Full Length Research Paper

ABSTRACT
Leaf blight caused by Rhizoctonia solani is one of the most serious diseases that cause damage to turmeric crop. Fifty percent disease incidence was observed in turmeric growing areas of Andhiyur (Erode district) and Mettur (Salem district) of Tamil Nadu, India. The antifungal efficacy of botanicals, bioagents and fungicides were tested against R. solani in vitro. It was observed that seed extract and oil cake of Madhuca longifolia suppressed the mycelia growth (34.81; 49.63%), sclerotial formation (100%) and production of biomass (2.18; 2.41 mg) of R. solani followed by leaf extracts of Azadirachta indica and Littorina littorea. Among the bioagents native isolate of BSoya1 of Bacillus spp., Cg1 of Chaetomium spp., Pf1 of Pseudomonas spp. and Tv1 of Trichoderma spp. shown more than 50% antagonistic effect on mycelia growth and 100% on sclerotial formation. The complete fungal growth inhibition was observed in Nativo and Bavistin fungicides amended medium. Then, the basal soil application of mahua cake and T. viride in the ratio of 1:2 (1 g of mahua cake mixed with 2 g T. viride per kg of pot soil) followed by foliar spray of Nativo @ 0.5% significantly mitigated the leaf blight (8.6%) over the inoculated control (50.51%) in pot culture experiments.
Key words: Turmeric, efficacy, fungicides, bioagents, botanicals, Rhizoctonia solani.
INTRODUCTION
Turmeric (Curcuma longa L.) known as the “golden spice”, is one of the most important herbs in tropical and subtropical countries. It is a rhizomatous perennial plant of the ginger family, Zingiberaceae (Dixit et al., 2002). Turmeric is used as condiment, dye, drug and cosmetic in addition in religious ceremonies (Gescher et al., 2005). India is the leading producer, consumer and exporter of turmeric in the world and contributes about 80% of the world production and 60% to the total trade (Parthasarthy et al., 2007).
Several fungal, bacterial and viral diseases are reported in turmeric (Hudge and Ghugul, 2010). Among the diseases, leaf blight caused by Rhizoctonia solani is an emerging disease to the turmeric crop (Roy, 1992). The pathogen is known to be soil borne and sclerotia are often found in the soil. Limited information is available on sustainable management and the disease control is generally with chemical applications. The increasing awareness of fungicide related hazards has emphasized the need for adopting biological methods as an alternative disease control method, which is also ecofriendly (Khare et al., 2010).
Biological control appears to be the best solution for long term sustainability and effective management of soil borne disease which can considerably minimize the disease. Successful management of R. solani on various crops by bioagents has been previously reported (Lahlali and Hijri, 2010; Seema and Devaki, 2012; Srinivas et al., 2014).
Hence, considering economic importance of the crop and the disease, the present investigation was under-taken to conduct the disease survey, to evaluate the efficacy of fungicide, botanicals and bioagents against R. solani and to find out the suitable management practice to mitigate the disease.
MATERIALS AND METHODS
Survey
Turmeric growing villages of Tamil Nadu, India were surveyed to assess the leaf blight disease incidence and severity on turmeric. The disease scoring was done in 50 plants at three different locations in a single village. The disease rating scale (0 to 9) was followed for the assessment of incidence and severity of leaf blight disease. The blight disease incidence was assessed and expressed in percent disease index (PDI) and also relative lesion height (RLH) was assessed based on the lesion height (Sharma et al., 1990). The following formula was used to calculate PDI.

Standard evaluation system for screening rice sheath blight (0-9 scale) is shown in Table 1.

The following formula was used to calculate relative lesion height (RLH):

Isolation of pathogen
R. solani was isolated from the naturally infected leaf samples through tissue segment method collected from different places of Tamil Nadu. Small portions of sheath showing typical lesions were cut into small bits of 0.5 - 1 cm, surface sterilized with 0.1% mercuric chloride and rinsed three times with sterile distilled water. Then, they were transferred to Petri dish containing potato dextrose agar (PDA) medium aseptically (Riker and Riker, 1936). The pure culture of the pathogen was obtained by single hyphal tip technique and the culture was maintained on PDA slants to carry out further studies (Taheri et al., 2007).
Pathogenicity tests
Diseases free (healthy) turmeric rhizomes (Erode local 8) were planted in 30 cm diameter earthen pots containing pot mixture at two rhizomes per pot. Actively growing plants at 30 days after planting (DAP) were inoculated with R. solani by inserting young immature sclerotia (two sclerotia per sheath) and incubated for seven days for the development of typical blight symptoms on the plants (Sriram et al., 1997).
