ABSTRACT
The microbial community structure from dark earth in a native archaeological site of the Lower Amazon was analyzed by PCR-DGGE, using 16S rRNA gene for prokaryote population and 18S rDNA and ITS regions (using clamp GC) for the eukaryote population. The bands were excised from gel and re-amplified for sequencing. The diversity found according to the region of amplification showed same profiles for the two primers pairs. The bacteria genus were: Bacillus, Klebsiella, Pantoea, Enterobacter, Lactobacillus, Escherichia, Leuconostoc and actinobacterias as Streptomyces and Microbacterium. Among the fungal community was Zygosaccharomyces, Lachancea, Saccharomyces, Cladosporium, Candida, Penicillium and Uncultured ascomycota and zygomycete were found. Molecular approaches revealed microbial groups that have never been reported in Lower Amazon soil as the Leuconostoc mesenteroides, Lactobacillus casei and Lactobacillus paracasei bacteria’s and Lachancea meyersii yeast. The soil pH was ~6.5; the soil had high levels of minerals with exception of Na (not detected) and Al (~0.2 mg/dm3). The organic matter was 3.5 dag/kg. This study also shows that the Amazon soil is rich in minerals. This can be an important factor in the species richness in the Amazon region. The present data show that the Lower Amazon represents a vast resource for the biotechnology area.
Key words: PCR-DGGE, fungi and yeast, soil microdiversity, soil chemical.
The soils are biodiverse ecosystems. Microorganisms are a component of these ecosystems (Pereira et al., 2006). Microorganisms are involved in biodegradation, decomposition and mineralization, and inorganic nutrient cycling in soils.
The Amazon soil contains great unexplored ground. The Terra Preta are anthropogenic soil (Lehmann et al., 2003) which are fertilized through increasing the cation exchange capacity and the nutrient content (Lehmann et al., 2003; Kim et al., 2007).
The microbial diversity present in Amazon soils is little known. Therefore, the Brazilian government provides financial support for research. The application of molecular techniques in the microbial diversity and structure analysis of these communities has been used previously (Bossio et al., 2005; Pereira et al., 2006; Castro et al., 2008; Taketani et al., 2013; Brossi et al., 2014). The molecular approaches have been proven to be powerful tools in providing an inventory of the microbial diversity in environmental samples (Ascher et al., 2010; Silva et al., 2012).
The assessment of diversity in Amazon soil is an important aspect in the quest for maintenance of soil biodiversity (Brossi et al., 2014). A characterization of the bacterial and fungal microbial community associated with Amazon soil from dark earth in a native archaeological site of the Lower Amazon is lacking. Furthermore, it is likely that richness of these bacterial and fungal communities is affected by anthropogenic modification of environment. The objectives of this study were: Firstly, molecular survey of the bacterial and fungal communities associated with Amazon soil, based on the sequencing of different rDNA regions, and secondly to analyze the physicochemical characteristics of this soil.
Soil sampling
Anthropogenic Dark Earth soil samples were collected from 5 points (duplicate) in a native archaeological site of the Lower Amazon, Brazil (Figure 1).
Composited samples were collected at a depth of 0 to 20 cm. The soils were stored in sterile Nasco® plastic in 4°C bags for further use.
DNA extraction and PCR-DGGE
Sample (approximately 0.25 g soil wet weight) of soil was used for DNA extraction by using a NucleoSpin Tissue kit (Macherey-Nagel, Düren, Germany). Total DNA was used for PCR amplification of prokaryote and eukaryote ribosomal target regions, for PCR-DGGE analyses. Two primers sets were used for the microbial population. Primers and analysis conditions are given Table 1.
Bands from the PCR-DGGE gels were excised and were reamplified using the same primers for prokaryote and eukaryote (Table 1). The amplicons were sequenced by Macrogen Inc. (Seoul, South Korea). GenBank searches
(http://www.ncbi.nlm.nih.gov/BLAST/) were performed to determine the species of the obtained sequences. The PCR-DGGE gels were analyzed for Bio-Numerics software (version 1,5, Applied Maths, Kortrijk, Belgium) for determining the amplicons diversity.
