Full Length Research Paper

ABSTRACT
INTRODUCTION
MATERIALS AND METHODS
RESULTS
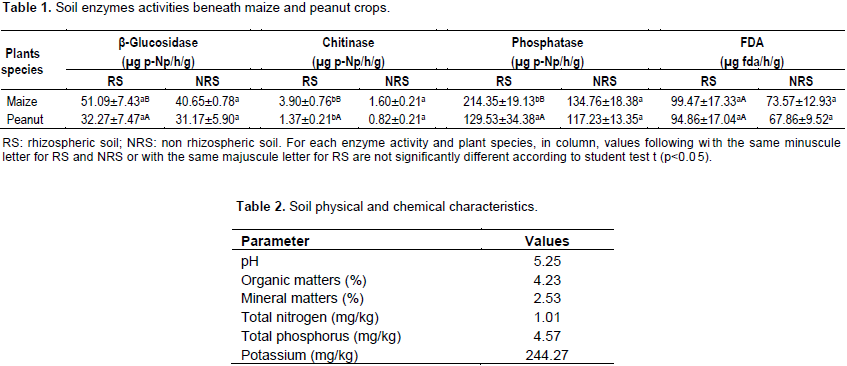



DISCUSSION
CONCLUSION
CONFLICT OF INTERESTS
ACKNOWLEDGEMENTS
REFERENCES
|
Acosta-Martínez V, Cruz L, Sotomayor-Ramírez D, Pérez-Alegría L (2007). Enzyme activities as affected by soil properties and land use in a tropical watershed. Appl. Soil Ecol. 35:35-45. |
|
|
Adam G, Duncan H (2001). Development of a sensitive and rapid method for the measurement of total microbial activity using fluorescein diacetate (FDA) in a range of soils. Soil Biol. Biochem. 33:943-951. |
|
|
Amato M, Ladd JN (1988). An assay for microbial biomass based on ninhydrin-reactive nitrogen in extracts of fumigated soils. Soil Biol. Biochem. 20: 107-l14. |
|
|
Asibuo JY, Akromah R, Adu-Dapaah HK, Safo-Kantanka O (2008). Evaluation of nutritional quality of groundnut (Arachis hypogaea L.) from Ghana. Afr. J. Food Agric. Nutr. Dev. 8(2):133-150. |
|
|
Azcon R, Rubio R, Barea JM (1991). Selective interactions between different species of mycorrhizal fungi and Rhizobium meliloti strains, and their effects on growth, N2 fixation (N15) in Medicago sativa at four salinity levels. New Phytol. 117:399-404. |
|
|
Bâ AM, Dalpé Y, Guissou T (1996). Les Glomales d'Acacia holosericea et d'Acacia mangium. Bois et Forêt des Tropiques. 250:5-18. |
|
|
Bago B, Pfeffer PE, Shachar-Hill Y (2000). Carbon metabolism and transport in arbuscular mycorrhizas. Plant Physiol. 124(3):949-958. |
|
|
Barea JM (2015). Future challenges and perspectives for applying microbial biotechnology in sustainable agriculture based on a better understanding of plant-microbiome interactions. J. Soil Sci. Plant Nutr. 15(2):261-282. |
|
|
Barea JM, Toro M, Orozco MO, Campos E, Azcón R (2002). The application of isotopic (32P and 15N) dilution techniques to evaluate the interactive effect of phosphate-solubilizing rhizobacteria, mycorrhizal fungi and Rhizobia to improve the agronomic efficiency of rock phosphate for legume crops. Nutr. Cycl. Agroecosyst. (in press). |
|
|
Baxter JW, Dighton J (2001). Ectomycorhizal diversity alters growth and nutrient acquisition of grey birch (betulapopulifolia) seedlings in host-symbiont culture conditions. New Phytol. 152:139-149. |
|
|
Berg B, McClaugherty C (2014). Plant litter: decomposition, humus formation, carbon sequestration, 3rd ed. Springer, Verlag, Berlin, Heidelberg, Germany. |
|
|
Böhme L, Böhme F (2006). Soil microbiological and biochemical properties affected by plant growth and different long-term fertilisation. Europ. J. Soil Biol. 42:1-12. |
|
|
Bremner JM (1960). Determination of nitrogen in soil by the Kjeldahl method. J. Agr. Sci. 55:11-33. |
|
|
Brockwell J (1980). Experiment with crop and pasture legumes: principles and practice. In: Bergersen FJ (Ed), Methods for evaluating BNF. Wiley. New York, 417-488. |
|
|
Cheng W, Parton WJ, Gonzalez-Meler MA, Phillips R, Asao S, McNickle GG, Brzostek E, Jastrow JD (2014). Synthesis and modeling perspectives of rhizosphere priming. New Phytol. 201:3-44. |
|
|
Chi XQ, Zhang JJ, Zhao S, Zhou NY (2013). Bioaugmentation with a consortium of bacterial nitrophenol-degraders for remediation of soil contaminated with three nitrophenol isomers. Environ. Pollut. 172:33-41. |
|
|
Chen L, Hu X, Yang W, Xu Z, Zhang D, Gao S (2015). The effects of arbuscular mycorrhizal fungi on sex-specific responses to Pb pollution in Populus cathayana. Ecotox. Environ. Safe. 113:460-468. |
|
|
Dalpé Y (1997). "Vesicular-arbuscular mycorrhiza". In soil sampling and methods of analysis, 3rd edn, Carter MR (ed). Can. Soc. Soil Science. pp. 287-301. |
|
|