In vitro effect of botanicals on radial mycelial growth of R. solani
Leaves of nine plants viz., Lawsonia inermis L. (Maruthani), Ocimum tenuiflorum L. (Thulasi), Azadirachta indica L. (Neem), Morinda citrifolia L. (Noni), Vinca major (Periwinkle), Gloriosa suberba (Glory lily), Justicia adhatoda (Adathoda), Vitex nigundo (Nochi) and Madhuca longifolia (Mahuva seed extract) and seven oilcakes viz., mahuva (Mahua longifolia L.), pungam (Pongamia glabra L.), sesamum (Sesamum indicum L.), groundnut (Arachis hypogaea L.), castor (Ricinus communis L.), neem (Azadirachta indica L.) and sunflower (Helianthus annuus L.) were collected and tested for their efficacy against R. solani by poisoned food technique. Fresh plant leaves and seed extracts were prepared by cold water extraction method (Shekhawat and Prasad, 1971). The leaves were first washed with distilled water and finally with sterile water and the oilcakes were first soaked in sterile distilled water at the rate of one g in 1 ml of water and kept overnight. Then, ground in pestle and mortar by adding sterile water at the ratio of 1:1 (w/v). The macerate was squeezed using cotton to get the extract. The extract was strained through two layers of muslin cloth and finally through Whatman No.1 filter paper and this formed the standard plant extract solution (100%). This was further diluted with sterilized distilled water (v/v) to have the required concentrations (10, 15, and 20%). The PDA medium was mixed with different concentrations viz., 10, 15 and 20% of plant extracts and oil cakes. Mycelial plug of the pathogens (7 mm) was placed at the centre of each Petri plate and incubated at 28 ± 1°C and three replications were maintained for each treatment. The experiment was arranged in a completely randomized block design. The fungus grown on PDA without any plant extracts and oil cakes served as control. The growth diameter was recorded and the percent inhibition was calculated.
In vitro efficacy of botanicals on biomass production of R. solani
The effect of plant products on mycelia growth of the pathogen in liquid medium was done by the method suggested by Neslihan et al. (2008). Different concentrations (10, 15, and 20%) of plant extracts and oil cakes were amended with the 100 ml of potato dextrose broth. A mycelial plug (7 mm) was transferred to the flasks which contain different concentrations of plant extracts and incubated at 28±2°C for one week. Three replications were maintained for each treatment. The experiment was arranged in a completely randomized block design.The fungal mycelial mat was collected, dried and mycelium weight recorded.
In vitro efficacy of bioagents on mycelial growth and sclerotial formation of R. solani
The native isolates of bio-agents viz., Bacillus isolates (EPCO8, EPCO44, EPCO26, BG1, Bsoya1, Bsoya2, BG2, BC3, EPCO81, EPCO PF, BC1, EPCO78, BC9, BC8, BC7, BC6, EPCO P6); Pseudomonas isolates (Pf1, KAU46, APF6, Pf3, PC3, 18P, 6P, PC4 and Pf26); Chaetomium isolates (Cg1, Cg2, Cg3, Cg4, Cg5, Cg6 and Cg249) and Trichoderma viride isolates (Tv1, Tv2, Tv3, Tv4, Tv5, Tv6 and Tv7) were used for testing their efficacy against R. solani by dual plate technique (Dennis and Webster, 1971). The bioagents as well as the pathogen were inoculated equidistant on PDA medium aseptically and incubated at 28 ± 1°C. In each case, three replications were maintained for each isolates along with control. The experiment was arranged in a completely randomized block design. After obtaining the maximum growth in the control, the observation on radial growth of pathogen was taken and percent inhibition was worked out using Vincent’s formula. The sclerotial formation was recorded on 7th and 14th days after inocu-lation.
In vitro efficacy of bioagents on biomass production of R. solani
The effect of culture filtrates of the native isolates of bioagents viz., Bacillus isolates (EPCO8, EPCO44, EPCO26, BG1, Bsoya1, Bsoya2, BG2, BC3, EPCO81, EPCO PF, BC1, EPCO78, BC9, BC8, BC7, BC6, EPCO P6) ; Pseudomonas isolates (Pf1, KAU46, APF6, Pf3, PC3, 18P, 6P, PC4 and Pf26); Chaetomium isolates (Cg1, Cg2, Cg3, Cg4, Cg5, Cg6 and Cg249) and Trichoderma viride isolates (Tv1, Tv2, Tv3, Tv4, Tv5, Tv6 and Tv7) on the biomass production of R. solani was studied as per method given by Dennis and Webster (1971). Sterilized potato dextrose (PD) broth (100ml) was taken in 250 ml flask and inoculated with mycelial plugs (7 mm) of the biocontrol native isolates taken from the edge of four day old culture. Inoculated flasks were incubated at 28 ± 2°C for one week and the cultural filtrate was extracted by centrifuging the content at 10000 rpm for 30 min and the culture filtrate was collected. The culture filtrate of bioagents were mixed with PD broth at a rate of 1:4 (v/v) and inoculated with mycelial plugs of pathogen (7 mm), then incubated at 28±2°C for seven days. Three replications were maintained for each isolates. The experiment was arranged in a completely randomized block design. After the incubation period, fungal mycelial mat was collected and dried. The dried mycelial weight was recorded.
In vitro efficacy of fungicides on mycelial growth of R. solani
Evaluation of fungicides was carried out for their efficacy to inhibit the mycelial growth of R. solani isolate by “poisoned food technique” as described by Sharvelle (1961). The fungicides viz., azoxystrobin (Mirador 250 SC), tebuconazole (Orius 25.9% EC), tridemorph (Calixin 80% EC), fosetyl-Aluminium (Aliette WP 80), trifloxystrobin (Flint 50 WG), fenamidone 10% + mancozeb 50% (Sectin 60 WG), Carbendazim (Bavistin 50WP), tebuconazole 50% + trifloxystorbin 25% (Nativo WG 75) were used at 10, 50, 100, 250, 500, 750 and 1000 ppm concentration. Mycelial plugs of the pathogens (7 mm) taken from a seven day old culture was placed at the centre of each Petri plate and incubated at 28 ± 1°C. Three replications were maintained for each treatment. The experiment was arranged in a completely randomized block design. The fungus grown on PDA without any fungicides served as control. The radial growth of the colony was recorded when maximum growth was observed in the control and it was calculated by using the following formula.