Physico-chemical analysis of the soil
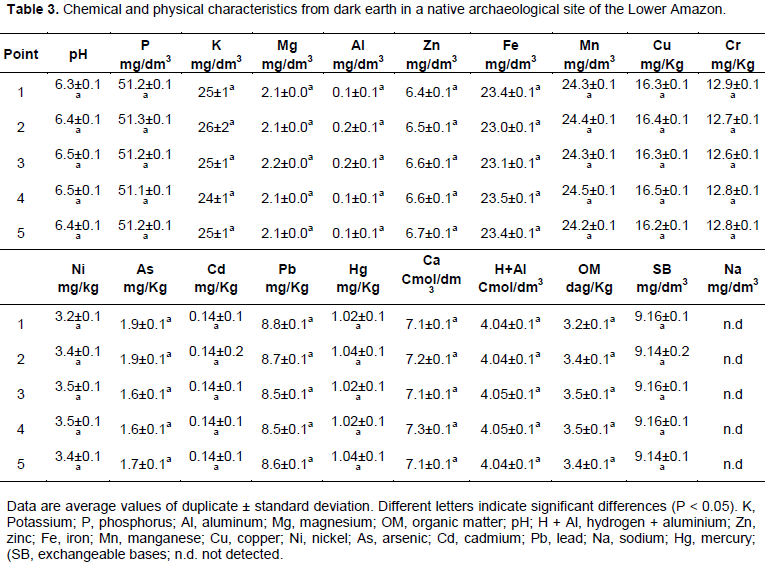
The physical and chemical characteristics of Amazon soil from dark earth in a native archaeological site of the Lower Amazon were analyzed. The soils were analyzed in duplicate according to Embrapa (1997). The followings were determined: The value of pH, concentration of hydrogen + aluminium (H + Al), calculation of exchangeable bases (SB), zinc (Zn), iron (Fe), manganese (Mn), copper (Cu), nickel (Ni), arsenic (As), potassium (K), phosphorus (P), aluminum (Al), magnesium (Mg), organic matter (OM), cadmium (Cd), lead (Pb), sodium (Na) and mercury (Hg). For statistical analysis, the SAS System 9.1 (SAS Institute Inc., Cary, NC, USA) was used.
Use of different primers to assess eukaryote and prokaryote communities from dark earth in a native archaeological site of the Lower Amazon
The bacterial and fungal DGGE profiles of Amazon dark earth in a native archaeological site of the Lower Amazon are shown in Figure 2. These profiles exhibited the species abundance. For eukaryote, primer pair ITS1fGC/ITS4r and NS3fGC/YM951r were able to provide a diversity of bands and to differentiate filamentous fungi (Penicillium and Cladosporium) and yeasts (Saccharomyces, Zygosaccharomyces, Candida and Lachancea).
For eukaryote, the primers 338fGC/518r and 968fGC/1401r were able to demonstrate variety of bands with the appearance of Bacillus, Klebsiella, Enterobacter, Pantoea, Escherichia, Lactobacillus, Leuconostoc and actinobacterias.
The obtained results using different pairs of primers, show a diverse PCR-DGGE profile, suggesting the presence microbial consortium. This has high relevance in terms of the efficiency of the characterization of microbial diversity of Amazon dark earth in a native archaeological site of the Lower Amazon.
Identification of microbial communities from dark earth in a native archaeological site of the Lower Amazon
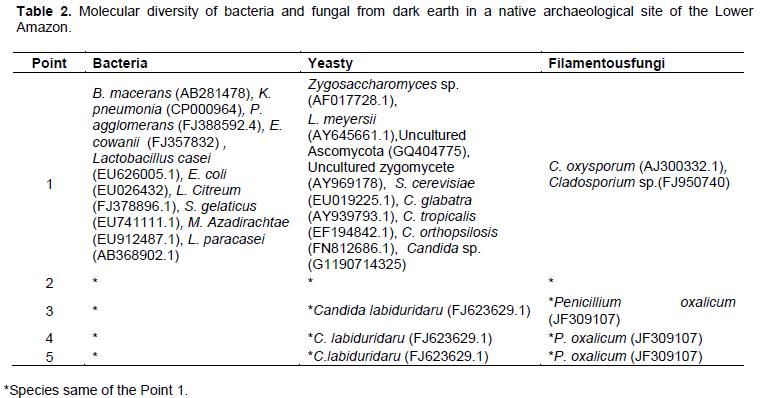
Table 2 and Figure 2 showed the molecular diversity of bacterial and fungal from dark earth in a native archaeological site of the Lower Amazon. Identical profiles were obtained for the following primers pairs: Prokaryote (968fGC/1401r and 338fGC/518r) and eukaryote (ITS1fGC/ITS4 and NS3fGC/YM951r). The sequencing bands exhibited equal and higher than 98% identity with sequences available in the Gen-Bank.
