Full Length Research Paper

ABSTRACT
Radio-sensitivity test informs the optimal irradiation dose for use in mutation breeding. In this study, four rice varieties (Basmati 217, Basmati 370, ITA310, and Komboka) were irradiated with gamma-ray doses ranging from 0 to 500 Gy, to examine their morphological responses at the seedling stage, and estimate their optimal doses. Two experiments were carried out at varied time points between March and May 2022. In the first experiment, the irradiated seeds were pre-germinated before sowing in soil premixed with base fertilizer, while in the second experiment, irradiated seeds were imbibed by soaking in water for 24 h followed by direct sowing in soil. Observations were made on germination percentage, rate of seedling emergence, survival, and seedling height. The results indicated that sensitivity to gamma irradiation varied across varieties. Basmati 370 and Komboka were the most sensitive, ITA310 was moderate, while Basmati 217 was the least sensitive. At low doses of 50, 100, and 150 Gy, gamma irradiation enhanced seedling survival and height, but at higher doses above 300 Gy, they were significantly reduced. LD50 values ranged from 354 to 556 Gy, reduction dose 30 (RD30) ranged from 267 to 426 Gy, while reduction dose 50 (RD50) ranged from 335 to 531 Gy.
Key words: Mutation breeding, radio-sensitivity, LD50, RD30, RD50, gamma irradiation.
INTRODUCTION
Rice (Oryza sativa L.) is an important staple food for more than 50% of the world’s population (FAO, 2012), with the bulk of the production globally based in Asia, which is also the largest consumer (FAO, 2018). In Africa, and more specifically in Kenya, there is overreliance on imports with only 20% of the consumed rice grown locally (FAO, 2021). There are many constraints toward increased rice productivity, majority of which can be dealt with by breeding of high productive cultivars, resistance to biotic, and abiotic stresses.
Mutation breeding is a powerful method of rapidly introducing novel genetic diversity from which favorable traits can be selected (Till et al., 2014). Mutation breeding scheme is based on induction of mutation, detection, fixation, development of mutant line and the release of new mutant cultivars (Chepkoech, 2018; FAO/IAEA, 2021). More than 3,200 mutants have been released with over 25% of the mutants being rice varieties (FAO/IAEA, 2021).
In Kenya, mutation breeding has been applied in a few crops including wheat, cassava, potato, pigeon peas and cowpeas (Chepkoech et al., 2020; FAO/IAEA, 2021; Kinyua and Okwaro, 2021; Njau et al., 2005). To introduce mutation breeding technique in Kenyan rice cultivars, there is a need to determine the appropriate doses for the local varieties. This can be achieved through radio-sensitivity study of the plant materials to be utilized in breeding (FAO/IAEA, 2018; Solim and Rahayu, 2021; Ulukapi and Ozmen, 2018). Radio-sensitivity differs depending on plant species, varieties, plant part and water content of the plant material to be irradiated (Kant et al., 2020; Solim and Rahayu, 2021; Toker et al., 2005). Effective mutations have been achieved at lethal dose 50% (LD50), where half of the materials irradiated die, and where 30% or 50% of growth reduction (RD30 and RD50) is achieved (Al-Azab, 2013; FAO/IAEA, 2018; Kant et al., 2020). At these optimal dosages there is maximal probability of getting beneficial mutations with minimal impact on the genome (Al-Azab, 2013; Kant et al., 2020).
This study was aimed at determining the optimal dosages of three most popular, locally adapted rice varieties in Kenya, Basmati 370, Basmati 217, ITA310 and a newly released variety, Komboka.
METHODOLOGY
Plant and treatment with mutagen
The genotypes used in the study were four locally adapted lowland rice varieties. These included Komboka, Basmati 370, Basmati 217 and ITA310 which were obtained from the Kenya Agricultural and Livestock Research Organization (KALRO), Mwea Research Center. Average moisture content of seeds prior to radiation was 11%.