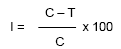
Where, I = Percent inhibition; C = radial growth in control; T = radial growth in treatment.
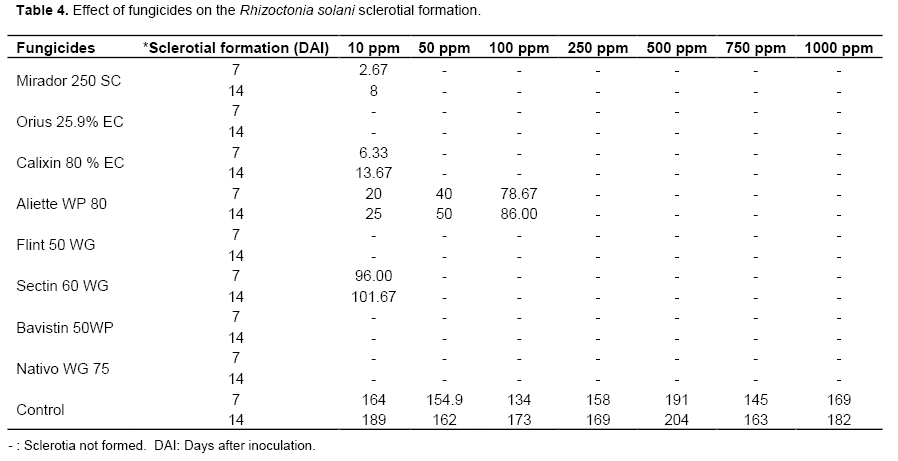
The sclerotial formation was recorded on 7th and 14th days after inoculation.
Management of leaf blight of turmeric incited by R. solani – Greenhouse studies
Studies were conducted to test the efficacy of effective treatment against leaf blight disease of turmeric under greenhouse condition with nine treatments with fungicide, bioagents and botanical. The Erode local 8 was sown in the pots containing 5 kg/pot of sterilized soil. The pathogen was inoculated 30 DAP. Three replications were maintained for each treatment. The experiment was arranged in a completely randomized block design. The blight incidence was recorded on 40 days after sowing and the effectiveness of the treatments on the intensity of leaf blight disease was observed seven days after inoculation, with a 0-9 scale of the Standard Evaluation System for rice, IRRI (2002) and expressed as percent disease index (PDI).
RESULTS AND PLUGUSSION
Survey and pathogenicity studies of R. solani in turmeric
Among the fifteen villages surveyed, the turmeric plants at Andhiyur, Erode district recorded the maximum of 54.5% disease index (PDI) of leaf blight which was followed by Mettur in Salem district which recorded 44.3 PDI. The disease incidence was prevalent in only four villages viz., Annur (Rs1), Andhiyur (Rs2), Mettur (Rs3) and Parama-thivellur (Rs4) and the incidence ranged from 6.4 to 54.5 PDI (Table 2). The results revealed that placing of sclerotia in the injured sheath region exhibited the maximum lesion length of 16.89 cm and leaf blight lesion length. The symptom observed under field condition was very typical when compared with the artificially inoculated plants. Though the symptoms produced by all the isolate were similar, the most virulent Andhiyur, Erode isolate was highly virulent as compared to other isolates and produced larger lesion length.
.png)
Efficacy of fungicides on turmeric leaf blight pathogen (R. solani)
Nativo and Bavistin were individually effective against the pathogen even at the lowest concentration of 10 ppm by completely inhibiting the mycelia growth and sclerotia production. It was significantly superior over other fungicides and on par with each other. It was followed by Mirador, Orius and Calixin which exhibited total inhibition at 50 ppm (Table 3). The least effective fungicides were Aliette, Flint and Sectin (48.15% inhibition at 1000 ppm). The same trend was observed in the liquid culture technique in which the two fungicides viz., Nativo and Bavistin completely inhibited the mycelia growth at the lowest concentration of 10 ppm (Table 3). Aliette had recorded sclerotial formation of 86 numbers on 14 DAI at 100 ppm. As the concentration of fungicides increased, the sclerotial formation was arrested (Table 4).
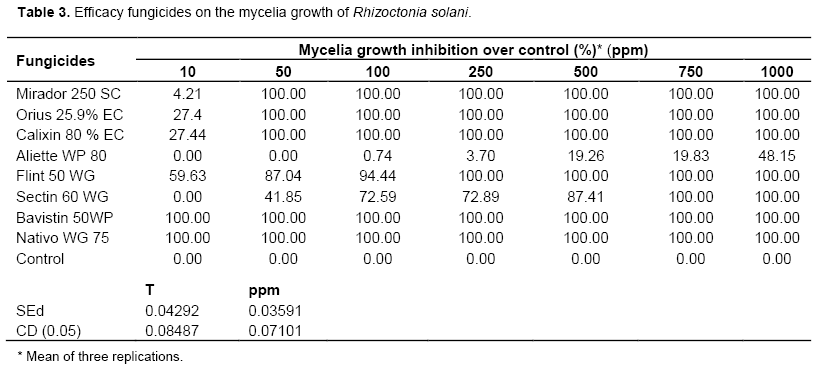
Fungicides viz., trifloxystrobin + tebuconazole, tebuconazole and propiconazole showed higher level of efficacy against R. solani of rice in laboratory conditions (Hunjan et al., 2011). Among the new formulations, Filia and Nativo were equally effective in controlling sheath blight (Swamy et al., 2009). In the present study, among different fungicides screened for, R. solani Nativo and Bavistin were individually effective against the pathogen in inhibiting the mycelia growth and sclerotial production even at the lowest concentration of 10 ppm.
