relative to the control with no fatty acids present. Further, the Maillard reaction was inhibited to a significantly greater degree in the presence of MCTs than LCTs. This study, thus, demonstrated that the glucose-lysine Maillard reaction is induced by light irradiation and yet inhibited by the presence of MCTs. Taken together with previous study results (Toyosaki, 2018) the inhibitory effect of MCTs on light irradiation induced glucose-lysine Maillard reaction is, hereby, proposed.
Effect of different fatty acids on the induction of the Maillard reaction by light irradiation
MCTs were found to have a greater inhibitory effect on the Maillard reaction than LCTs. Given the likelihood that these differences in inhibitory effect are related to differences in fatty acid composition, an investigation was conducted using both saturated and unsaturated fatty acids of varying chain lengths. In this experimental system, saturated fatty acids with 8 and 10 carbons (C8:0 and C10:0, respectively) which constitute the MCTs were used. For comparison, unsaturated fatty acids with 18 and 20 carbons (C18:2 and C20:4, respectively), which constitute the LCTs were used. The results of these comparisons are shown in Figure 2.
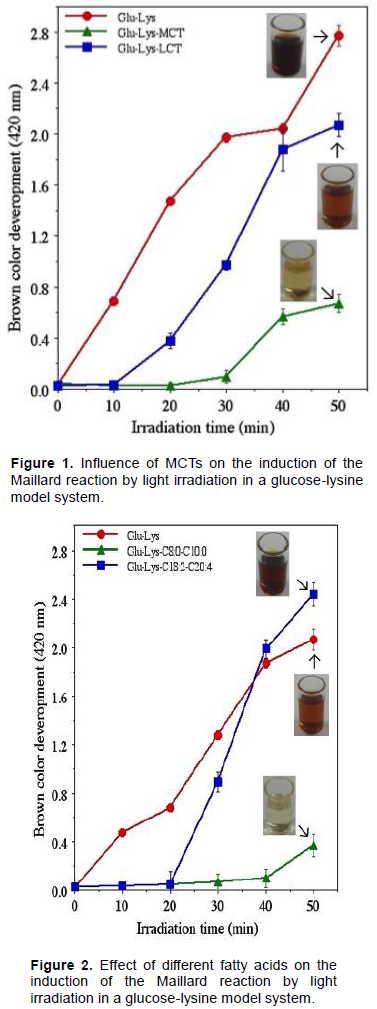
In the absence of fatty acids, the Maillard reaction was immediately induced with the start of light irradiation. In the presence of LCTs, the Maillard reaction was induced after 20 min of light irradiation. In the presence of MCTs, the Maillard reaction was negligibly induced after 40 min of light irradiation and only slightly induced after about 50 min of light irradiation. These results indicate that the medium-chain fatty acids that comprise MCTs inhibit the induction of the Maillard reaction by light irradiation.
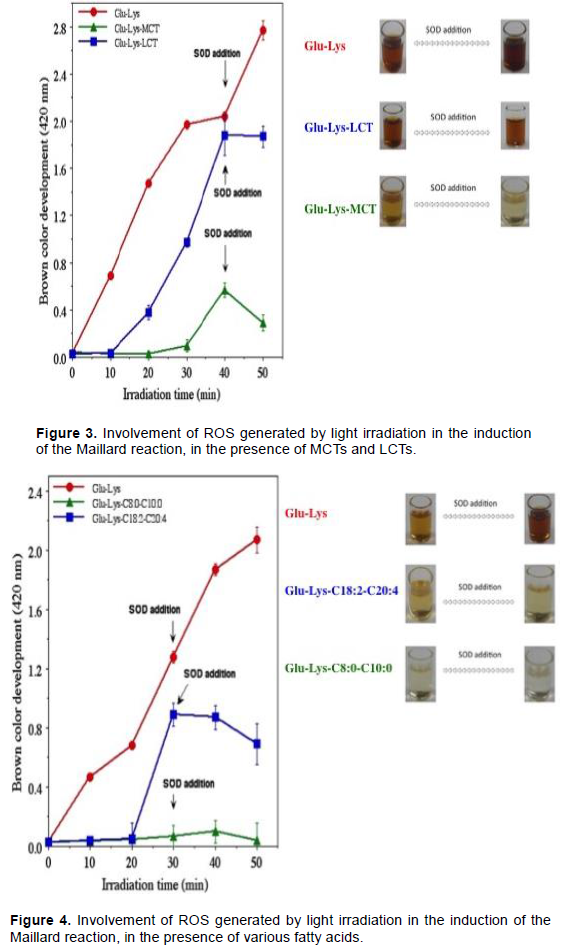
Involvement of ROS in the induction of the Maillard reaction by light irradiation in the presence of MCTs and LCTs
