Full Length Research Paper

ABSTRACT
The rising antibiotic resistance is urging researchers to explore for new forms of antibiotics, notably from soil microorganisms such as Bacillus species. This study aimed to screen Bacillus strains from soil samples in Sudan for antifungal activity and to review relevant compounds from members of the genus in the literature. Out of 10 isolates from soil in Sudan, the strain JS6 (DSM 28831) was found active against representative zygomycete fungi and consequently subjected to thorough identification and chemical analyses of secondary metabolites. Confirmation of the initially identified Bacillus spp. was done using 16S rDNA gene sequence analysis which indicated a novel species (accession MF099872) that is closely related to Bacillus siamensis, Bacillus amyloliquefaciens, and Bacillus nakamurai. The crude culture-free filtrate and both chloroform and ethyl acetate extracts authenticated the initial antifungal activity of this strain, which exceeded that of amphotericin B, a standard antifungal agent. GC-MS results of the extracts revealed 32 compounds which included long-chain fatty acids, fatty acid methyl esters, alkaloids, and alcoholic compounds. Seven biologically active compounds were identified from Bacillus spp. strain JS6 and are equally found in the literature originating from plant or microbial sources. In the literature, these compounds show various activities such as antifungal, antioxidant, hypocholesterolemic, nematicide, pesticide, antiandrogenic, flavour, haemolytic, alpha reductase inhibitor, and other antimicrobial activities. The analysis identified a major unique antifungal peak (rt, 23.142; area, 25.36%) as 5,10-Diethoxy-2,3,7,8-tetrahydro-1H,6H-dipyrrolo[1,2-a:1',2'-d]pyrazine, a heterocyclic aromatic organic compound, that has been previously detected in Lactobacillus casei.
Key words: Bacillus species, chromatography, TLC, GC-MS, soil, aromatic organic compounds.
INTRODUCTION
Members of the genus Bacillus are Gram-positive aerobic bacteria that are widely distributed in nature, notably soil. They are known to produce large amounts of secondary metabolites with different biological and biotechnological activities including antimicrobials (Boottanun et al., 2017). Since the late 19th Century, Bacillus species have been recognized as a significant source of antibiotics (Schaffer, 1969).
Bacillus spp. express a range of secondary metabolite gene clusters encoding polyketide synthases and non-ribosomal peptide synthetases responsible for a notable number of polyketides and lipopeptides. Such compounds have been, in part, exploited for medical and agricultural purposes (Aleti et al., 2015). These metabolites have been a subject for wide arrays of scientific studies including chemical, biological activities, and industrial pharmaceuticals (Aleti et al., 2015; Mondol et al., 2013; Sansinenea and Ortiz, 2011). Screening of extracts from Bacillus spp. uncovered a considerable structural variety of many natural compounds with broad biological activities. These activities include antimicrobial, antiviral, immunosuppressive, antitumor activities, and antioxidants (Youcef-Ali et al., 2014).
Microbial sources have been shown to be powerful natural products of many kinds (Zeliha, 2017). Several studies have focused on plant compounds (Erkan et al., 2008). However, only a few reports have been conducted on the antioxidant power of microbial extracts (Moktan et al., 2008). Production of such compounds allows the bacterium to survive in its natural ecosystem (Sansinenea and Ortiz, 2011). Antifungal peptides produced by Bacillus spp. include mycobacillins (Majumdar and Bose, 1958), surfactins (Kluge et al., 1988), mycosubtilins (Peypoux et al., 1976), and fungistatins (Islam et al., 2012). Bacillus spp. can produce a wide range of other metabolites, including chitinases and other cell wall-degrading enzymes (Frandberg and Schnurer, 1994), volatiles (Sadfi et al., 2001), and compounds that elicit plant resistance mechanisms (Islam et al., 2012). Volatile metabolites produced from Bacillus spp. have been reported to inhibit mycelia growth of Fusarium oxysporum, with the highest effect on the reduction of Fusarium wilt of onion (Sharifi Tehrani and Ramezani, 2003). Ryu et al. (2003)reported on the promotion of growth and induction of systemic resistance (ISR) response in Arabidopsis thaliana against Erwinia carotovora subsp. carotovora by volatile substances (VS) (acetylbuanediol and acetoin, same as the present purified compounds) from Bacillus amyloliquefaciens isolate IN937a and Bacillus subtilis isolate GB03. Therefore, VS-producing bacteria can be used as biocontrol agents for protection against microbial plant diseases (Islam et al., 2012). Genomic studies have revealed that the genome of B. amyloliquefaciens holds many gene clusters involved in the synthesis of antifungal and antibacterial acting secondary metabolites. Five gene clusters, srf, bmy, fen, nrs, dhb, covering altogether 137 kb, direct non-ribosomal synthesis of the cyclic lipopeptides surfactin, bacillomycin, fengycin, an unknown peptide, and the iron siderophore bacillibactin have been identified (Chen et al., 2009).