In vitro antagonistic effect of bioagents against R. solani
Bacillus sp.
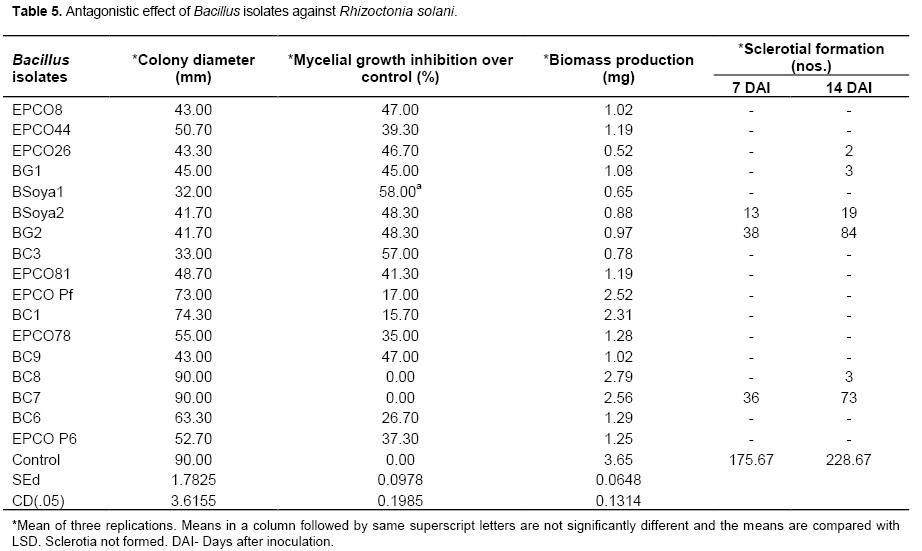
Bacillus isolates, BSoya1 and BC3 respectively recorded 32 and 33 mm colony diameter which accounted for 58 and 57% mycelia growth inhibition over the control. BSoya1 and BC3 Bacillus isolates were significantly superior and on par with each other in controlling the mycelia growth of R. solani. It was interesting to note that the sclerotial formation was completely inhibited in eleven isolates. The remaining six isolates of Bacillus viz., EPCO26, BG1, BSoya2, BG2, BC7 and BC8 had sclerotial formation. Among the seventeen isolates a maximum number of 84 sclerotia were observed in the BG2 as against the control which recorded 228.67 sclerotia (Table 5). All the Bacillus isolates reduced the mycelial weight of the pathogen over the control. The least biomass production was seen with the Bacillus isolate EPCO26 (0.52 mg) followed by Bsoya1 (0.65 mg) and BC3 (0.78 mg) (Table 6). Similar antagonistic effect of Bacillus spp. against R. solani infecting many other crops was reported by several workers. Calvo et al. (2010) reported that, Bacillus spp. from rhizosphere region of potato showed high antagonistic effect against R. solani causing various diseases in crops plants. Padaria and Kapoor (2011) reported that, Bacillus pumillus obtained from rice field exhibited antagonistic activity against R. solani infecting various crops. Bacillus sp. strain 916, isolated from the soil showed strong activity against R. solani causing rice sheath blight (Wang et al., 2012). Bacillus sp. shows inhibition against the growth of a wide range of plant fungal pathogens like R. solani and Fusarium verticillioides by producing morphological modifications on the pathogenic fungi hyphae like total collapse of the cell. In the present study, among the 17 Bacillus isolates, BSoya1 and BC3 were significantly superior over other antagonists and it also reduced the mycelial weight and sclerotial production of the pathogen.
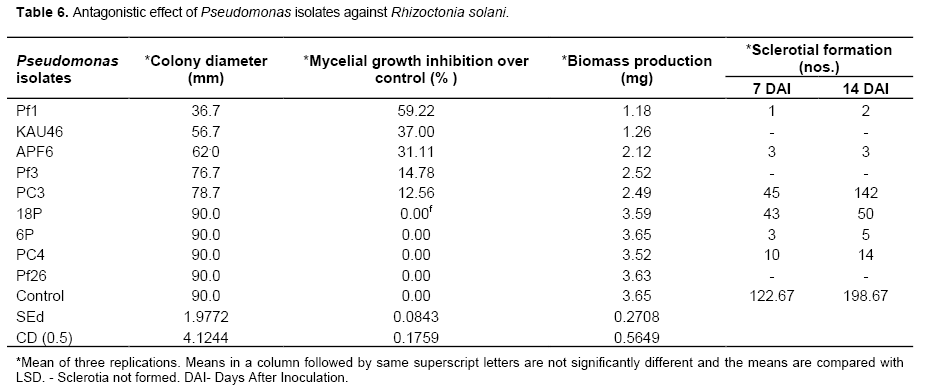
Pseudomonas spp.