Radio-sensitivity protocol by FAO/IAEA (2018) with a few modifications was adopted for the study. Two experiments were conducted at varied time points. In the first experiment, eight gamma ray dosages, that is, 0, 100, 150, 200, 250, 300, 400, and 500 Gy were used (dosages adopted from Lee et al., (2019) with a few modifications). Approximately 60 seeds were randomly drawn from the seed lots and placed in labeled brown paper envelopes to constitute 8 dosages and placed in the plastic jar on the irradiator.
In the second experiment, eleven portions each containing 60 seeds were randomly drawn from the same seed lots as the first and were treated with eleven gamma ray dosages from 0 to 500 Gy at 50 Gy intervals; 0, 50, 100, 150, 200, 250, 300, 350, 400, 450 and 500 Gy.
The irradiation was conducted using 60Co Gammacell 220 irradiator emitting 2.25 KGy/h belonging to the KALRO Biotechnology Research Institute, Muguga, Kenya.
Planting of the M1 seeds
First experiment
The irradiated seeds were immediately pre-germinated according to Kega et al. (2015) and PHILRICE (2007)with some modifications. Seeds were soaked in water for 24 h then transferred and incubated in moist paper towels contained in plastic containers. Germination percentage was recorded on the 4th day after initiation of pre-germination step, when the radicle had emerged. The germinated seeds were sown in completely randomized design in 160-cell plastic seedling trays filled with soil collected from rice fields. Prior to seedlings transfer, the soil was mixed thoroughly with concentrated phosphate-base fertilizer (diammonium phosphate, DAP). Thereafter, the trays were placed on water pods in a greenhouse at the KALRO, Mwea Research Center.
Second experiment
The irradiated seeds were soaked in water for 24 h and immediately sown in soil media in order of increasing gamma ray dosage according to Mba et al. (2010), with three replications sown in different trays. Irrigation was done on daily basis. Planting of the imbibed irradiated seeds was carried out in a greenhouse at the KALRO, Mwea Research Center.
Data collection
Data collected on the study included germination percentage, rate of seedling emergence, survival rates, and seedling height according to Solim and Rahayu (2021)and Gupta et al. (2021). Germination percentage was recorded on the 4th day after initiation of the pre-germination step. Data on seedling emergence was recorded on the 7th day after sowing (7 DAS) in the first experiment and 11 DAS in the second experiment (due to some delays in emergence of Basmati 370 and Komboka varieties). Seedling survival rate and seedling height were recorded at 14 and 21 DAS in both experiments. LD50 was calculated from the emergence and survival rates, while the median reduction dose 50 (RD50) and reduction dose 30 (RD30) (Lee et al., 2019)were calculated by analyzing plant height at 21 DAS according to Gupta et al. (2021)and Kant et al. (2020).
Statistical analysis
The data was entered in Excel and the analysis of variance computed in R software. Tukey HSD test (P<0.05) was used in mean separation of the significant groups from the ANOVA table. Data on seedling germination percentage, rate of emergence, survival rate and seedling height were fitted in regression models using Curve Expert Professional software version 2.7.3 available at https://www.curveexpert.net/. On the regression models, seedling emergence, survival rate and seedling height were the dependent variables while the gamma ray dosage was the independent variable. GraphPad prism 9 software version 9.3.1 (available at https://www.graphpad.com/) was utilized in combining data of the dependent and independent variables for the varied varieties in single plots.
