ABSTRACT
Escherichia coli present the ampC naturally, and the observation of phenotypical resistance to cefoxitin is related to this gene deregulation. Mutations in the regulatory region in ampC cause exaggerated expression. The most frequent alterations in the E. coli AmpC promoter/attenuator leading to this overexpression is described at the positions: -88, -82, -42, -18, -1 and +58. Mastitis studies were carried in Rio de Janeiro and Mato Grosso, Brazil. Two cefoxitin and amoxicillin-clavulanic acid-resistant E. coli from farms animals were unusually detected once these characteristics are not observed together in this species. The objective of this work was to determine if these isolates had a chromosomal gene mutation, determining AmpC hyperproduction. After DNA sequencing, mutations were observed at -88, -82, -73, -18, -1 and +58 positions, confirming the initially suspected AmpC hyperexpression. In Brazil, this is the first work to report E. coli hyperproducing this enzyme.
Key words: Ampc attenuator, AmpC hyperproduction, ampC promoter, bovine feces, mastitic milk.
AmpC is a serine-β-lactamase that belongs to group 1 of Bush-Jacoby-Medeiros and class C of Ambler classifications. This enzyme is codified by the chromosomal gene, ampC, which is a natural gene in Escherichia coli species. The ampC is a non-inducible gene in this species because it has lost its regulator gene. So, the resistance to cephamycins is not phenotypically observed in this species (Ambler, 1980; Bush and Jacoby, 2010). The hyperproduction of AmpC in E. coli caused by spontaneous mutations that produce
deregulation of ampC has been reported and is responsible to resistance to first, second and third-generation cephalosporins and to Extended-Spectrum Beta-Lactamase (ESBL) inhibitors (Siu et al., 2003; Jorgensen et al., 2010; Kohlmann et al., 2018). This hyperproduction does not cause resistance to cefepime that is related to ESBL co-production or Extended-Spectrum AmpC (ESAC) production in the isolates. AmpC may occur in bacteria producing another β-lactamase as ESBL, and it decreases the therapeutic options in the treatment of bacterial infectious diseases (Kojima et al., 2005; Mammeri et al., 2006; Rodríguez-Martínez et al., 2012).
Two regions are associated with controlling the enzyme production. The first is the promoter region that contains two important boxes, -35 box and -10 box, located between -42 and -18 positions. Another critical region is the attenuator of the ampC that is located between +17 and +37 locations (Olsson et al., 1983; Caroff et al., 1999; Corvec et al., 2002; Siu et al., 2003; Jorgensen et al., 2010). The mutations in the ampC gene may occur alone or in combination, although a single mutation in a specific position is sufficient to cause high enzyme production (Olsson et al., 1983).
The most frequent insertions or deletions in hyperproducers ampC E. coli occurs in -88, -82, -42, -18, -1 and +58 positions, but mutations at -32, -11, +6, +24 and +31 (Olsson et al., 1983; Caroff et al., 1999; Caroff et al., 2000; Haenni et al., 2014). There are data in the literature about these alterations in human clinical strain, but in E. coli from animal samples, it is not frequently demonstrated. Considering E. coli isolated from animals, these mutations had been described in Spain, Denmark and Japan (Briñas et al., 2002; Olesen et al., 2004; Kojima et al., 2005; Hiroi et al., 2011). However, there are no data about AmpC-hyperproducing E. coli in Brazil. The aim of this work was to detect the mechanism responsible for AmpC phenotypic characteristics observed in two E. coli from feces and milk in dairy cows during an antimicrobial resistance study.
E. coli were isolated from milk and feces of cows on dairy farms in Rio de Janeiro (RJ) and Mato Grosso (MT), Brazil, within six years (2009-2015) (protocol no. CEUA-3664040915, Federal Rural University of Rio de Janeiro, Brazil). Routine biochemical tests identified 238 E. coli isolates, which was further confirmed by matrix-assisted laser desorption/ionization time-of-flight MS assay (Rodrigues et al., 2017; Santiago, 2017). Antimicrobial resistance of the E. coli isolates was obtained by the disk diffusion method, and two strains (G27 and S10) was suspected to AmpC hyperproduction. The cefoxitin-resistance was confirmed by MIC, according to CLSI (2017). To evaluate ampC promoter/attenuator was used the primers AB1 (5′-GATCGTTCTGCCGCTGTG-3′) and AmpC2 (5′-GGGCAGCAAATGTGGAGCAA-3′), yielding a 271-bp amplification product (Corvec et al., 2002). They were sequenced (ABI 3130xl, Applied Biosystems, São Paulo, Brazil) and analyzed by DNA Sequence Assembler version 4 (HeracleBioSoft, Arges, Romania) and Mega software version 7 (Caspermeyer, 2016). The sequences were deposited in a GenBank database (Genbank accession numbers: MK559376, MK559377 and MK559378). E. coli ATCC 25922 obtained from FIOCRUZ (Rio de Janeiro, Brazil) were used as a control for phenotypic and genotypic tests (CLSI, 2017).