Among nine isolates of Pseudomonas spp., Pf1 strain was significantly superior and recorded 59.22% mycelial growth inhibition and 1.18 mg biomass production. This was followed by KAU46 which recorded 56.7 mm colony diameter which accounted for 37% mycelial growth inhibition over control (Table 6). After 14 days of inoculation, the sclerotial count was recorded and it was observed that except three Pseudomonas strains viz., KAU46, Pf3 and Pf26, all the other strains supported sclerotia formation of R. solani but the production was drastically reduced. Among the various strains of Pseudomonas, the maximum number of 142 sclerotia was recorded in PC3 strain as against the control which recorded 198.67 numbers of sclerotia. Though the Pseudomonas isolate KAU 46 exhibited 56.7 mm mycelial growth, it had completely inhibited the sclerotial formation in R. solani. Pf 26 has recorded full growth of the test pathogen but it had also inhibited the sclerotial formation (Table 7). Hence the mycelial growth could not be correlated with the sclerotial formation or inhibition. Similarly, Zachow et al. (2010) found that P. fluorescens L13-6-12 applied to the R. solani infected soil in sugar beet, formed large microcolonies consisting of hundreds of cells.
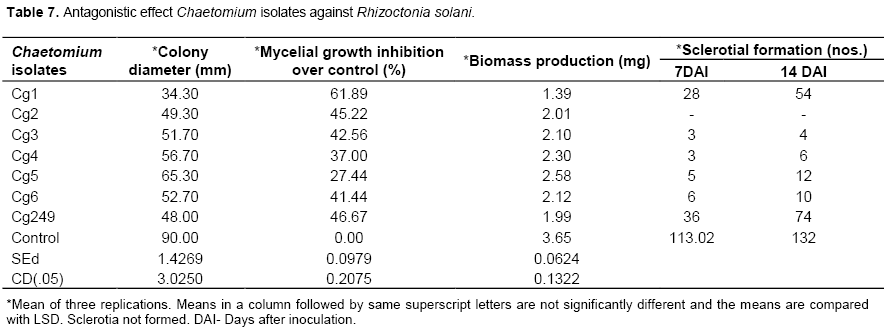
Chaetomium sp.
On studying the interaction between the Chaetomium and the pathogen, the isolate Cg1 showed the maximum percent inhibition (61.89%) which was significantly higher than other isolates (Table 6). It was followed by Cg249 (46.67%) and Cg2 (45.22%) and they were on par with each other. The reduction in mycelial biomass was also recorded with Cg1 (1.39 mg) and the isolate was individually significantly superior when compared with other isolates. It was followed by Cg249 (1.99 mg) and Cg2 (2.01 mg). All the isolates drastically reduced the number of sclerotial formation over control. The isolates Cg2 completely inhibited the sclerotial formation. The maximum sclerotial production was recorded in the control (132 numbers), where as the minimum sclerotial production was seen in the isolate Cg 3 (4 numbers) (Table 7). Kaushik et al. (2010) reported that Chaetomium globosum showed good activity against Sclerotinia sclerotiorum, Fusarium oxysporum and R. solani.
Trichoderma viride
Among the seven isolates of Trichoderma spp. tested, maximum percentage of inhibition was recorded with the isolate Tv1. All the seven isolates inhibited the growth of R. solani in dual culture. The isolate Tv1 showed maximum percent inhibition of mycelial growth (78.11%) and it was individually significantly superior, followed by Tv5 (61.11%) and Tv3 (57.00%). The least percent inhibition was observed with the isolate T. harzianum (44.78%) (Table 8). The Tv1 culture filtrate significantly reduced the mycelial biomass of the pathogen by recording 0.40 mg biomass production. It was followed by Tv3 (1.10 mg) and Tv5 (1.41 mg). All the isolates of Trichoderma inhibited the formation of sclerotia over control which recorded 121 sclerotia after 14 days of inoculation (Table 8).
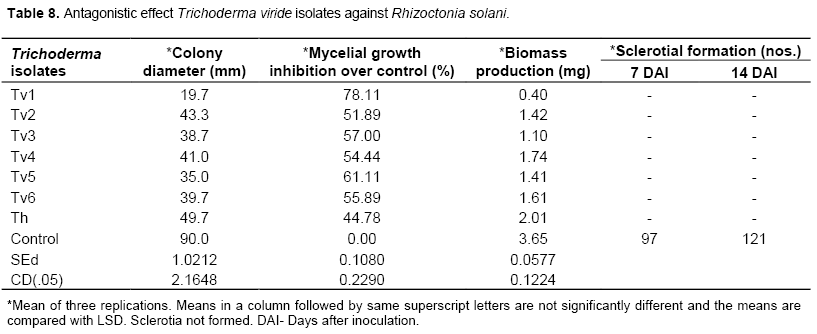
Similarly, Khan and Sinha (2007) reported that, T. harzianum (rice leaf sheath isolate) was found most effective against R. solani in in vitro and glasshouse conditions. T. harzianum is well known biocontrol agents against several soil borne pathogens. Osman et al. (2011) showed that, T. harzianum was found effective in inhibiting the R. solani causing soya beans root rot. In the present study, T. viride was found to be effective among the other antagonists. Among the seven isolates of Trichoderma spp tested, maximum percentage of inhibition was recorded with the isolate Tv1. All the seven isolates inhibited the growth, mycelial biomass and sclerotial production of R. solani in dual culture. Alamri et al. (2012) proved that the mechanism of T. harzianum JF419706 to suppress the pathogenic fungi viz., Alternaria alternate, Fusarium oxysporum, Exserohilum rostratum, Macrophomina phaseolina, Pythium ultimum, and R. solani was though competition for nutrients, mycoparasitizing and lysis of pathogen’s cell walls. Harman et al. (2012) mentioned that Trichoderma spp. parasitize a range of phytopathogenic fungi.