The present study was carried out to screen potential bioactive compounds from a Bacillus strain JS6, to review the literature and compare these with relevant ones in the literature.
MATERIALS AND METHODS
Isolation of bacilli from soil and their identification
Soil samples collected from different locations in Sudan were spread onto Tryptic soya agar (TSA; Oxoid Inc., UK) and incubated at 37°C for one week. Ten colonies were isolated from the soil samples and were identified to be members of the genus Bacillus. The isolates were at first identified using phenotypic criteria: microscopy, growth properties, physiological and biochemical tests (Turnbull, 1999), and subsequently considered for antifungal screening.
Confirmation of the initially identified Bacillus spp. was done using 16S rDNA gene sequence analysis. The sequencing was done by Macrogen Inc. (Seoul 08511, Korea). The acquired DNA sequences (1495 bp) were initially inspected and adjusted by Chromas (version 2.6.6 (2018), Technelysium Pty Ltd., South Brisbane, Queensland, Australia). The sequence was then aligned alongside all obtainable 16S rRNA gene data of valid species by using the BLAST: Basic Local Alignment Search Tool (nih.gov) (Zhang et al., 2000)and the EZbioCloud 16S database (www.ezbiocloud.net) to show the phylogenetic relationship. The final phylogenetic tree was constructed by the MEGA7 program (Kumar et al., 2016), using the neighbor-joining method with bootstrap values based on 100 replications.
Antibiotic production
Isolated bacilli were screened for potential antibiotic production. Each bacillus was cultured as one line and fungi were streaked on a perpendicular line on TSA plates and incubated at 37°C. The tested fungi were Basidiobolus zygomycetes, namely: Basidiobolus haptosporus-like strain 49-4, strain V81 (DSM06014), and strain Doza.
Incubated plates were checked daily for zones of inhibition. One isolate, Bacillus spp. strain JS6, was found potentially active and selected for further analysis.
Potential bioactive Bacillus spp. strain JS6, which showed zones of inhibition, was purified, subcultured in TSA broth (250-500 mL), incubated at 37°C, and checked for purity. A few drops from the broth containing bacteria and envisaged antimicrobials were placed on a fresh TSA plate containing bacteria and fungi to see and confirm the presence of inhibition again. The crude antibiotic solution which contained live bacilli and media ingredients in the 250 to 500 mL flask was subjected to separation via filtration. The pure filter solution was ready for chemical work.
Paper disk assay
Paper disk assay was performed as per CLSI Guidelines (CLSI, 2018). 500 μL of inoculums was spread on the surface of nutrient agar medium. Sterile discs (6 mm diameter) made of Whatman paper No. 1 were dipped into the test extract and were put onto the agar surface after complete drying. Commercially available discs were served as positive control. Plates were then incubated at 30°C for 24 h. After incubation, plates were observed for zones of inhibition.
Antibiotic extraction and thin layer chromatography
The culture broth (500 L) was centrifuged at 5,000 rpm for 10 min to separate the supernatant and mycelial biomass. Mycelium and culture supernatants were extracted with ethyl acetate and chloroform, respectively. The extracts were dried and resuspended in a small volume of methanol and kept for the next separation steps. To visualize the number of compounds present in the extracts, thin layer chromatography (TLC) was performed. Aluminium plates precoated with silica gel and two mobile phases [ethyl acetate: methanol (1:1) and Petroleum ether: chloroform (1:1)]. Chromatograms were visualized by ninhydrin reagent and then observed under UV light.
Gas chromatography mass spectrometer (GC-MS)
The antifungal compounds were identified by using GC-MS technique (GC-MS, Shimadzu QP2010 Ultra, Japan) following standard method (Adams, 2005). GC-MS was equipped with a capillary column (30 m × 0.25 mm i.d.; 0.25 mm) and a HP 5975B mass selective detector and 70 eV energy was used for electron ionization detection at 50°C for 3 min, then steadily increased to 250°C at 3°C/min rate and held at this temperature for 4 min. Injector and MS transfer line temperature was held at 220 and 250°C and helium was used as carrier gas (1 mL/min glow rate). The compounds were identified on the basis of comparison of the retention time and mass spectra with those in the NIST98 GC-MS library.