Diatta ILD, Kane A, Agbangba CE, Sagna M, Diouf D, Bertossi FA, Duval Y, Borgel A, Sane D (2014). Inoculation with arbuscular mycorrhizal fungi improves seedlings growth of two sahelian date palm cultivars (Phoenix dactylifera L., cv. Nakhlahamra and cv. Tijib) under salinity stresses. Adv. biosci. biotechnol. 5:64-72. |
|
|
Dobermann A, Cassman KG (2004). "Environmental dimensions of fertilizer nitrogen: what can be done to increase nitrogen use efficiency and ensure global food security?" in Agriculture and the nitrogen cycle: assessing the impacts of fertilizer use on food production and the environment. Mosier AR et al. ed. Washington DC: Island Press. pp. 261-278. |
|
|
Doley K, Jite PK (2012). Response of groundnut ('JL-24') cultivar to mycorrhiza inoculation and phosphorous application. Nat. Sci. Biol. 4(3):118-125. |
|
|
Duineveld BM, Kowalchuk GA, Keijzer A, van Elsas JD, van Veen JA (2001). Analysis of bacterial communities in the rhizosphere of Chrysanthemum via denaturing gradient gel electrophoresis of PCR-amplified 16S rRNA as well as DNA fragments coding for 16S rRNA. Appl. Environ. Microbiol. 67:172-178. |
|
|
Farrar JL (1995). Trees in Canada. Canadian forest service publications. Natural resources Canada, Canadian forest service, Headquarters, Ottawa, Co published by Fitzhenry and Whiteside Limited, Markham, Ontario. 502p. |
|
|
Gerdermann JW, Nicolson TH (1963). Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Trans. Br. mycol. Soc. 46:235-244. |
|
|
Gill TS, Singh RS (2002). Effect of Glomus fasciculatum and Rhizobium inoculation on VA mycorrhizal colonization and plant growth of chickpea. J. Mycol. Plant Pathol. 32: 162-167. |
|
|
Gottardi W, Nagl M (1998). Which conditions promote a remnant (persistent) bactericidal activity of chlorine covers? Zentralbl. Hyg. Umweltmed. 201:325-335. |
|
|
Graham PH (2008). Ecology of the root-nodule bacteria of legumes. Dilworth MJ et al. (eds.), Nitrogen-fixing leguminous symbioses. pp. 23-58. |
|
|
Guissou T, Bâ AM, Guinko S, Duponnois R, Plenchette C (1998). Influence des phosphates naturels et des mycorhizes à vésicules et à arbuscules sur la croissance et la nutrition minérale de Zizyphus mauritiana Lam. dans un sol à pH alcalin. InAnnales des sciences forestières. EDP Sciences. 55(8):925-931. |
|
|
Hamdan N, Kavazanjian EJ (2016). Enzyme-induced carbonate mineral precipitation for fugitive dust control. Géotechnique. 66:546-555. |
|
|
Hayano K (1973). A method for determination of β-glucosidase activity in soil. J. Soil Sci. Plant Nutr. 19: 103-108. |
|
|
Hirsch PR, Miller AJ, Dennis PG (2013). Do root exudates exert more influence on rhizosphere bacterial community structure than other rhizodeposits? Mol. Microb. Ecol. Rhizosphere. 1:229-242. |
|
|
Hite DRC, Auh C, Scandalios JG (1999). Catalase activity and hydrogen peroxide levels are inversely correlated in maize scutella during seed germination. Redox Rep. 4:29-34. |
|
|
Hungria M, Vargas MAT (2000) Environmental factors affecting N2 fixation in grain legumes in the tropics, with emphasis on Brazil. Field Crops Res. 65:151-164. |
|
|
Kellogg EA (2001). Evolutionary history of the grasses. Plant Physiol. 12:1198-1205. |
|
|
Kidd DR, Ryan MH, Haling RE, Lambers H, Sandral GA, Yang Z, Culvenor RA, Cawthray GR, Stefanski A, Simpson RJ (2016). Rhizosphere carboxylates and morphological root traits in pasture legumes and grasses. Plant Soil. 402:77-89. |
|
|
Kouakou Y, Kone B, Bonfoh B, Kientga S, N'Go Y, Savane I, Cisse G (2010). L'étalement urbain au péril des activités agro-pastorales à Abidjan, revue électronique en sciences de l'environnement, volume 10, numéro 2. |
|