RESULTS AND DISCUSSION
Germination percentage
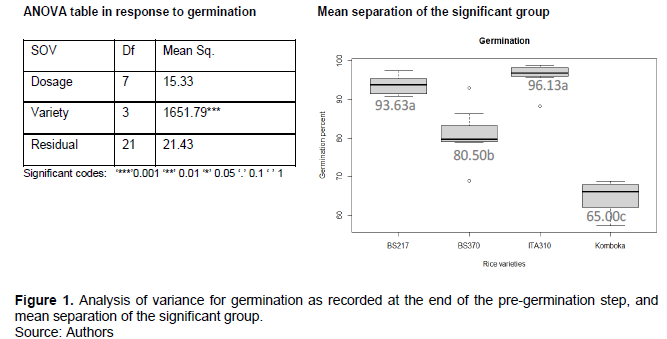
In the first experiment, data on germination was recorded when the primary root or radicle had emerged from the cotyledon. This was done on the 4th day after initiation of pre-germination step. Analysis of variance in response to germination rate was computed in R software, followed by mean separation of the significant group using Tukey HSD at P<0.05. As depicted in the ANOVA Figure 1, gamma irradiation had no significance effect on germination, but there were significant varietal differences not associated with radiation. The results were consistent with previous studies on rice (Harding, 2012; Singh et al., 1998), common beans (Ulukapi and Ozmen, 2018)and groundnuts (Mondal et al., 2017), where the authors reported no significant effects of gamma irradiation on seedling germination.
As shown in Figure 1, ITA310 variety had the highest germination percentage, followed by Basmati 217 and Basmati 370 in that order, with Komboka having the lowest germination percentage.
Germination percentages for ITA310 (96.13%) and Basmati 217 (93.63%) were not significantly different, but were significantly different from those of Basmati 370 (80.50%) and Komboka (65%). In addition, Basmati 370 and Komboka showed significantly different germination percentages.
Effect of varied gamma ray dosage on seedling emergence
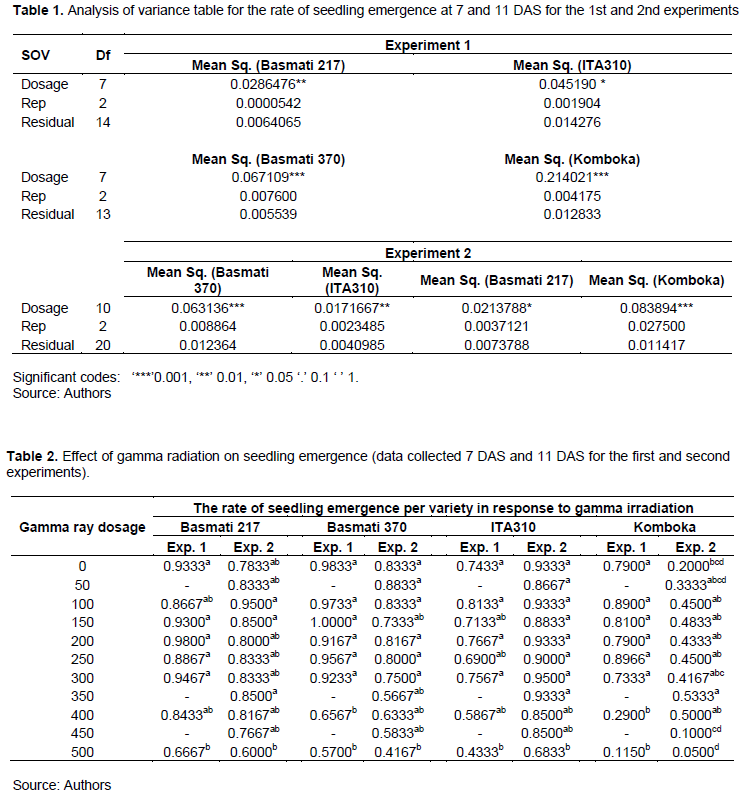
The data on the rate of seedling emergence was subjected to analysis of variance per variety. From the ANOVA Table 1, gamma radiation caused significant effect on the rate of seedling emergence in both experiments and in varieties.