RESULTS
Isolation and identification of bacilli from soil
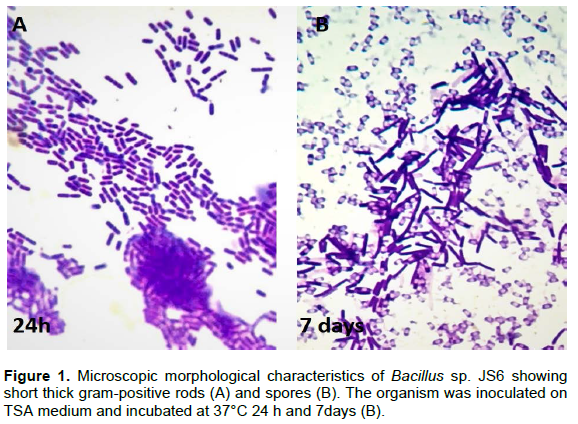
Ten isolates were initially identified as belonging to the genus Bacillus. The strain JS6, which was found active against tested fungal species, was subjected to detailed identification and stored for future research. A few morphological characteristics were found sufficient to designate the isolate as a Bacillus sp. (Figure 1). The strain JS6 showed typical bacillus properties, namely, short thick Gram-positive rods, which was demonstrated in a 5-day-old aerobic culture on TSA medium at 37°C. The strain was deposited in the Leibniz Institute DSMZ-German Collection of Microorganisms and Cell Cultures GmbH, Inhoffenstraße 7B 38124 Braunschweig, Germany (DSM 28831) for future description or use.
16S rDNA sequence analysis
16S rDNA nucleotide sequence data (1495 bp; Genbank accession no. MF099872) from Bacillus spp. JS6 was run on BLAST system with nearly all Bacillus spp. found in Genbank database. The sequence was further analyzed by MEGA6 software (Tamura et al., 2013), which assigned it to Bacillus sp. (Figure 2). All positions containing gaps and missing data were eliminated from the final analysis.
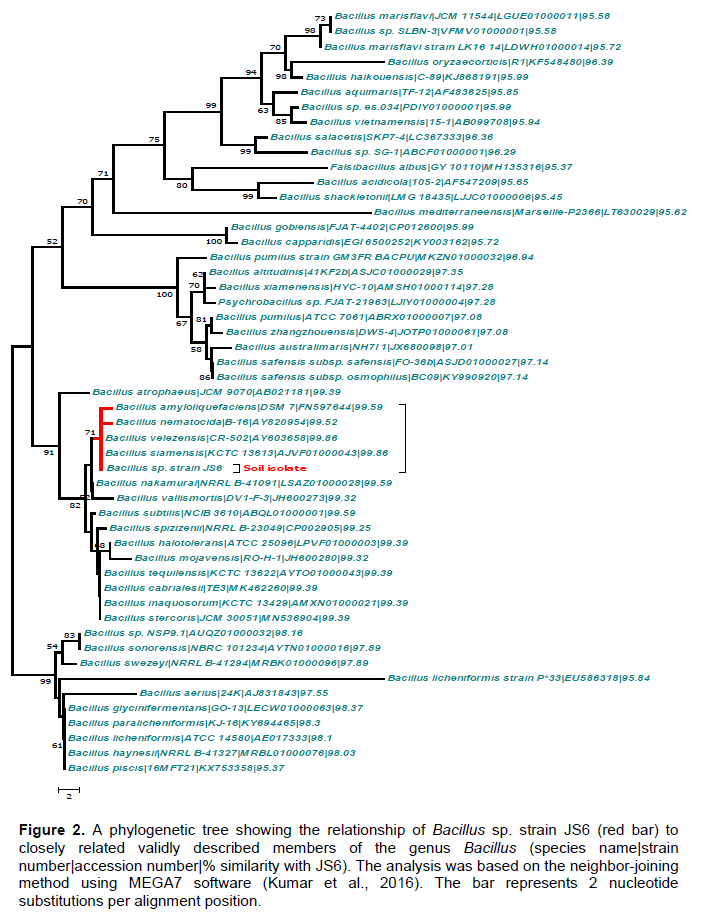
The strain was found to have a closer resemblance to Bacillus siamensis KCTC 13613, 99.86%; B. amyloliquefaciens strain DSM 7, 99.59%; Bacillus nakamurai NRRL B-41091, 99.59%; B. subtilis NCIB 3610, 99.59%; and Bacillus nematocida B-16, 99.52%.
Thin layer chromatography
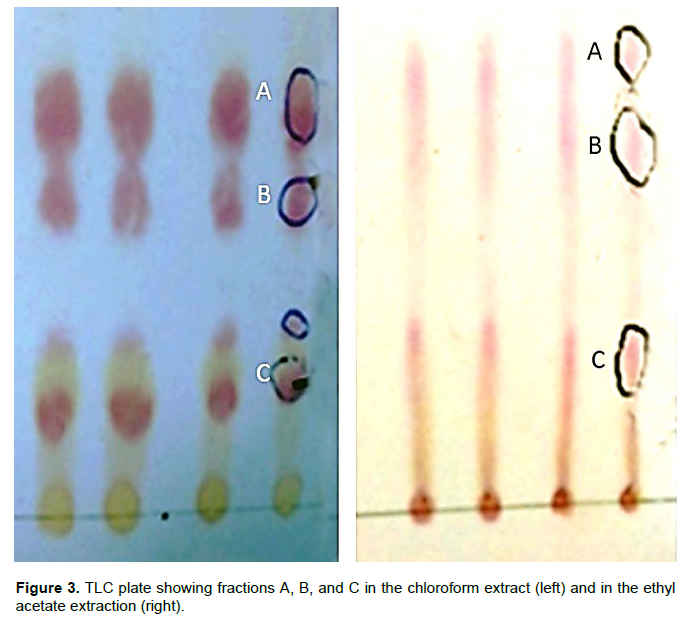
The present study adopts solvent extraction, thin layer chromatography (TLC) separation, and GC-MS chromatography to purify and identify the active antifungal compounds produced by strain JS6. Ethyl acetate and chloroform extraction was performed on purified cell-free culture solution. The TLC plate of the chloroform extract showed a better result than ethyl acetate extract, but in both extract fractions A, B and C were distinct (Figure 3). Some tailings can also be seen, and hence more solvent systems with polarity adjusting should be used for further studies.