Based on the finding that Maillard reaction is induced by light irradiation in the glucose-lysine system and the strong likelihood that ROS are generated by light irradiation, the addition of SOD, which eliminates superoxide anions (O2-) was tested, in order to investigate the involvement of ROS in the induction of the Maillard reaction (Figure 3). The addition of SOD at 40 min after the start of light irradiation had an inhibitory effect on the Maillard reaction in the presence of both MCTs and LCTs. The inhibitory effect on the Maillard reaction was especially striking in the presence of MCTs. In contrast, in the absence of fatty acids, induction of the Maillard reaction was further promoted by the addition of SOD. These results strongly suggest that the induction of the Maillard reaction is accelerated with the generation of ROS by light irradiation. Although ROS are generated in the presence of both MCTs and LCTs, it is likely that differences in the fatty acid composition of MCTs and LCTs substantially influence the generation of ROS. Given the possibility that this influence is due to differences in chain length and degree of saturation, the use of fatty acids of varying compositions was investigated.
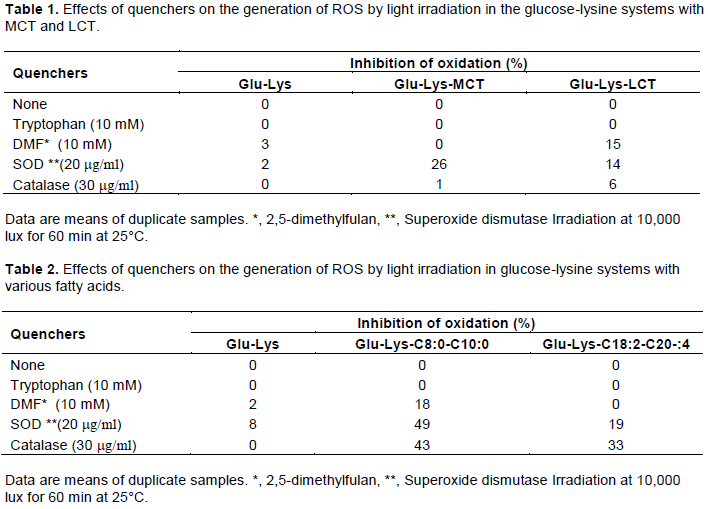
Involvement of ROS generated by light irradiation in the induction of the Maillard reaction, in the presence of various fatty acids
The addition of SOD was done to investigate the generation of ROS by light irradiation, in the presence of various fatty acids with different molecular structures (Figure 4). In the absence of fatty acids, addition of SOD at 30 min after the start of light irradiation accelerated the induction of Maillard reaction, however, in the presence of both saturated and unsaturated fatty acids, the addition of SOD inhibited the induction of Maillard reaction. These results reveal that, in the presence of fatty acids, ROS are generated by light irradiation and also that the generated oxygen species play a role in the induction of the Maillard reaction.
The inhibitory effect on the Maillard reaction was greater in the presence of MCTs than LCTs. This difference is likely largely attributable to differences in the amount of reactive oxygen generated and differences in the reactivity of the species generated. Consequently, the relationship between the ROS generated and the inhibitory effect on the Maillard reaction was investigated.