Mean separation for the significant group was carried out using Tukey HSD at P<0.05 on R software. As shown in Table 2, high gamma ray dosages from 400 Gy caused significant reduction on the rate of seedling emergence. Variation in radiation sensitivity among varieties was evident. In the first experiment, seedlings emanating from seeds irradiated at 400 and 500 Gy showed significant reduction in seedling emergence for Komboka and Basmati 370, while for Basmati 217 and ITA310 varieties, significant reduction in emergence was achieved at 500 Gy. In the second experiment, there was significant reduction in emergence on Basmati 370 and ITA310 mutants at high dosage of 500 Gy, but this was not the case for Basmati 217. Our results were consistent with similar studies in rice by Cheema and Atta (2003), where they reported no significant effect of radiation on seedlings emergence at lower dosages. At high dosage, gamma radiation causes generation of free radicals in the irradiated material, causing metabolic disorders that ultimately cause growth retardation of the seedlings(Kumar et al., 2013). It has also been attributed to reduced enzyme activity and inhibition of auxins in seeds exposed to high gamma ray doses (Kant et al., 2020; Kumar et al., 2013). In the context of this study, it is possible that even though the seeds germinated, the seedlings vigor was greatly depressed by the inhibitory effects of deleterious mutations at higher gamma ray dosages causing low emergence.
In the second experiment, very low rates of emergence were observed for Komboka variety, with only 20% of the sowed seeds emerged from the non-irradiated group. For this variety, the rate of seedling emergence increased with increasing gamma ray dosage, with the highest rate at 350 Gy, above which, it reduced significantly to 10 and 5% at 450 and 500 Gy, respectively. Similar findings have been documented by Kim et al. (2000)on Chinese cabbage and radish, where seedling emergence was significantly higher at low dosages compared to the non-irradiated samples.Abdel-Hady et al. (2008)in their review paper indicated that the stimulatory effect of low dosages of gamma ray radiation on germination is attributed to activation of RNA and protein synthesis.
Effect of gamma ray radiation on seedling survival
The rate of seedling survival was recorded on 14 and 21 DAS. In the first experiment (as shown in Figures 6 and 7 in the Appendix), at 21 DAS, most of the entries including the controls had dried up due to some external factors (so the data at 21 DAS was skewed). Therefore, in the analysis of variance and estimation of LD50, data on emergence and survival at 7 and 14 DAS were utilized. In the second experiment, data on emergence at 7 DAS and survival at 14 and 21 DAS were included in the analysis.
In both experiments, gamma radiation caused significant reduction on seedling survival rates as shown in Tables 3 and 4. This is consistent with similar studies in common beans, cowpeas, groundnuts and rice (Harding, 2012; Kang et al., 2020; Mondal et al., 2017; Ulukapi and Ozmen, 2018). DAS group in the ANOVA Table 3 included the rate of emergence at 7 and 11 DAS (for the first and second experiments, respectively), as well as seedling survival rates at 14 and 21 DAS. DAS group was only significantly different for Basmati 370 in the first experiment, indicating that the seedling emergence rate was significantly different from seedling survival rate for this variety. However, it was not significantly different on Basmati217, ITA310 and Komboka varieties.
Upon computing the mean separation of the significant DAS for Basmati 370, the number of seedlings at 14 DAS was significantly reduced compared to what was recorded as emergence at 7 DAS, implying that radiation significantly affected this variety’s seedling survival.
Mean separation of the dosage group is presented in Table 4. Of the four varieties, Basmati 370 was the most sensitive, with gamma ray dosages from ≥300 and ≥350 Gy in the first and second experiments, respectively, causing significant reduction in seedling survival. Significant reduction on seedling survival rate for ITA310 mutants was reported at 400 and 500 Gy in the first and second experiments, respectively. For Basmati 217, mutant seedlings emanating from seeds irradiated at 100 Gy had the highest survival rate at 93.89%, this was significantly higher than the other treatments in the group, including the non-irradiated samples. Among the four varieties, Basmati 217 was the least sensitive to gamma irradiation, with significant reduction on survival rate only at 500 Gy in both experiments.