In relation to the prokaryotic community analysis, the bands a to j (Figure 2a) were identified as different bacteria species. The band a was identified as Bacillus macerans (AB281478), band b - Klebsiella pneumonia (CP000964), band c - Pantoea agglomerans (FJ388592.4),, band d -Enterobacter cowanii (FJ357832), band e - Lactobacillus casei (EU626005.1), band f - Escherichia coli (EU026432), band g - Leuconostoc citreum (FJ378896.1), band h - Streptomyces gelaticus (EU741111.1) ,band i - Microbacterium azadirachtae (EU912487.1) and band j as Lactobacillus paracasei (AB368902.1).
In relation to the eukaryotic community analysis (Figure 2b), the band k was identified as Zygosaccharomyces sp. (AF017728.1), band l - Lachancea meyersii (AY645661.1), band m -Uncultured Ascomycota (GQ404775), band n - Uncultured zygomycete (AY969178), band o - Saccharomyces cerevisiae (EU019225.1), band p - Cladosporium oxysporum (AJ300332.1), band q - Cladosporium sp. (FJ950740), band r - Candida glabatra (AY939793.1), band s - Candida tropicalis (EF194842.1), band t - Candida orthopsilosis (FN812686.1),band u – Candida sp. (G1190714325), band v - Penicillium oxalicum (JF309107), and band x as Candida labiduridaru (FJ623629.1).
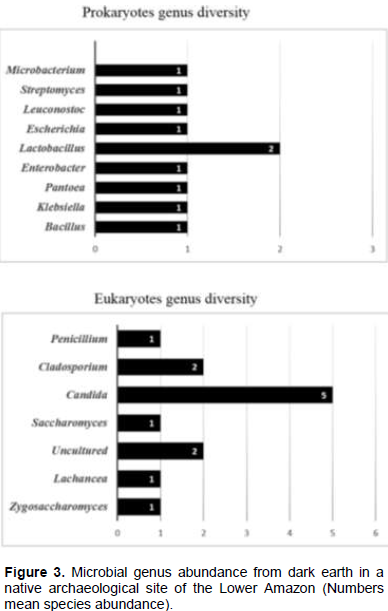
C. labiduridaru (FJ623629.1) yeast and P. oxalicum (JF309107) filamentous fungi were found in only samples of the points 3, 4 and 5 numbers. Figure 3 describes the microbial genus abundance in dark earth in a native archaeological site of the Lower Amazon. The bacterial genus of greater abundance was Lactobacillus and the fungal genus of greater abundance was Candida.
Physico-chemical properties from dark earth in a native archaeological site of the Lower Amazon
Chemical and biochemical properties of dark earth in a native archaeological site of the Lower Amazon are given in Table 3. The soil pH value was close to neutral (~6.5). The soil had high levels of minerals with exception of Na and Al (Table 3). The organic matter was found as 3.5 dag/kg.
Studies of microbiota and soil chemistry show that the fertility of anthropogenic soils results from combination of mineral and organic components (Navarrete et al., 2010; Taketani et al., 2013). The microbial population profiles can be dependent on the PCR primers used (Anderson and Cairney, 2004; Schwarzenbach et al., 2007). When Amazon soil samples were subjected to analysis by PCR-DGGE, important information became available. Species present in low concentrations can be detected by PCR-DGGE (Pereira et al., 2011).