Inhibitory activity of the chloroform and ethyl acetate extracts
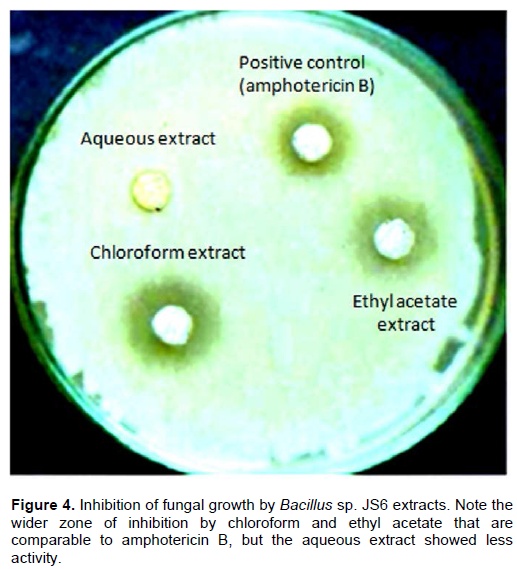
Fractions A, B, and C of the chloroform extract and ethyl acetate extraction were found to have inhibitory activity against many molds. Figure 4 shows that the wider zones of inhibition by chloroform and ethyl acetate have been compared to amphotericin B and the aqueous extract. No significant e=inhibition was revealed by the aqueous extract. The activity that is comparable to amphotericin B but the aqueous extract showed less activity.
GC-MS chromatogram
The GC-MS chromatogram of strain JS6 in general showed narrow, resolved peaks; a noisy baseline could indicate the existence of a trace amount of compounds in the extracts. The GC-MS analysis identified a total of 32 compounds from the extract of both ethyl acetate (Figure 5) and chloroform (Figure 6). The extract of ethyl acetate revealed 19 compounds (Table 1), and 13 compounds from the chloroform extract (Table 2), but any of these compounds could be responsible for the antifungal activity that detected in this study. Interestingly, these compounds found in the chloroform extract differ from the compounds found in the ethyl acetate extract due to the solvent’s polarity difference. The antifungal activity of both extracts is significant due to the presence of a variety of interesting compounds.
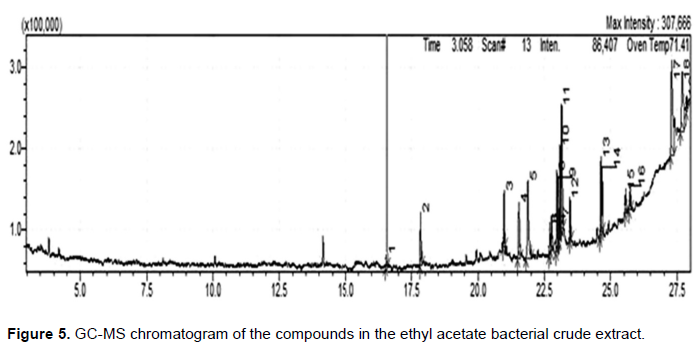
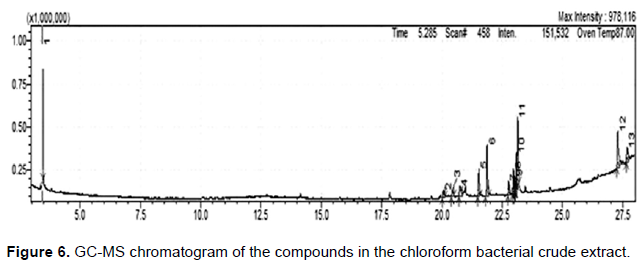

DISCUSSION
The discovery of novel medicinal drugs is a top task of scientific, economic, and biomedical importance. Traditionally, the bulk of the new drugs have been generated from natural products (secondary metabolites) and compounds originated from natural products are likely sources of fascinating bioactive secondary metabolites (Zhang et al., 2020). During the previous two decades, pharmaceutical research in the field of natural products has dropped, partly because of an emphasis on high-throughput screening of synthetic libraries. At present, there is a considerable drop in new drug approvals and this could lead to an imminent loss of patent protection for important medicines (Li and Vederas, 2009; Zhang et al., 2020).