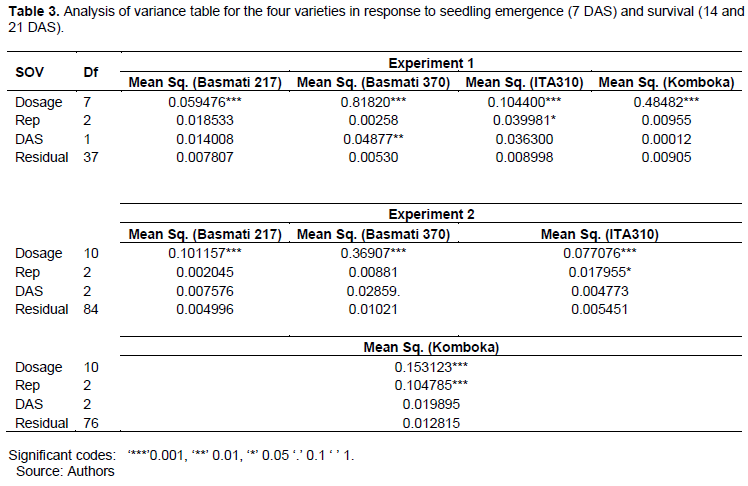
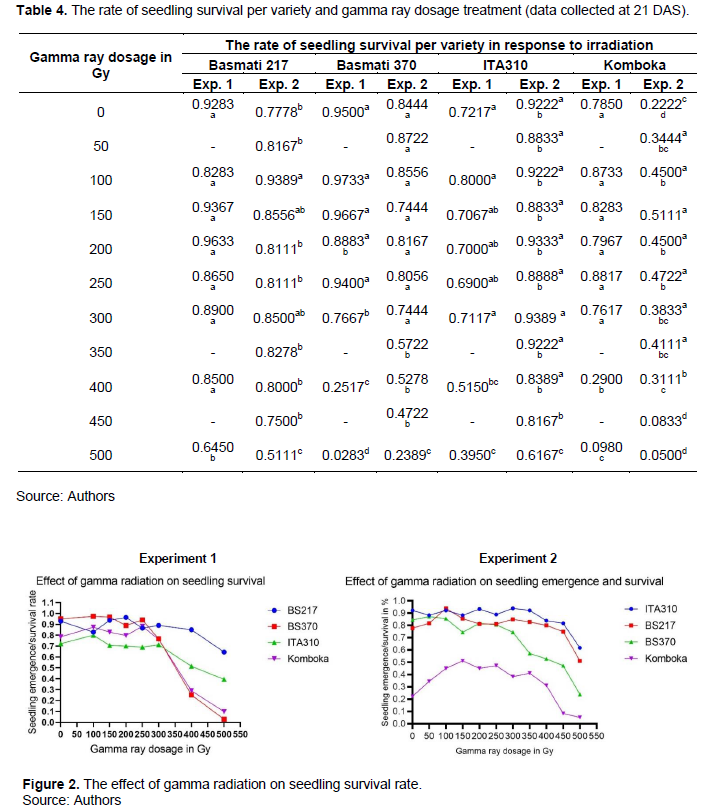
In the first experiment, Komboka mutants at 400 and 500 Gy had significantly reduced survival rates compared to the non-irradiated samples. But in the second experiment, significantly higher survival rates were exhibited by doses between 100 and 250 Gy compared to the control. In the second experiment, at 350 Gy, seedling survival reduced from 53.33% recorded as emergence rate at 11 DAS to 41.11% at 21 DAS as shown in Tables 2 and 4, respectively.