|
Kruger M, Kruger C, Walker C, Stockinger H and Schuûler A (2012). Phylogenetic reference data for systematics and phylotaxonomy of arbuscular mycorrhizal fungi from phylum to species level. New Phytol. 193:970-984. |
|
|
Landon JR (1991). Booker Tropical Soil Manual. A handbook for soil survey and agricultural land evaluation in the tropics and subtropics. Longman, Booker Take limed, Oxon, UK. 474p. |
|
|
Lange M, Habekost M, Eisenhauer N, Roscher C, Bessler H, Engels C, Oelmann Y, Scheu S, Wilcke W, Schulze ED, Gleixner G (2014). Biotic and abiotic properties mediating plant diversity effects on soil microbial communities in an experimental grassland. Plos One 9(5):e96182. |
|
|
Lau JA, Bowling EJ, Gentry LE, Glasser PA, Monarch EA, Olsen WM et al. (2012). Direct and interactive effects of light and nutrients on the legume-rhizobia mutualism. Acta Oecol. 39:80-86. |
|
|
Leye EHM, Ndiaye M, Diouf M, Diop T, (2015). Etude comparative de l'effet de souches de champignons mycorhiziens arbusculaires sur la croissance et la nutrition minérale du sésame cultivé au sénégal. Afr. Crop Sci. J. 23(3):211-219. |
|
|
Li X, Chen Z (2004). Soil microbial biomass C and N along a climatic transect in the Mongolian steppe. Biol. Fertil. Soils. 39:344-351. |
|
|
Malherbe S, Marais D (2015). Nematode community profiling as a soil biology monitoring tool in support of sustainable tomato production: A case study from South Africa. Appl. Soil Ecol. 93:19-27. |
|
|
Martínez-García LB (2010). Micorrizas arbusculares en ecosistemas semiáridos. Respuesta a factor esdeestrés ambiental. Thesis Doctorales, Almería: Universidad de Almería. |
|
|
Martínez-García LB, Pugnaire FI (2009). Interacciones entre las comunidades de hongos formadores de micorrizas arbusculares y de plantas. Algunos ejemplos en los ecosistemas semiáridos. Ecosistemas. 18:44-54. |
|
|
Maseko S, Dakora F (2013). Rhizosphere acid and alkaline phosphatase activity as a marker of p nutrition in nodulated cyclopia and aspalathus species in the cape fynbos of South Africa. S. Afr. J. Bot. 89:289-295. |
|
|
Matus F, Rumpel C, Neculman R, Panichini M, Mora ML (2014). Soil carbon storage and stabilisation in andic soils: a review. Catena. 120:102-110. |
|
|
Merino C, Godoy R, Matus F (2016). Soil enzymes and biological activity at different levels of organic matter stability. J. Soil Sci. Plant Nutr. 16:14-30. |
|
|
Moir JL, Moot DJ (2010). Soil pH, exchangeable aluminium and lucerne yield responses to lime in a South Island high country soil. Proc. N. Z. Grassl. Assoc. 72:19-196. |
|
|
Morte A, Honrubia M (2002). Growth response of Phoenix canariensis to inoculation with arbuscular mycorrhizal fungi. Palms. 46:76-80. |
|
|
Mustafa AA, Othman R, Abidin MAZ, Ganesan V (2010). Growth response of Sweet Corn (Zea mays) to Glomus mossae inoculation over different ages. Asian J. Plant Sci. 9(6):337-343 |
|
|
Natywa M, Selwet M (2011). Respiratory and dehydrogenase activities in the soils under maize growth in the conditions of irrigated and non irrigated fields. Acta Sci. Pol. Agric. 10 (3):93-100 |
|
|
Ndoye F, Kane A, Bakhoum N, Sanon A, Fall D, Diouf D, Sylla SN, Bâ AM, Sy MO, Noba K (2013). Response of Acacia senegal (L.) Wild. to inoculation with arbuscular mycorrhizal fungi isolates in sterilized and unsterilized soils in Senegal. Agroforest. Syst. J. 87:941-952. |
|
|
Panettieri M, Knicker H, Murillo JM, Madejón E, Hatcher PG (2014). Soil organic matter degradation in an agricultural chronosequence under different tillage regimes evaluated by organic matter pools enzymatic activities and CPMAS 13C NMR. Soil Biol. Biochem. 78:170-181. |
|
|