In this study, one Bacillus strain out of 10, which was isolated from soil in Sudan, was characterized and found to have antifungal activity. This first objective, which dealt with identifying the strain and its bioactive compounds, was further expanded by comparing and listing compounds from all known species of the genus Bacillus. Our investigation revealed 32 compounds comprising long-chain fatty acids, fatty acid methyl esters, alkaloids, and alcoholic compounds. Seven of these were found to have biological activities. Besides, the major distinctive antifungal 5,10-Diethoxy-2,3,7,8-tetrahydro-1H,6H-dipyrrolo[1,2-a:1',2'-d]pyrazine.
Activity of 5,10-Diethoxy-2,3,7,8-tetrahydro-1H,6H-dipyrrolo[1,2-a:1',2'-d]pyrazine
The strain JS9 of this study produced the unique compound 5,10-diethoxy-2,3,7,8-tetrahydro-1H,6H-dipyrrolo[1,2-a:1',2'-d]pyrazine. This compound showed antifungal activity against zygomycete fungi. This compound is documented in the Springer Nature References (PubCheM 2004). It has been first detected in Lactobacillus casei (Li et al., 2012a)and found to have antifungal activity. This result supports the current findings both in chemical structure and antimicrobial properties. To our knowledge, no other information are available regarding this unique compound.
A chain of 2-pyrazolines 5-9 has been manufactured from α,β-unsaturated ketones 2-4. Some of the newly identified pyrazoline and pyrazole derivatives were found to have antibacterial and antifungal activity (Hassan, 2013). Several compounds including 4-(2,5-dimethylpyrrol-1-yl)/4-pyrrol-1-yl benzoic acid hydrazide analogs have been synthesized and were found to have in vitro antibacterial, antifungal, and antitubercular activity at non-cytotoxic concentrations (Joshi et al., 2013).
Biologically active compounds identified from Bacillus sp. strain JS6 and are equally detected from other sources
In this search, seven major compounds have been detected in the strain JS6. These compounds have already been detected from different sources, and all have biological activities. Names of these compounds, their natures, activities, sources, and references are shown in Table 3. The structures of these compounds are as shown in Figure 7.
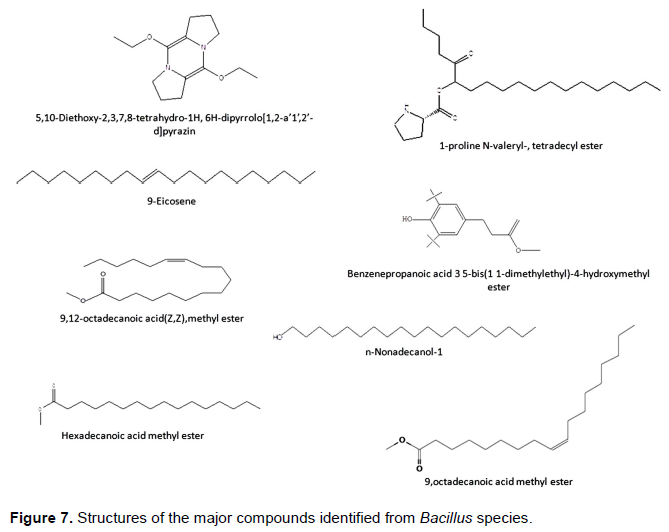
Beevi et al. (2014)have investigated the antibacterial activity and chemical characterization of metabolites of B. subtilis isolated from Sea Surface Microlayer. GC-MS analysis revealed the presence of 9-Ecosene with 15.0% with a potential antibacterial activity (Akpuaka et al., 2013). Hema et al. (2011)carried out the GC/MS significant producers of diverse groups of peptides determination of bioactive components of Murraya koenigii, the curry tree. The curry tree, sometimes called sweet neem, is a tropical tree (Rutaceae) which is native in most Asian countries. Their results revealed the presence of 9-octadecanoic acid, methyl ester (E)- in this plant. Additionally, Asghar et al. (2011)have done GC-MS analysis of petroleum ether extract (oil) and bioassay of crude extract of Iris germanica, which is a species of flowering plant in the family Iridaceae commonly known as bearded iris or the German bearded iris. The petroleum ether extract (oil) of this plant has resulted in the identification of eleven compounds 9-octadecanoic acid methyl ester (E)- was identified with an anti-carcinogenic activity (Yeong et al., 1989).
Li et al. (2012b)distinguished 5,10-Diethoxy-2,3,7,8-tetrahydro-1H,6H-dipyrrolo[1,2-a:1',2'-d]pyrazine as an antifungal in the identification of antifungal compounds produced by Lactobacillus AST18. The antioxidant activity, total phenolics, and GC-MS study of Vitex negundo was investigated (Kumar et al., 2010). The study revealed the activity of 9, 12-octadecanoic acid (Z,Z), methyl ester (E)- and hexadecanoic acid methyl ester shown in Table 3. Bashir et al. (2012)studied the chemical composition and antifungal, phytotoxic, brine shrimp cytotoxicity, insecticidal, and antibacterial activities of the essential oils of Acacia modesta, benzenepropanoic acid 3 5-bis (1 1-dimethylethyl)-4-hydroxy- methyl ester was identified with antifungal and antioxidant activities. N-nonadecanol-1 and the steroid β-sitosterol were isolated for the first time from Cryptantha incana (Alemu et al., 2015). The results proved that the polar extracts of the seeds and pods of C. incana possess antibacterial activity.