In vitro effect of botanicals against R. solani
Madhuca longifolia (mahua) seed extract showed maximum inhibition of 34.81% at 20% concentration and it was found significantly superior to other extracts (Plate 1). This was followed by A. indica leaf extract which showed an inhibition of 20% at 20% concentration and O. tenuiflorum (thulasi) leaf extract, 18.15%. All the nine aqueous plant extracts were ineffective at 10% concen-tration (Table 9). The botanicals reduced the mycelial biomass of the pathogen over control. Extract from the M. longifolia seed significantly reduced the mycelial biomass (2.18 mg) over other treatments. It was followed by O. tenuiflorum (2.73 mg) and A. indica (2.95 mg) and they were on par with each other. The leaf extracts of A. indica, L. littorea and M. longifolia seed extract completely inhibited the sclerotial formation at all the three concentrations tested viz., 10, 15 and 20% (Table 8).
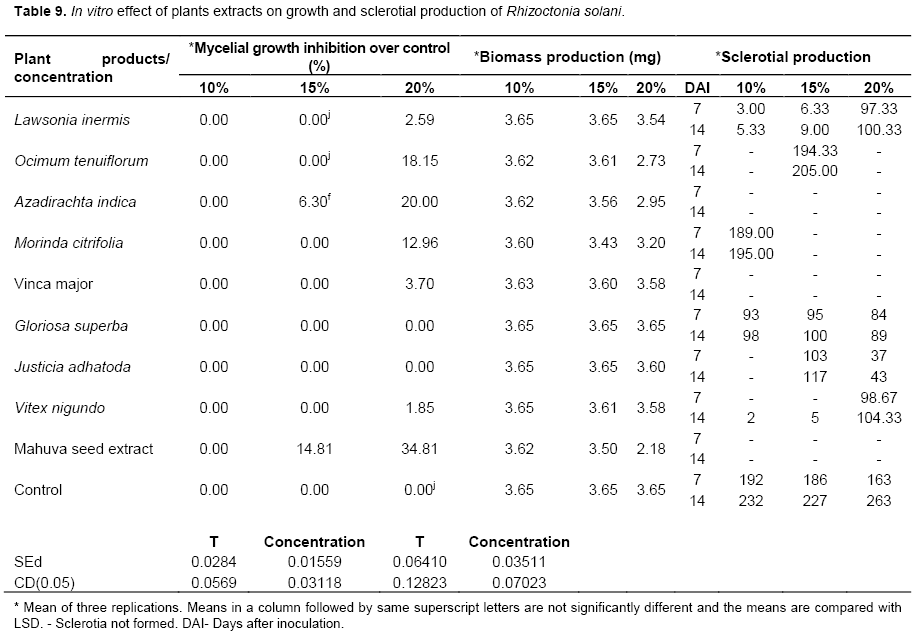
Plant extract are not only easy to prepare but also non-polluting and low priced as compared to commercial fungicides. This is supported by the work of Alabi and Olorunju (2004). In their studies, plants sprayed with neem seed extract gave yields higher than the plants sprayed with black soap and cow dung extract (Alabi and Olorunju, 2004). Gujar et al. (2012) reported that A. indica and A. vera showed inhibition of mycelial growth of the pathogen and can be utilized for the management of fungal diseases caused by the Aspergillus niger, Aspergillus flavus, R. solani, Rhizoctonia bataticola. In this present study, M. longifolia seed extract showed maximum inhibition and it was significantly superior to other plant extracts. All these plant extracts inhibited the mycelial weight and sclerotial formation completely.
All the oil cake extracts tested were not inhibitory at the lowest concentration of 10%. Mahua oil cake was the only extract which was inhibitory at 15% concentration recording 14.81% mycelial growth inhibition over the control. Among the seven extracts, mahua oil cake extract exhibited the maximum mycelial growth inhibition of 49.63% against the pathogen R. solani at 20% concentration (Plate 2). It was followed by neem oil cake (14.07%) and castor oil cake (8.15 %) (Table 11). Mahua oil cake and neem oil cake extract completely inhibited the formation of sclerotia at all concentrations. Among the treatments, maximum sclerotial formation was seen in the Petri plate poisoned with gingelly oil cake extract (117 numbers) at 20% concentration after 14 days of inoculation. The control recorded 199 numbers of sclerotia. It was interesting to note that the lower concentration of the seven oil cake extracts viz., 10 and 15% had inhibited sclerotial formation in solid medium (Table 10). Alice et al. (1998) reported that the presence of antifungal principles present in the mahua cake extract (10%) was effective in combating the jasmine wilt incidence caused by Sclerotium rolfsii.