Data on seedling emergence and survival was used in generating the plots on Figure 2 using GraphPad Prism 9 software. As shown in Figure 2, Basmati 370 in the first and second experiments, and Komboka in the first experiment were most affected by gamma radiation. From 300 Gy, the number of seedlings reduced with the increasing gamma ray dosage. Dosages at 400 and 500 Gy significantly caused stunted growth in most of the seedlings, with 500 Gy causing more than 90% seedling death in Komboka and Basmati 370 varieties in the first experiment. On the contrary, ITA310 and Basmati 217 were less sensitive to irradiation in both experiments, with survival rates above 50% at 500 Gy relative to the 0 Gy dose. The results are consistent with similar studies in varied crops, including rice and cowpeas, where gamma radiation above 300 Gy caused significant reduction on seedling survival (Harding, 2012; Kadhimi et al., 2016; Kang et al., 2020).
Seedling emergence and survival at 0 Gy was used as the reference (converted to 100%), where all the other doses were compared against. From the output in Table 4, the reduction on the rate of seedling emergence and survival, relative to the control (dose 0Gy) was calculated for each dose and variety, then fitted on regression models using Curve Expert Professional version 2.7.3. where estimation of LD50 was done.
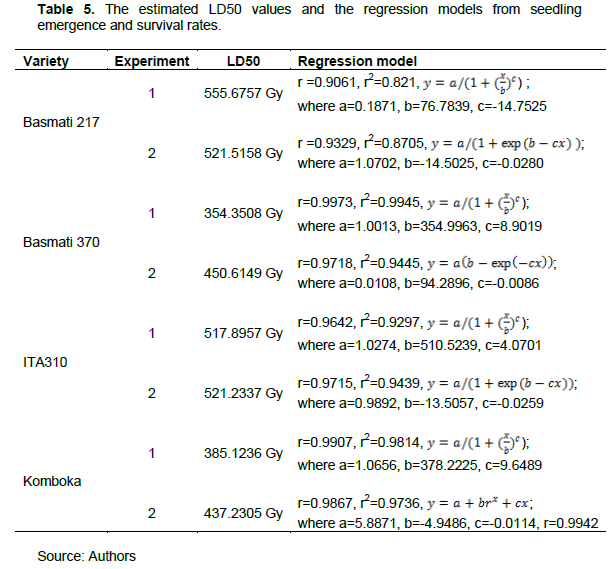
From the curve expert output presented in Table 5, more than 80% of the data fitted the regression models perfectly as depicted by the coefficient of determination (r2). LD50 values across varieties ranged from 354.3508 to 555.6757 Gy. Basmati 217 had the highest LD50 values, thus the least radio-sensitive variety with LD50 ranging from 521.5158 to 555.6757 Gy, followed by ITA310 with LD50 ranging from 517.8957 to 521.2337 Gy. Komboka rice variety had its LD50 ranging from 385.1236 to 437.2305 Gy, while Basmati 370 had the lowest LD50 ranging from 354.3508 to 450.6149 Gy.
A low LD50 consistent with that of Basmati 370 has been reported on MRQ74, MR269 and White Ponni rice varieties on similar studies, which had LD50 at 365.1061, 351.3429 and 354.8000 Gy, respectively (Kadhimi et al., 2016; Ramchander et al., 2015).
Varieties with low sensitivity, Basmati 217 and ITA310 had higher LD50 values than the highest value of the thirteen cultivated varieties in Sierra Leone studied by Harding (2012), whose LD50 ranged from 345 to 423 Gy. These were also slightly lower than Galon rice genotype in Bangladesh which was reported at 575 Gy by Gupta et al. (2021). Higher LD50 values in rice varieties have also been reported by Solim and Rahayu (2021) for Mira-1 and Bastari in Indonesia whose LD50 ranged from 521.40 and 663.68 Gy. Suliartini et al. (2020) in a similar study on four lowland rice varieties, reported an LD50 value of 518 Gy for one of the varieties, Inpago Unram-1.
The outcomes are consistent with the varieties with the lowest sensitivity in our study, Basmati 217 and ITA310 whose LD50 ranged from 517.8957 to 555.6757 Gy.