Compounds identified from Bacillus spp. with antifungal activity
Members of the Bacillaceae family, including Bacillus, Brevibacillus, Paenibacillus, Aneurinibacillus, and Halobacillus species are with antibacterial, antifungal, and antiviral activities (Zhao et al., 2018). A range of bacteria produce lipopeptides, but Bacillus and Paenibacillus spp. produced several potent antimicrobial lipopeptides. These lipopeptides have been renowned for years as a prospective source of antibiotics such as polymyxins, octapeptins, polypeptins, iturins, surfactins, fengycins, fusaricidins, tridecaptins, and kurstakins (Cochrane and Vederas, 2016). Iturins, which is a special class of pore-forming lipopeptides, have been extracted from culture media of B. subtilis and was found to have strong antifungal activity against many yeasts and fungi (Maget-Dana and Peypoux, 1994).
The antifungal antibiotics fusaricidins A, B, C, and D, which are yielded by Bacillus polymyxa KT-8, were found more effective than bacillopeptins in their antimicrobial activity (Kaneda and Kajimura, 2002).
Bacillus licheniformis 09IDYM23 produces Ieodoglucomide C glycolipids which are considered good candidates for the development of new fungicides (Tareq et al., 2015). Bacillus spp., among other microbes associated with sponges, were recognized as noticeable producers of antimicrobial compounds (Indraningrat et al., 2016). Bacillus brevis produces an antimicrobial peptide named tyrothricin. Tyrothricin showed activity against bacteria, fungi, and some viruses (Lang and Staiger, 2016). Lipopeptides of Bacillus spp. were found to have a range of biological activities, including interactions with biofilms and anti-fungal, anti-inflammatory, anti-tumor, anti-virus, and antiplatelet properties (Zhao et al., 2017). Mass spectrometry and HPTLC bioautography analysis of purified compounds from Bacillus isolates indicated the presence of lipopeptides, thus confirming their biocontrol function (Fira et al., 2018).
CONCLUSION
The present study was an attempt to search for antifungal compounds from an alternative natural source. Seven biologically active compounds were identified from our Bacillus sp. strain JS6 and are equally detected from other sources recovered from the literature. These compounds exhibit various activities such as antifungal, antioxidant, hypocholesterolemic, nematicide, pesticide, antiandrogenic, flavour, haemolytic, alpha reductase inhibitor, and other antimicrobial activities.
The present study revealed the presence of a distinctive antifungal compound, a pyrrole-pyrazine derivative, peak: rt, 23.142 with an area 25.36%, as 5,10-diethoxy-2,3,7,8-tetrahydro-1H,6H-dipyrrolo[1,2-a:1',2'-d]pyrazine. This is a heterocyclic aromatic organic compound, which has been detected in L. casei before. Optimizing culture conditions may influence the production of secondary metabolites from Bacillus spp.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
The authors would like to thank Ms. Reem and all staff of the Phytochemistry Laboratory in the Faculty of Pharmacy, University of Medical Sciences and technology. They are grateful to Mr. Babiker Siddig and Dr Alawia Imam’s of the Pharmaceutical Development Center and Mr. Adil Mahjoub in Preventive Medicine and Public Health, Faculty of Veterinary Medicine, University of Khartoum for laboratory assistance.