The isolates were sequestered from Agar MacConkey and thereafter subjected to Gram test to confirm the morphological and tinctorial characteristics. The isolates were identified as E. coli by phenotypic laboratory tests in accordance with Koneman et al. (2010). All E. coli were confirmed by matrix-assisted laser desorption/ionization time-of-flight MS assay (Rodrigues et al., 2017).
After specie identification, these isolates were submitted for antibiotics tests for detection of resistance to β-lactams. So, two E. coli isolates, G27 and S10, presented resistance to cefoxitin, amoxicillin and amoxicillin-clavulanic acid and susceptibility to cefepime, suspected to AmpC hyperproduction. G27 and S10 presented MIC < 4 (CLSI 2017). The E. coli G27 was isolated from dairy cows' milk samples in suspected mastitis cases in Rio de Janeiro (2010), and the E. coli G27 was isolated from dairy cows' milk samples in suspected mastitis cases in Rio de Janeiro during evaluations in 2010, and the E. coli S10 was isolated in cow feces from Mato Grosso, in 2014.
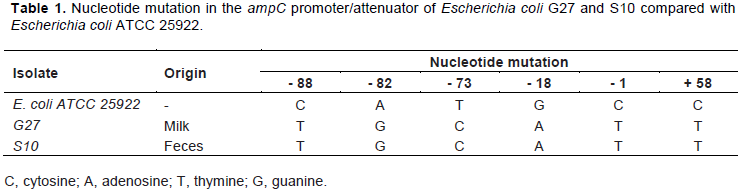
These isolates were submitted to PCR and the primers used include the -35 box, the -10 box, and the attenuator segment. The sequencing of the regulatory region of G27 and S10 were analyzed using DNA Sequence Assembler version 4 (HeracleBioSoft, Arges, Romania) and Mega software version 7. Some alterations were observed in important positions. G27 and S10 ampC regulatory region revealed the mutations previously described in the literature responsible for causing the AmpC hyperproduction. Both isolates presented the most common substitutions for -88, -82, -18, -1 and +58 positions, although they have also shown a replacement at -73 position (Table 1).
Many authors described alterations in important regions in E. coli ampC regulator from human and animals samples. Naturally, E. coli produces AmpC enzyme in a low quantity because it is responsible for wall maintenance as a biological function (Johnson et al., 2013; Santiago et al., 2016).
The ampC promoter studies demonstrated -1 and +58 mutations are associated with increased strength of promoter taking higher gene transcription in E. coli (Olsson et al., 1983; Caroff et al., 1999; Corvec et al., 2002; Jorgensen et al., 2010; Haenni et al., 2014). Siu et al. (2003) and Yu et al. (2009) found replaced C (cytosine) by T (timine) at -58 position as in this study.
In other studies involving E. coli from animal was detected this species expressing resistance to cefoxitin with mutations at -88, -82 -42, -32, -18, -1, +37 +58 and +70 positions in promoter region of ampC gene (Briñas et al., 2002; Olesen et al., 2004; Kojima et al., 2005; Hiroi et al., 2011; Haenni et al., 2014). The mutation in -73 position was only observed in human isolates studied by Yu et al. (2009). They described one E. coli containing mutation at -73 position and it was associated with other mutations, among them at 80, -28, -1, +58 and +82 position.
The AmpC-hyperproducing E. coli has not been reported in dairy cattle. However, many positions of mutation observed in E. coli beef cattle, broiler, and meat were described in human samples (Briñas et al., 2002; Hiroi et al., 2011). These changes demonstrate that there is a relationship between the transmission of these bacteria in the food chain and dissemination through the environment.