Effect of radiation on seedling height at 14 and 21 DAS
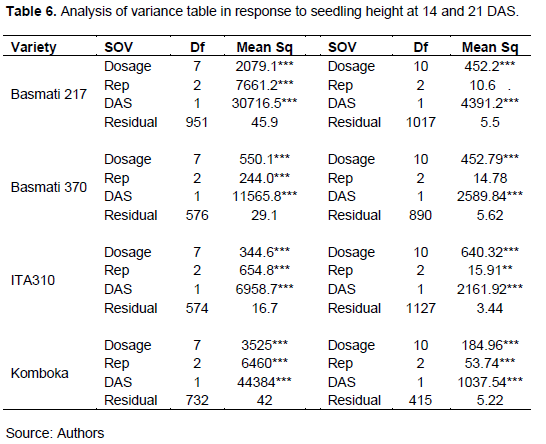
Seedlings height was measured and recorded at 14 and 21 DAS. Analysis of variance was computed in R software, from which the output in Table 6 was generated. As shown in Table 6, dosages, replication and DAS groups were significantly different for all the varieties in the first experiment. In the second experiment, dosage and DAS groups were significantly different in all varieties, while the replication group was significantly different in ITA310 and Komboka varieties. Mean separation using Tukey HSD at P<0.05 was computed for the significant groups in R software. Seedling height at 21 DAS was significantly greater than the height at 14 DAS.
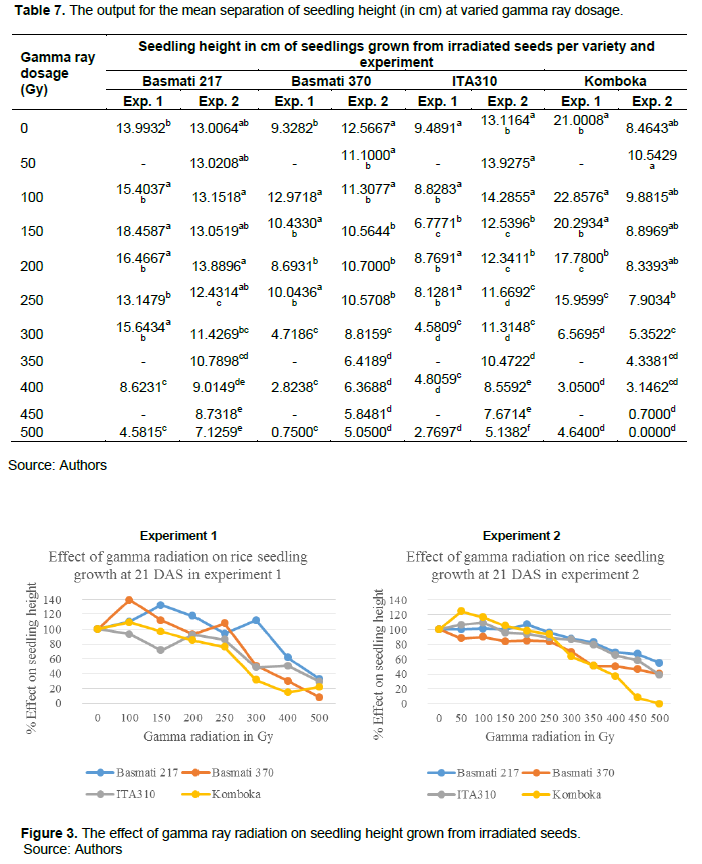
As shown in Table 7, in the first experiment, Basmati 217 and Komboka were significantly taller at lower dosages than other entries in the first and second experiments. In addition, at 100 and 150Gy doses of Basmati 370 and Basmati 217, respectively, mutants were significantly taller than the non-irradiated (0Gy) samples as presented in Table 7 and Figure 3. The same was reported in the second experiment on ITA310 mutants at 50 and 100 Gy. Our results were consistent with similar studies in rice, chickpeas and Chinese cabbage (Gupta et al., 2021; Kadhimi et al., 2016; Kim et al., 2000; Toker et al., 2005). In rice, increased seedling height relative to non-irradiated samples at 100 and 200 Gy was reported by Gupta et al. (2021) and Kadhimi et al. (2016). Kim et al. (2000) showed increased seedling height in Chinese cabbage at low dosages of 4 and 10 Gy. Toker et al. (2005) reported increased seedling growth on chickpeas at 100 and 200 Gy, while Ulukapi and Ozmen (2018) reported enhanced root growth in common beans F16 cultivar at 100 Gy compared to the non-irradiated samples.