REFERENCES
|
Adams RP (2005). Identification of essential oil components by gas chromatography/quadrupole mass spectroscopy Journal of the American Society for Mass Spectrometry 16:1902-1903. |
|
|
Akpuaka A, Ekwenchi MM, Dashak DA, Dildar A (2013). Biological activities of characterized isolates of n-hexane extract of Azadirachta indica A. Juss (Neem) leaves. Nature and Science 11(5):141-147. |
|
|
Alemu MA, Mekannen HG, Annisa ME (2015). Phytochemical analysis and antibacterial activity on seed and pod extracts of Crotalaria incana. Journal of Pharmacy and Pharmacognosy Research 3(4):100-108. |
|
|
Aleti G, Sessitsch A, Brader G (2015). Genome mining: Prediction of lipopeptides and polyketides from Bacillus and related Firmicutes. Computational and structural biotechnology journal 13:192-203. |
|
|
Asghar SF, Rehman HU, Choudahry MI, Rahman AU (2011). Gas Chromatography-Mass Spectrometry (GC-MS) analysis of petroleum ether extract (oil) and bioassays of crude extract of Iris germanica. International Journal of Genetics and Molecular Biology 3(7):95-100. |
|
|
Bashir A, Ibrar K, Shumaila B, Sadiq A (2012). Chemical composition and antifungal, phytotoxic, brine shrimp cytotoxicity, insecticidal, and antibacterial activities of the essential oils of Acacia modesta. Journal of Medicinal Plants Research 6(31):4653-4659. |
|
|
Beevi AH, Maruthupandy M, JeevaPriya R, Anand M (2014). Characterization of metabolites of bacillus subtilis isolated from sea surface microlayer and its antibacterial activity. International Journal of Pharmaceutical Science and Health Care 2(4):15-24. |
|
|
Boottanun P, Potisap C, Hurdle JG, Sermswan RW (2017). Secondary metabolites from Bacillus amyloliquefaciens isolated from soil can kill Burkholderia pseudomallei. AMB Express 7(1):16. |
|
|
Chen XH, Koumoutsi A, Scholz R, Borriss R (2009). More than anticipated - production of antibiotics and other secondary metabolites by Bacillus amyloliquefaciens FZB42. Journal of molecular microbiology and biotechnology 16(1-2):14-24. |
|
|
Clinical and Laboratory Standards Institute (CLSI) (2018). Performance standards for antimicrobial disk susceptibility tests; approved standard-12th ed. M02-A13. Wayne, PA. |
|
|
Cochrane SA, Vederas JC (2016). Lipopeptides from Bacillus and Paenibacillus spp.: A Gold Mine of Antibiotic Candidates. Medicinal research reviews 36(1):4-31. |
|
|
Erkan N, Ayranci G, Ayranci E (2008). Antioxidant activities of rosemary (Rosmarinus Officinalis L.) extract, blackseed (Nigella sativa L.) essential oil, carnosic acid, rosmarinic acid and sesamol. Food chemistry 110(1):76-82. |
|
|
Fira D, Dimkic I, Beric T, Lozo J, Stankovic S (2018). Biological control of plant pathogens by Bacillus species. Journal of biotechnology 285:44-55. |
|
|
Frandberg E, Schnurer J (1994). Evaluation of a chromogenic chito-oligosaccharide analogue, p-nitrophenyl-beta-D-N,N'-diacetylchitobiose, for the measurement of the chitinolytic activity of bacteria. The Journal of applied bacteriology 76(3):259-63. |
|
|
Hassan SY (2013). Synthesis, antibacterial and antifungal activity of some new pyrazoline and pyrazole derivatives. Molecules 18(3):2683-711. |
|
|
Hema R, Kumaravel S, Alagusundaram K (2011) GC/MS Determination of Bioactivecomponents of Murraya koenigii. Journal of American Science 7(1):80-83. |
|
|
Hsouna AB, Trigui M, Mansour RB, Jarraya RM, Damak M, Jaoua S (2011). Chemical composition, cytotoxicity effect and antimicrobial activity of Ceratonia siliqua essential oil with preservative effects against Listeria inoculated in minced beef meat. The International Journal of Food Microbiology 148(1):66-72. |
|
|
Indraningrat AA, Smidt H, Sipkema D (2016). Bioprospecting Sponge-Associated Microbes for Antimicrobial Compounds. Marine drugs 14(5). |
|
|
Islam MR, Jeong YT, Lee YS, Song CH (2012). Isolation and Identification of Antifungal Compounds from Bacillus subtilis C9 Inhibiting the Growth of Plant Pathogenic Fungi. Mycobiology 40(1):59-66. |
|
|
Joshi SD, More UA, Kulkarni VH (2013). Synthesis, Antimicrobial and cytotoxic activity of New Heterocyclic Hybrids Based on 2,5-Dimethylpyrrole and Pyrrole Scaffolds. Indian journal of pharmaceutical sciences 75(3):310-23. |
|
|
Kaneda M, Kajimura Y (2002). [New antifungal antibiotics, bacillopeptins and fusaricidins]. Yakugaku zasshi : Journal of the Pharmaceutical Society of Japan 122(9):651-71. |
|
|
Kluge B, Vater J, Salnikow J, Eckart K (1988). Studies on the biosynthesis of surfactin, a lipopeptide antibiotic from Bacillus subtilis ATCC 21332. FEBS letters 231(1):107-10. |
|
|
Kumar PP, Kumaravel S, Lalitha C (2010). Screening of antioxidant activity, total phenolics and GC-MS study of Vitex negundo. African Journal of Biochemistry Research 4(7):191-195. |