The results show that the eukaryote population in the Amazon soils reveals a microbial diversity, Aspergillus sp., Penicillium sp., Trichoderma sp., Zygosaccharomyces sp. Lachancea sp., Saccharomyces sp., Cladosporium sp., and Candida sp. The large variability is due the incorporating of different types and quantities of organic matter (Bissett et al., 2013). Recently, Bresolin et al. (2010) observed microbiota present in Brazilian Cerrado soil using DNA analysis by PCR-DGGE. The microbial species isolated were related to Uncultured soil fungus and Uncultured soil bacteria. Schwarzenbach et al. (2007) observed the presence of Ascomycota yeasts in sandy loam soil sample situated in Central Switzerland.
Among the genus and species of yeast and fungi found in this study; Lachancae meyersii is the first species in the collected genus from Amazon soil. According to Fell et al. (2004), all of the isolations of the other species of Lachancea have been from plants, plant products or plant-associated insects, fruits or food. The specific ecological niche of L. meyersii has not been determined. Zygosaccharomyces is a yeast genus as synonymous with spoilage. Zygosaccharomyces includes osmotolerant (Thomas and Davenport, 1985). These characteristics of resistant microorganisms to different environments are related to presence in soils. The Cladosporium genus constitutes one of the largest genus of Hyphomycetes (Mukherjee and Mittal, 2005). Cladosporium grows when there is not enough ventilation; sometimes on walls and wallpaper in rooms (Mukherjee and Mittal, 2005). However, recent data suggest that they are present in soil samples (Paul et al., 2008). The Candida genus is commonly found in soils in forest soil in Taiwan including the new species Candida jianshihensis, Candida yuanshanicus, Candida dajiaensis and Candida sanyiensis (Meyer et al., 1998). Shin et al. (2001) also found Candida genus in Korean soils.
The obtained results showed the presence of several bacterial species in Amazon soil (Figures 2 and 3 and Table 2). The bacterial species Bacillus sp. commonly
are found in soil samples (Quirino et al., 2009).
Klebsiella, Enterobacter, Pantoea, and Escherichia genera were found in this study. These genera are found naturally in soil, water, and plants (Quirino et al., 2009). Leuconostoc and Lactobacillus genera are not commonly found in soil, however it was found in this study. The presence of this specie in soil is associated with plants and fruits present in vegetation Amazon.
This study also showed the presence of actinobacterias as Streptomyces gelaticus (EU741111.1) and Microbacterium azadirachtae (EU912487.1). Actinobacteria are genus colonizers in soils. Many species produce enzymes for degradation of cellulose, chitin and, in part, starch. Actinobacteria often occur in degraded organic materials (Schäfer et al., 2010). In the present work, the Streptomyces genus was also found. The Streptomyces genus is focus of research because of the produced substances and has been modified with advances in molecular biology (Souza et al., 2008). Microbacterium genus can be isolated from air, soil and water. Many Microbacterium spp. play a significant role in oil, lactone and xylan degradation, production of bio-surfactants, and as a growth promoter in plants (Lin et al., 2012).
The soil physico-chemical properties are important and how these properties could be related to microbial profiles in different soils must be evaluated (Peixoto et al., 2010). The properties affect the native microbial populations (Bresolin et al., 2010). This study showed small amounts of aluminum in the Amazon soil (Table 3). High quantities of aluminum in the soil promote the soil impoverishment (Ruggiero et al., 2002). This study also showed the Lower Amazon soil is rich in minerals. This can be an important factor in the species richness in the Amazon region.
In summary, this research has furthered our knowledge about microbial community structure and chemical composition of archaeological site of the Lower Amazon. The application of PCR-DGGE technique based approaches for prokaryote and eukaryote population analysis has confirmed that microbial ecosystems of Amazon soil support a wide diversity of microorganisms that may be responsible for some characteristics these soils. On the other hand, molecular approaches revealed microbial groups that had never been reported in native archaeological site of the Lower Amazon as the Leuconostoc mesenteroides, Lactobacillus casei and Lactobacillus paracasei bacteria’s and L. meyersii yeast.
The data presented adds important information that will help future studies in these environments.
The authors have not declared any conflict of interests.
The authors acknowledge the CNPq, CAPES, FAPEAM and FAPEMIG for financial support.
ITS, Internal transcribed spacer, PCR-DGGE, polymerase chain reaction -denaturing gradient gel electrophoresis.