Involvement of ROS generated by light irradiation in the presence of MCTs and LCTs
Using quenching agents for various ROS, the role played by different ROS in the induction of Maillard reaction in the presence of MCTs or LCTs was investigated (Table 1). The inhibition of the induction of Maillard reaction in the presence of both MCTs and LCTs by SOD suggests that O2- is involved in the Maillard reaction. The lack of inhibitory effect by the addition of tryptophan and 2,5-dimethylfuran (DMF) on the Maillard reaction in the presence of MCTs indicates that singlet molecular oxygen (1O2) does not play a role in the induction of the Maillard reaction. In contrast, quenching of 1O2 by DMF inhibited induction of the Maillard reaction in the presence of LCTs, suggesting that 1O2 plays a role in the Maillard reaction. These results suggest that the chemical properties of MCTs and LCTs differ, though slightly, in terms of quenching ROS. Catalase did not accelerate the Maillard reaction in the presence of MCTs, indicating that hydrogen peroxide (H2O2) does not play a role in the Maillard reaction. On the other hand, catalase had an inhibitory effect, though small, on the Maillard reaction in the presence of LCTs, suggesting that H2O2 is at least involved in the induction of the Maillard reaction.
Given that the main mechanisms for generation of ROS are the Fenton reaction and the Haber-Weiss reaction, this study demonstrated that hydroxy radicals (.OH), a ROS, play a role in the induction of the Maillard reaction. Based on the fact that the addition of ferric iron (Fe3+) inhibited induction of the Maillard reaction in the presence of both MCTs and LCTs, it is proposed that .OH generated by reactions 1 and 2 is the main ROS involved in the Maillard reaction in the model system used in this study.
Involvement of ROS generated by light irradiation in the presence of various fatty acids
This study demonstrated that ROS are generated by light irradiation and play a role in induction of the Maillard reaction. The observed differences between LCTs and MCTs are likely as a result of differences in the mechanism for generating ROS. It was further observed that these differences are largely attributable to differences in the fatty acid composition of LCTs and MCTs (differences in degree of unsaturation) and that the chemical reactions with respect to the Maillard reaction also differ. Accordingly, investigations were done to ascertain the mechanisms for generation of ROS based on fatty acid composition and the results are shown in Table 2. The fact that DMF, SOD, and catalase accelerated the Maillard reaction in the presence of saturated fatty acids suggests that, although 1O2, O2-, and H2O2 are generated in the presence of saturated fatty acids, these are not directly involved in the Maillard reaction. Meanwhile, the addition of SOD and catalase demonstrated that O2- and H2O2 play a role in the induction of the Maillard reaction in the presence of unsaturated fatty acids. The accelerated induction of the Maillard reaction with the addition of Fe3+ in the presence of highly unsaturated fatty acids suggests that .OH may be the main ROS involved. Results of the investigation into mechanisms for generation of ROS indicate that different ROS play different roles in the glucose-lysine Maillard reaction induced by light irradiation. Furthermore, the mechanisms for generation of ROS were found to differ slightly due to differences in the chemical properties of constituent fatty acids and the extent of involvement in the Maillard reaction.
Relationship between mechanisms for generation of ROS by light irradiation and the Maillard reaction in a glucose-lysine model system
The main mechanisms for generation of ROS are the Fenton reaction 1 and the Haber-Weiss reaction 2 (Kellogg and Fridovich, 1975). It is proposed that the main ROS contributing to the Maillard reaction in the study experimental system is .OH. Consequently, water-soluble heme iron (Biocon, Japan Ltd.) was used to determine the main ROS generated in the experimental system used. The finding that the addition of water-soluble heme iron accelerated induction of the Maillard reaction in the presence of both MCTs and LCTs suggests that .OH in the ROS is involved in the induction of Maillard reaction in the experimental system used. Furthermore, given that no water-soluble iron exist in the experimental system used, it is highly likely that the hydroxy radicals were generated by the Haber-Weiss reaction 2.
Fe3+ + O2- ® Fe2+ + O2
Fe2++ H2O2 ® Fe3++ ·OH + OH- (1)
O2-+ H2O2 ® OH- + ·OH + O2 (2)
The findings from this study suggest that ROS generated by light irradiation play a role in inducing glucose-lysine Maillard reactions in the presence MCTs or LCTs. Induction of the glucose-lysine Maillard reaction was inhibited in the presence of MCTs. .OH generated through the Haber-Weiss reaction is thought to be the main ROS involved in induction of the Maillard reaction. This study is, possibly, the first to experimentally demonstrate that ROS are involved in induction of the Maillard reaction by light irradiation and contributes substantially to knowledge in the area of food science.
The authors have not declared any conflict of interests.