|
|
Kumar S, Stecher G, Tamura K (2016). MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Molecular biology and evolution 33(7):1870-4. |
|
|
Lang C, Staiger C (2016). Tyrothricin--An underrated agent for the treatment of bacterial skin infections and superficial wounds? Die Pharmazie 71(6):299-305. |
|
|
Li H, Liu L, Zhang S, Cui W, Lv J (2012a). Identification of antifungal compounds produced by Lactobacillus casei AST18. Current microbiology 65(2):156-61. |
|
|
Li H, Liu L, Zhang S, Cui W, Lv J (2012b). Identification of antifungal compounds produced by Lactobacillus casei AST18. Current microbiology 65(2):156-61. |
|
|
Li JW, Vederas JC (2009). Drug discovery and natural products: end of an era or an endless frontier? Science 325(5937):161-5. |
|
|
Maget-Dana R, Peypoux F (1994). Iturins, a special class of pore-forming lipopeptides: biological and physicochemical properties. Toxicology 87(1-3):151-74. |
|
|
Majumdar SK, Bose SK (1958). Mycobacillin, a new antifungal antibiotic produced by B. subtilis. Nature 181(4602):134-5. |
|
|
Moktan B, Saha J, Sarkar PK (2008). Antioxidant activities of soybean as affected by Bacillus-fermentation to kinema. Food Research International 41(6):586-593. |
|
|
Mondol MA, Shin HJ, Islam MT (2013). Diversity of secondary metabolites from marine Bacillus species: chemistry and biological activity. Marine drugs 11(8):2846-72. |
|
|
Peypoux F, Michel G, Delcambe L (1976). [The structure of mycosubtilin, an antibiotic isolated from Bacillus subtilis (author's transl)]. European journal of biochemistry 63(2):391-8. doi:10.1111/j. |
|
|
PubCheM (2004). PubChem Compound Summary National Library of Medicine (US), National Center for Biotechnology Information. [cited 2021 Jan. 23] Available from: https://pubchem.ncbi.nlm.nih.gov/compound/551125, Bethesda (MD) |
|
|
Ryu CM, CM, Farag Ma, Hu CH, Reddy, MS, Wei, H.-X., Paré, PW, Kloepper JW (2003). Bacterial volatiles promote growth in Arabidopsis. Proc Natl Acad Sci U S A 100(8):4927-32. |
|
|
Sadfi N, Chérif M, Fliss I, Boudabbous AHA (2001). Evaluation of bacterial isolates from salty soils and Bacillus thuringiensis strains for the biocontrol of Fusarium dry rot of potato tubers. Journal of Plant Pathology 83:101-118. |
|
|
Sansinenea E, Ortiz A (2011). Secondary metabolites of soil Bacillus spp. Biotechnology letters 33(8):1523-38. |
|
|
Schaffer P (1969). Sporulation and the production of antibiotics, exoenzymes, and exotoxins. Bacteriological Reviews 33(1):155-164. |
|
|
Sharifi Tehrani A, Ramezani M (2003). Biological control of Fusarium oxysporum, the causal agent of onion wilt by antagonistic bacteria. Communications in agricultural and applied biological sciences 68(4 Pt B):543-7. |
|
|
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013). MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Molecular biology and evolution 30(12):2725-9. |
|
|
Tareq FS, Lee HS, Lee YJ, Lee JS, Shin HJ (2015). Ieodoglucomide C and Ieodoglycolipid, New Glycolipids from a Marine-Derived Bacterium Bacillus licheniformis 09IDYM23. Lipids 50(5):513-9. |
|
|
Turnbull PC (1999). Definitive identification of Bacillus anthracis--a review. Journal of applied microbiology 87(2):237-240. |
|
|
Yeong LH, Nancy KG, Micheal WP (1989). Newly recognized nticarcinogenic fatty acids: identification and quantification in natural and processed cheeses. The Journal of Agricultural and Food Chemistry 37(1):75-81. |
|
|
Youcef-Ali M, Chaouche Nk, Dehimat L, Bataiche I, Mounira Ka, Cawoy He, Thonart P (2014). Antifungal activity and bioactive compounds produced by Bacillus mojavensis and Bacillus subtilis. African Journal of Microbiology Research 8(6):476-484. |
|
|
Zeliha S (2017). Polyphenolic compounds in human Health with pharmacological properties. Journal of traditional medicine and clinical naturopathy 6:4. |
|
|
Zhang B, Zhang T, Xu J, Lu J, Qiu P, Wang T, Ding L (2020). Marine Sponge-Associated Fungi as Potential Novel Bioactive Natural Product Sources for Drug Discovery: A Review. Mini reviews in medicinal chemistry 20(19):1966-2010. |
|
|
Zhang Z, Schwartz S, Wagner L, Miller W (2000) A greedy algorithm for aligning DNA sequences. Journal of computational biology : a journal of computational molecular cell biology 7(1-2):203-14. |
|
|
Zhao H, Shao D, Jiang C, Shi J, Li Q, Huang Q, Jin M (2017). Biological activity of lipopeptides from Bacillus. Applied Microbiology and biotechnology 101(15):5951-5960. |
|
|
Zhao P, Xue Y, Gao W, Li J, Zu X, Fu D, Zhang F (2018). Bacillaceae-derived peptide antibiotics since 2000. Peptides 101:10-16. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