REFERENCES
|
Anderson IC, Cairney JW (2004). Diversity and ecology of soil fungal communities: increased understanding through the application of molecular techniques. Environ. Microbiol. 6:769-779.
Crossref
|
|
|
|
Ascher J, Ceccherini MT, Chro-áková A, Jirout J, Borgogni F, Elhottová D, Simek M, Pietramellara G (2010). World evaluation of the denaturing gradient gel electrophoresis apparatus as a parameter influencing soil microbial community fingerprinting. J. Microbiol. Biotechnol. 26:1721-1726.
Crossref
|
|
|
|
|
Bissett A, Brown MV, Siciliano SD, Thrall PH (2013). Microbial community responses to anthropogenically induced environmental change: towards a systems approach. Ecol. Letters. 16:128-139.
Crossref
|
|
|
|
|
Bossio DA, Girvan MS, Verchot L, Bullimore J, Borelli T, Albrecht A, Scow KM, Ball AS, Pretty JN, Osborn AM (2005). Soil Microbial Community Response to Land Use Change in an Agricultural Landscape of Western Kenya. Microb. Ecol. 49:50-62.
Crossref
|
|
|
|
|
Bresolin JD, Bustamante MMC, Krüger RH, Silva MRSS, Perez KS (2010). Structure and composition of bacterial and fungal community in soil under soybean monoculture in the Brazilian Cerrado. Brazilian J. Microbiol. 41:391-403.
Crossref
|
|
|
|
|
Brossi MJd-L, Mendes LW, Germano MG, Lima AB, Tsai SM (2014). Assessment of Bacterial bph Gene in Amazonian Dark Earth and Their Adjacent Soils. PLoS ONE. 9:95-97.
|
|
|
|
|
Castro AP, Quirino BF, Pappas G Jr, Kurokawa AS, Neto EL, Krüger RH (2008). Diversity of soil fungal communities of Cerrado and its closely surrounding agriculture fields. Arch. Microbiol. 190:129-139.
Crossref
|
|
|
|
|
Embrapa: Empresa Brasileira de Pesquisa Agropecuária (1997). Manual de métodos de análise de solos. 2. ed. Rio de Janeiro, P 212.
|
|
|
|
|
Fell JW, Statzell–Tallman A, Kurtzman CP (2004). Lachanceameyersii sp. nov., an ascosporogenous yeast from mangrove regions in the Bahama Islands. Studies Mycol. 50:359-363.
|
|
|
|
|
Kim JS, Sparovek G, Longo RM, De Melo WJ, Crowley D (2007). Bacterial diversity of terra preta and pristine forest soil from the Western Amazon. Soil Biol. Biochem. 39:684-690.
Crossref
|
|
|
|
|
Lehmann J, Silva Jr. JP, Steiner C, Nehls T, Zech W, Glaser B (2003). Nutrient availability and leaching in an archaeological Anthrosol and a Ferralsol of the Central Amazon basin: fertilizer, manure and charcoal amendments. Plant Soil. 249:343-357.
Crossref
|
|
|
|
|
Lin L, Guo W, Xing Y, Zhang X, Li Z, Hu C, Li S, Li Y, An Q (2012). The Actinobacterium Microbacterium sp. 16SH accepts pBBR1-based pPROBE vectors, forms biofilms, invades roots, and fixes N2 associated with micropropagated sugarcane plants. Appl. Microbiol. Biotechnol. 93:1185-1195.
Crossref
|
|
|
|
|
Magalhães KT, Pereira GVM, Dias DR, Schwan RF (2010). Microbial communities and chemical changes during fermentation of sugary Brazilian kefir. World J. Microbiol. Biotechnol. 26:1241-1250.
Crossref
|
|
|
|
|
Meyer SA, Payne RW, Yarrow D (1998). Candida Berkhout. In 'The Yeasts, a Taxonomic Study'. (EdsCP Kurtzman, JW Fell) pp454-573. Elsevier: Amsterdam.
Crossref
|
|
|
|
|
Mukherjee I, Mittal A (2005). Bioremediation of Endosulfan Using Aspergillusterreus and Cladosporium oxysporum. Environ. Contamin. Toxicol. 75:1034-1040.