Another important fact is that some mutations were observed in human isolates before been reported in animal samples. That way, we believe the dissemination of these bacteria occurred before 1999, but only years later; the studies involving animals were published. This may have happened due to the new paradigm implemented by the One Health concept in 2007.
Interestingly, the occurrence of AmpC-hyperproducing E. coli was low during the period evaluated. den Drijver et al. (2018) studied the prevalence of AmpC-producing E. coli from a Dutch teaching hospital and affirmed these characteristics had been declined. In Brazil, AmpC-hyperproducing E. coli had not been reported until now, so it is challenging to state about the epidemiology of these isolates.
These mutations demonstrate that AmpC enzyme has been hyperproduced by these isolates. This study indicates that AmpC-hyperproducing E. coli also exist in Brazil, specifically in dairy herds. This is the first Brazilian report to consider hyperproduction of AmpC enzyme in E. coli isolated from dairy cows. Future studies can be conducted with the aim of identifying other animals and animal products containing E. coli with these characteristics.
The authors have not declared any conflict of interests.
This work was financially supported by Foundation for Research Support of the State of Rio de Janeiro (FAPERJ, Brazil) and Ph.D. Sandwich Scholarship (Coordination of Improvement of Higher Level Personnel; CAPES) PDSE 99999.008079/2014-05 (Brazil).
REFERENCES
|
Ambler RP (1980). The structure of β-lactamases. Philosophical Transactions of the Royal Society B: Biological 289(1036):321-331.
Crossref
|
|
|
|
Briñas L, Zarazaga M, Sáenz Y, Ruíz-Larrea F, Torres C (2002). B-lactamases in Ampicillin-Resistant Escherichia coli Isolates from Foods, Humans and Healthy Animals. Antimicrobial Agents and Chemotherapy 46(10):3156-3163.
Crossref
|
|
|
|
|
Bush K, Jacoby GA (2010). Updated functional classification of β-lactamases. Antimicrobial Agents and Chemotherapy 54(3):969-976.
Crossref
|
|
|
|
|
Caroff N, Espaze E, Bérard I, Richet H, Reynaud A (1999). Mutations in the ampC promoter of Escherichia coli isolates resistant of oxyimino cephalosporins without extended-spectrum beta-lactamase production. FEMS Microbiology Letters 173(2):459-465.
Crossref
|
|
|
|
|
Caroff N, Espaze E, Gautreau D, Richet H, Reynaud A (2000). Analysis of the effects of -42 and -32 ampC promoter mutations in clinical isolates of Escherichia coli hyperproducing AmpC. Journal of Antimicrobial Chemotherapy 45:783-788.
Crossref
|
|
|
|
|
Caspermeyer J (2016). Mega Evolutionary Software re-engineered to Handle Today's Big Data Demands. Molecular Biology and Evolution 33(7):1887.
Crossref
|
|
|
|
|
CLSI (2017). Performance Standards for Antimicrobial Susceptibility Testing. 27th ed. CLSI supplement M100. Wayne, PA: Clinical and Laboratory Standard Institute: 2017.
|
|
|
|
|
Corvec S, Caroff N, Espaze E, Marraillac J, Reynaud A (2002). -11 Mutation in the ampC Promoter Increasing Resistance to β-Lactams in a Clinical Escherichia coli Strain. Antimicrobial Agents and Chemotherapy 46(10):3265-3267.
Crossref
|
|
|
|
|
den Drijver E, Verweij JJ, Verhulst C, Oome S, Soer J, Willemsen I, Schrauwen EJA, Kluytmans-van den Bergh MFQ, Kluytmans JAJW (2018). The decline in AmpC β-lactamase-producing Escherichia coli in a Dutch teaching hospital (2013-2016). PLoS One 13(10):e0204864.
Crossref
|
|
|
|
|
Hiroi M, Harada T, Kawamori F, Takahashi N, Kanda T, Sugiyama K, Masuda T, Yoshikawa Y, Ohashi N (2011). A survey of b-lactamase-producing Escherichia coli in farm animals and raw retail meat in Shizuoka Prefecture, Japan. Japanese Journal of Infectious Diseases 64:153-155.
|
|
|
|
|
Haenni M, Châtre P, Madec J (2014). Emergence of Escherichia coli producing extended-spectrum AmpC β-lactamases (ESAC) in animals. Front Microbiology 5:53.
Crossref
|
|
|
|
|
Johnson JW, Fisher JF, Mobashery S (2013). Bacterial cell-wall recycling. Annals of New York Academy of Science 1277(1):54-75.