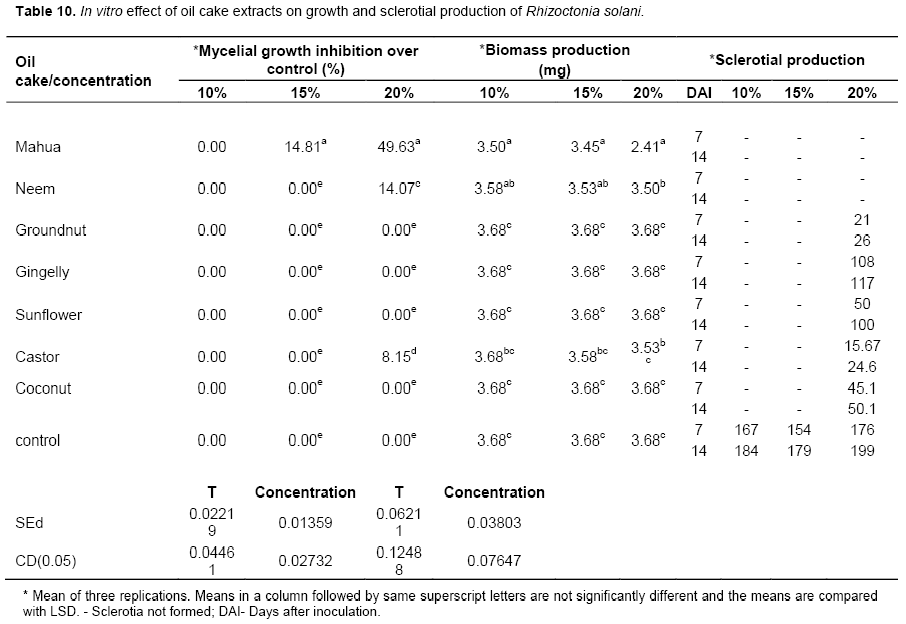
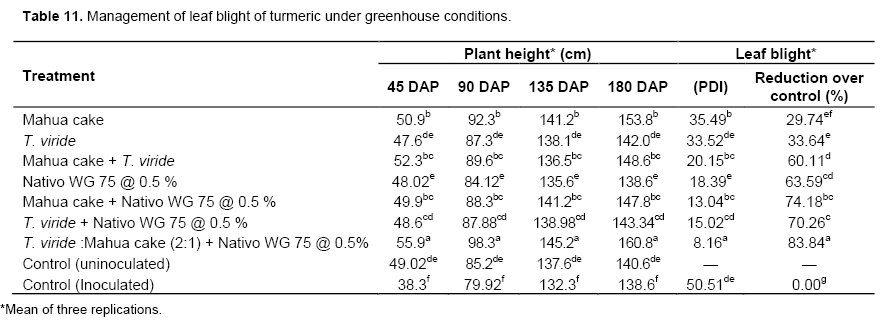
Management of leaf blight of turmeric under greenhouse studies
The effective treatments were culled out from the present research findings and used to perform the pot culture experiments to find out its efficacy against R. solani causing leaf blight of turmeric. Among the seven treatments, the basal soil application of mahua cake and T. viride in the ratio of 1:2 (1 g of mahua cake mixed with 2 g T. viride per kg of pot soil) followed by foliar spray of Nativo @ 0.5% significantly reduced the percent disease incidence over other treatments. This effective treatment recorded 8.6 PDI and a plant height of 160.8 cm on 180 DAP as against the inoculated control which recorded 50.51 PDI with a plant height of 138.6 cm. The disease reduction over control was 83.84% and the increase in plant height accounted for 11.00% (Table 11). The reduction in the turmeric sheath blight incidence and increased plant height in the treated plants in the present study is due to the biocontrol nature of all the treatments imposed.
Similarly, application of T. harzianum amended with organic fertilizer was more efficient alone in managing dam-ping off disease (R. solani) in cucumber (Huang et al., 2011). Clove extract at a concentration of 4% as well as the chemical fungicide (Rizolex-T) significantly reduced the incidence of R. solani in pea (Al-Askar and Rashad, 2010). Addition of mustard oil cake in French bean (Phaseolus vulgaris) along with the inoculation of arbuscular mycorrhizal fungi (AMF) and P. fluorescens was found highly effective in reducing the root rot caused by R. solani (Neeraj and Kanchan, 2011).
CONCLUSION
Research presented here has expanded knowledge of the association of R. solani with turmeric crops in Tamil Nadu, India and about disease symptomology. Fungicide, botanical and bioagents were shown to vary in effectiveness at controlling mycelia growth, sclerotial formation and biomass production of R. solani. Basal application of T. viride + mahua cake and also foliar application of Nativo 0.5% were explored as a means for integrated disease management of leaf blight disease in turmeric crops.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Alabi O, Olorunju EP (2004). Evaluation of neem seed extract, black Soap and cow dung for the control of Groundnut leaf spot at samaru, Nigeria. Arch. Phytopathol. Plant Prot. 37:123-127. |
|
|
Alamri S, Hashem M, Mostafa SY (2012). In-vitro and In-vivo biocontrol of soil borne phytopathogenic fungi by certain bioagents and their possible mode of action. Biocontrol Sci. 17:155-167. |
|
|
Al-Askar AA, Rashad YM (2010). Efficacy of some plant extracts against Rhizoctonia solani on pea. J. Plant. Prot. Res. 50:239-243. |
|
|
Alice D, Ramamoorthy V, Muthenay, Seetharaman K (1998). Biocontrol of jasmine wilt incited by Sclerotium rolfsii Sacc. Indian J. Plant Prot. 26:64-67. |
|
|
Dennis C, Webster J (1971). Antagonist properties of speices group of Trichoderma I. Production of non-volatile antibiotics. Trans. Br. Mycol. Soc. 57:25-39. |
|
|
Dixit D, Srivatava NK, Kumar R, Sharma S (2002). Cultivar variation in yield, metabolite translocation and partitioning of 14 C02 assimilated photosynthate into essential oil and curcumin of turmeric (Curcuma longa L). J. Plant Biol. 29:65-70. |
|
|
Gescher A, Sharma R, Steward W (2005). Curcumin: the story so far. Europ. J. Cancer 41(13):1955-1968. |
|
|
Gurjar MJ, Shahid Ali, Masood A, Kangabam SS (2012). Efficacy of plant extracts in plant disease management. Agric. Sci. 3:425-433. |
|
|
Harman GE, Alfredo H, Herrera E, Benjamin A, Horwitz,Matteo L (2012). Trichoderma – from Basic Biology to Biotechnology. J. Med. Microbiol. 158:1-2. |
|
|
Huang X, Chen L, Ran W, Shen Q, Yang X (2011).Trichoderma harzianum strain SQR-T37 and its bio-organic fertilizer could control Rhizoctonia solani damping-offdisease in cucumber seedlings mainly by the mycoparasitism. Appl. Microbiol. Biotechnol. 91:741-755. |