Parham JA, Deng SP (2002). Detection, quantification and characterization of β-glucasaminidase activity in soil. Soil Biol. Biochem. 32:1183-1190. |
|
|
Parniske M (2008). Arbuscular mycorrhiza: the mother of plant root endosymbioses. Nat Rev Micro. 6: 763-775. |
|
|
Phillips JM, Hayman DS (1970). Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 55:158-161. |
|
|
Plenchette C, Bois JF, Duponnois R, Cadet P (2000). La mycorhization (Glomus aggregatum) du mil (Pennisetum glaucum). Etudes et Gestion des Sols, 7, 4. Numéro spécial. pp. 379-384. |
|
|
Plenchette C, Perrin R, Duvert P (1989). The concept of soil infectivity and method for its determination as applied to endomycorrhizas. Can. J. Bot. 67: \12-115. |
|
|
Rao RN, Talluri MVNK (2007). An overview of recent applications of inductively coupled plasma-mass spectrometry (ICP-MS) in determination of inorganic impurities in drugs and pharmaceuticals. J. Pharm. Biomed. Anal. 43:1-13. |
|
|
Rodríguezâ€Echeverría S, Teixeira H, Correia M, Timóteo S, Heleno R, Öpik M, Moora M (2016). Arbuscular mycorrhizal fungi communities from tropical Africa reveal strong ecological structure. New Phytologist. 213(1):380-390. |
|
|
Sánchez-Roque Y, Pérez-Luna Y, Becerra-Lucio A, Alvarez-Gutiérrez P, Pérez-Luna E, González-Mendoza D, Canseco-Pérez M, Salda-a-Trinidad S, Berrones-Hernández R (2016). Effect of arbuscular mycorrhizal fungi in the development of cultivars of Chili. Int. J. Adv. Agric. Res. 4:10-15. |
|
|
Sgrott AN, Booz MR, Pescador R, Heck TC, Stümor SL (2012). Arbuscular mycorrhizal inoculation increases biomass of Euterpe edulis and Archontophoenix alexandrae after two years under field conditions. Rev. Bras. de Ciên. Solo (36):1103-1112. |
|
|
Sherrell CG, Saunders WMH (1966). An evaluation of methods for the determination of total phosphorus in natural in soils. N. Z. J. Agr. Res. 9: 972-979. |
|
|
Smith MR, Charvat I, Jacobson RL (1998). Arbuscular mycorrhizae promote establishment of prairie species in a tall grass prairie restoration. Can. J. Bot. 76:1947-1954. |
|
|
Tabatabai MA, Bremner JM 1969. Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biol. Biochem. 1:301-307. |
|
|
Tedersoo L, Bahram M, Põlme S, Kõljalg U, Yorou NS, Wijesundera R, Ruiz LV, Vasco-Palacios AM, Thu PQ, Suija A, Smith ME (2014). Global diversity and geography of soil fungi. Science 346(6213):1256688. |
|
|
Trouvelot A, Kough JL, Gianinazzi-Pearson V (1986). Mesure du taux de mycorhization V. A d'un système radiculaire. Recherche de méthodes d'estimation ayant une signification fonctionnelle. In: Gianinazi-Pearson V, Gianinazzi S. Les mycorhizes: Physiologie et Génétique, 1er Séminaire Européen sur les mycorhizes, Dijon, INRA, Paris, pp.217-221. |
|
|
Turner BL, Hopkins DW, Haygarth PM, Ostle N (2002). β-Glucosidase activity in pasture soils. Appl. Soil Ecol. 20:157-162. |
|
|
Waceke JW, Waudo SW, and Sikora R (2001). Suppression of Meloidogynehapla by arbuscular mycorrhiza fungi (AMF) on pyrethrum in Kenya. Int. J. Pest Manag. 47(2):135-140. |
|
|
Wardle DA, Lindahl BD (2014). Disentangling global soil fungal diversity. Science 346:1052-1053. |
|
|
Yang Z, Liu P, Li Y, Ma L, Alva A, Dou Z, Chen Q, Zhang F (2013). Phosphorus in China's intensive vegetable production systems: over fertilization, soil enrichment, and environmental implications. J. Environ. Qual. 42:982-989. |
|
|
Zhou L, Zhou X, He Y, Shao J, Hu Z, Liu R, Zhou H, Hosseini BS (2016). Grazing intensity significantly affects belowground carbon and nitrogen cycling in grassland ecosystems: A meta-analysis. Glob. Change Biol. 22:31-57. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