Crossref
|
|
|
|
|
Jorgensen RL, Nielsen JB, Friis-Moller A, Fjeldsoe-Nielsen H, Schonning K (2010). Prevalence and molecular characterization of clinical isolates of Escherichia coli expressing an AmpC phenotype. Antimicrobial Chemotherapy 65:460-464.
Crossref
|
|
|
|
|
Kohlmann R, Bähr T, Gatermann SG (2018). Species-specific mutation rates for ampC derepression in Enterobacterales with chromosomally encoded inducible AmpC β-lactamase. Journal of Antimicrobial Chemotherapy 73(6):1530-1536.
Crossref
|
|
|
|
|
Kojima A, Yoshikazu I, Kanako I, Esaki H, Asai T, Oda C, Tamura Y, Takahashi T, Yamaguchi K (2005). Extended-Spectrum--Lactamase-Producing Escherichia coli Strains Isolated from Farm Animals from 1999 to 2002: Report from the Japanese Veterinary Antimicrobial Resistance Monitoring Program. Antimicrobial Agents and Chemotherapy 49(8):3533-3537.
Crossref
|
|
|
|
|
Koneman EW, Allen SD, Janda WM, Schreckenbergere PC, Winn WC (2010). Microbiologic Diagnostic. MEDS, Rio de Janeiro, RJ.
|
|
|
|
|
Mammeri H, Poirel L, Fortineau N, Nordmann P (2006). Naturally Occurring Extended-Spectrum Cephalosporinases in Escherichia coli. Antimicrobial Agents and Chemotherapy 50(7):2573-2576.
Crossref
|
|
|
|
|
Olesen I, Hasman H, Aareestrup FM (2004). Prevalence of b-lactamases among Ampicillin-Resistant Escherichia coli and Salmonella isolated from food animals in Denmark. Microbial Drug Resistance 10(4):334-340.
Crossref
|
|
|
|
|
Olsson O, Bergstrom S, Lindberg FP, Normark S (1983). AmpC β-lactamase hyperproduction in Escherichia coli: Natural ampicillin resistance generated by horizontal chromosomal DNA transfer from Shigella. Proceedings of the National Academy of Science USA 80:7556-7560.
Crossref
|
|
|
|
|
Rodrigues NMB, Bronzato GF, Santiago GS, Botelho LAB, Moreira BM, Coelho IS, Souza MMS, Coelho SMO (2017). The matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF MS) identification versus biochemical tests: A study with enterobacteria from a dairy cattle environment. Brazilian Journal of Microbiology 48:132-138.
Crossref
|
|
|
|
|
Rodríguez-Martínez JM, Fernández-Echauri P, Fernández-Cuenca F, Alba PD, Briales, A, Pascual A (2012). Genetic characterization of an extended-spectrum AmpC cephalosporinase with hydrolyzing activity against fourth-generation cephalosporins in a clinical isolate of Enterobacter aero genes selected in vivo. Journal of Antimicrobial Chemotherapy 67(1):64-68.
Crossref
|
|
|
|
|
Santiago GS, Motta CC, Bronzato GF, Gonçalves D, Souza MMS, Coelho IS, Ferreira HN, Coelho SMO (2016). Revisão: Produção de β-lactamases do Tipo AmpC em Enterobacteriaceae. Revista Brasileira Medicina Veterinária 38(Supl.3):17-30.
|
|
|
|
|
Santiago GS (2017). Compreensão de mecanismos fenotípicos e genotípicos relacionados à produção de β-lactamases do tipo AmpC em Enterobacteriaceae. MS Thesis. Federal Rural University of Rio de Janeiro, RJ, Brazil. 2017.
|
|
|
|
|
Siu LK, Lu PL, Chen JY, Lin FM, Chang SC (2003). High-Level Expression of AmpC b-Lactamase Due to Insertion of Nucleotides between _10 and _35 Promoter Sequences in Escherichia coli Clinical Isolates: Cases Not Responsive to Extended-Spectrum-Cephalosporin Treatment. Antimicrobial Agents and Chemotherapy 47(7):2138-2144.
Crossref
|
|
|
|
|
Yu W, Bing L, Zhenhua L (2009). ampC promoter and attenuator mutations affect function of three Escherichia coli strains. Current Microbiology 59(3):244-247.
Crossref
|
|