Crossref
|
|
|
|
|
Navarrete AA, Cannavan FS, Taketani RG, Tsai SM (2010). A molecular survey of the diversity of microbial communities in different Amazonian agricultural model systems. Diversity 2:787-809.
Crossref
|
|
|
|
|
Paul NC, Yu SH (2008). Two Species of Endophytic Cladosporium in Pine Trees in Korea. Mycobiol. 36:211-216.
Crossref
|
|
|
|
|
Peixoto RS, Chaer GM, Franco N, Reis Junior FB, Mendes IC, Rosado AS (2010). Decade of land use contributes to changes in chemistry, biochemistry and bacterial community structures of soils in the Cerrado. Ant. Van. Leeuw. 98:403-413.
Crossref
|
|
|
|
|
Pereira GVM, Magalhães KT, Lorenzetii ER, Souza TP, Schwan RS (2011). A Multiphasic Approach for the Identification of Endophytic Bacterial in Strawberry Fruit and their Potential for Plant Growth Promotion. Microbiol. Ecol. 63:405-417.
Crossref
|
|
|
|
|
Pereira RM, Silveira EL, Scaquitto DC, Pedrinho EAN, Val-Moraes SP, Wickert E, Carareto-Alves LM, Lemos EGM (2006). Molecular Characterization of bacterial populations of different soils. Brazil. J. Microbiol. 37:439-447.
|
|
|
|
|
Quirino B, Pappas GJ, Tagliaferro AC, Collevatti RG, Neto EL, da Silva MR, Bustamante MM, Krüger RH (2009). Molecular phylogenetic diversity of bacteria associated with soil of the savanna-like Cerrado vegetation. Microbiol. Res. 64:59-70.
Crossref
|
|
|
|
|
Ruggiero PGC, Batalha MA, Pivello VR, Meirelles ST (2002). Soil-vegetation relationships in cerrado (Brazilian savanna) and semi-deciduous forest, Southeas thern Brazil. Plant Ecol. 160:1-16.
Crossref
|
|
|
|
|
Schäfer J, Jäckel U, andKämpfer P (2010). Development of a new PCR primer system for selective ampli¢cation of Actinobacteria. FEMS Microbiol. Letters. 311:103-112.
Crossref
|
|
|
|
|
Schwarzenbach K, Enkerli J, Widmer F (2007). Objective criteria to assess representatively of soil fungal community profiles. J. Microbiol. Methods. 68:358-366.
Crossref
|
|
|
|
|
Shin KS, Shin YK, Yoon JH, Park YH (2001). Candida Thermophila sp. nov., a novel thermophilic yeast isolated from soil. Inter. J. Syst. Evolut. Microbiol. 51:2167-2170.
Crossref
|
|
|
|
|
Silva MS, Alenir NS, Magalhães KT, Dias DR, Schwan RF (2012). Brazilian Cerrado Soil Actinobacteria Ecology. Bio-Med. Res. Inter. pp. 1-10.
|
|
|
|
|
Souza RF, Coelho RR, Macrae A, Soares RM, Nery Dda C, Semêdo LT, Alviano CS, Gomes RC (2008). Streptomyces lunalinharesii sp. nov., a Chitinolytic streptomycete isolated from cerrado soil in Brazil. Inter. J. Syst. Evolut. Microbiol. 58:2774-2778.
Crossref
|
|
|
|
|
Taketani RG, Lima AB, Jesus EC, Teixeira WG, Tiedje JM, Tsai SM (2013). Bacterial community composition of anthropogenic biochar and Amazonian anthrosols assessed by 16S rRNA gene 454 pyrosequencing. Ant. Van. Leeuw. 104:233-242.
Crossref
|
|
|
|
|
Thomas DS, Davenport RR (1985). Zygosaccharomyces bailii: a profile of characteristics and spoilage activities. Food Microbiol. 2:157-169.
Crossref
|
|
|
|
|
Wallis PD, Haynes RJ, Hunter CH, Morris CD (2010). Effect of land use and management on soil bacterial biodiversity as measured by PCR-DGGE. Appl. Soil Ecol. 46:147-150.
Crossref
|
|