|
|
Hudge BV, Ghugul SA (2010). Losses in yield and quality of turmeric due to leaf spot disease caused by Colletotricum capsici. Int. J. Agric. Sci. 6:43-45. |
|
|
Hunjan MS, Lore JS, Pannu PPS, Thind TS (2011). Performance of some new fungicides against sheath blight and brown spot of rice. Plant Dis. Res. 26:61-67. |
|
|
Kaushik N, Kumar S, Proksch P (2010). Antifungal activity of extracts of endophytic fungi Chaetomium globosum, isolated from Withania somnifera. 12th IUPAC International Congress of Pesticide Chemistry at University of Melbourne, Australia. |
|
|
Khan AL, Sinha AP (2007). Screening of Trichoderma spp. against Rhizoctonia solanithe causal agent of rice sheath blight. Indian Phytopathol. 60:450-456. |
|
|
Khare A, Singh BK, Upadhyay RS (2010). Biological control of Pythium aphanidermatum causing damping–off of mustard by mutants of Trichoderma viridae1433. J. Agric. Technol. 6:231-243. |
|
|
Lahlali R, Hijri, M (2010) Screening, identi¢cationand evaluation of potential biocontrol fungal endophytes against Rhizoctonia solani AG3 on potato plants. FEMS. Microbiol. Lett. 311:152-159. |
|
|
Neeraj, Kanchan S (2011). Organic amendments to soil inoculated arbuscular mycorrhizal fungi and Pseudomonas fluorescens treatments reduce the development of root-rotdisease and enhance the yield of Phaseolus vulgaris L. Eur. J. Soil Biol. 47:288-295. |
|
|
Neslihan Dikbas, Recep Kotan, Fatih Dadasoglu, Fikrettin Sahin (2008). Control of Aspergillus flavus with essential oil and methanol extract of Satureja hortensis. Int. J. Food Microbiol. 124:179-182. |
|
|
Osman HME, Mostafa M, El Sheekh, Metwally A, Metwally, Abd ElA, Ismail, Mona M (2011). Antagonistic activity of some fungi and Cyanobacteria species against Rhizoctonia solani. Indian J. Plant Pathol. 2:101-114. |
|
|
Padaria JC, Kapoor V (2011). Plasmid borne gene of Bacillus pumillis MTCC 7615 responsible for fungal antagonism towards Rhizoctonia solani. Indian J. Biotechnol. 10:316-320. |
|
|
Roy AK (1992). Severity of Rhizoctonia solani on the leaves of rice and turmeric. Indian Phytopathol. 45:344-347. |
|
|
Seema M, Devaki NS (2012). In-vitro evaluation of biological control agents against Rhizoctonia solani. J. Agric. Technol. 8: 233-240. |
|
|
Sharma NR, Teng PS, Olivares FM (1990). Comparison of assessment methods for rice sheath blight disease. Philipp. Phytopathol. 26:20-24. |
|
|
Shekhawat PS, Prasad E (1971). Antifungal properties of some plant extracts: Inhibition of spore germination. Indian Phytopathol. 24:800-802. |
|
|
Srinivas P, Ved Ratan P, Narayan Reddy, Bindu Madhavi G (2014). In-vitro evaluation of fungicides, biocontrol agents and plant extractsagainst rice sheath blight pathogen rhizoctonia solani. Inter. J. Appl. Biol. Pharma. Technol. 5:121-126 |
|
|
Sriram S, Raguchander T, Vidhyasekaran P, Muthukrishnan S, Samiyappan R (1997). Genetic relatedness with special reference to virulence among the isolates of Rhizoctonia solani causing sheath blight in rice. J. Plant Dis. Prot. 104:260-271. |
|
|
Swamy HN, Sannaulla S, Kumar MD (2009). Screening of new fungicides against rice sheath blight disease. Karnataka J. Agric. Sci. 22:448-449. |
|
|
Taheri P, Gnanamanickam S, Hofte M (2007). Characterization, genetic structure, and pathogenicity of Rhizoctonia spp. associated with rice sheath diseases in India. Phytopathology 97:373-383. |
|
|
Wang X, Luo C, Chen Z (2012). Genome Sequence of the Plant Growth-Promoting RhizobacteriumBacillus sp. Strain 916. J. Bacteriol. 194(19):54-67. |
|
|
Zachow C, Jamshid F, Massimiliano C, Ralf T, Gabriele B (2010). Strain-specific colonization pattern of Rhizoctonia antagonists in the root system of sugar beet. FEMS. Microbiol. Ecol. 7:124-135. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