As shown in Table 7 and Figure 3, gamma radiation caused significant reduction in seedling height from as low as 150 Gy in experiment 2 for Basmati 370 mutants. The height significantly reduced with increasing gamma ray dosages with variations across varieties as shown in Figure 3, and Figures 4 to 12 in the Appendix. This is consistent with similar studies in various crops (Kadhimi et al., 2016; Kang et al., 2020; Sasikala and Kalaiyarasi, 2010). In the first experiment, significant growth reduction on Basmati 370 was observed from 300 Gy. Radiation caused significant growth reduction on Komboka mutants from ≥250 Gy and ≥300 Gy in the first and second experiments, respectively. For ITA310, mutants were significantly shorter from ≥250 Gy and ≥300 Gy for the second and first experiments, respectively compared to the non-irradiated samples. In the first experiment, ITA310 mutants at 150 Gy were also significantly shorter than the non-irradiated (0Gy) samples, but the mutants at 200 and 250 Gy were not. This is possible because of the random effect of radiation giving different responses (Suliartini et al., 2020). For Basmati 217, significant reduction on seedling height was observed from ≥350 Gy and ≥400 Gy on the second and first experiments, respectively. This placed Basmati 217 as the least sensitive variety and Basmati 370 as the most sensitive variety in our study. The results across varieties on the effect of radiation on seedling height were consistent with reports on similar studies by Harding (2012) and Solim and Rahayu (2021) where significant growth reduction was reported at radiation dosages >300 Gy and >400 Gy, respectively.
The reduction effects on varied growth parameters by gamma radiation have been associated with disruption of cell cycle at G2/M phase, disruption of DNA, RNA, protein, growth hormone synthesis (Kang et al., 2020; Solim and Rahayu, 2021; Ulukapi and Ozmen, 2018), disturbance in hormonal balance and enzyme activity (Kant et al., 2020).
The output in Table 7 was converted as a percentage of the non-irradiated samples and used in generation of growth curves presented in Figure 3. As shown in Figure 3, similar pattern on the effect of radiation on individual variety is notable in both experiments.
The data on seedling height at 21 DAS was fitted on regression models and utilized in estimation of RD30 and RD50 values. Data on seedling height fitted well on regression models as shown in Table 8, with coefficient of determination ≥0.9 for all the varieties, except for the data of ITA310 in the first experiment, where the r2 was 0.5410, indicating that only 54.10% of the ITA310 data set in the first experiment fitted the model. This can be explained by the fact that only a few plants across dosages for this variety survived per entry to 21 DAS data point (as shown in Figure 7 in the Appendix), so the data on seedling height for this variety was skewed.
From RD30 and RD50 values as presented in Table 8, the least radio-sensitive variety was Basmati 217 followed by ITA310, while the most sensitive variety was Komboka followed by Basmati 370. For Basmati 217, RD30 values ranged from 417.0106 to 425.9882 Gy, while RD50 values ranged from 496.2296 to 531.1608 Gy. For ITA310, RD30 values ranged from 330.3820 to 392.4310 Gy, while RD50 values ranged from 418.3331 to 470.0900 Gy. Basmati 370 had its RD30 ranging from 286.9128 to 311.0258 Gy, while RD50 ranged from 368.3795 to 421.8008 Gy. Komboka’s RD30 ranged from 266.5063 to 292.5661 Gy, while RD50 ranged from 335.3659 to 351.3659 Gy.
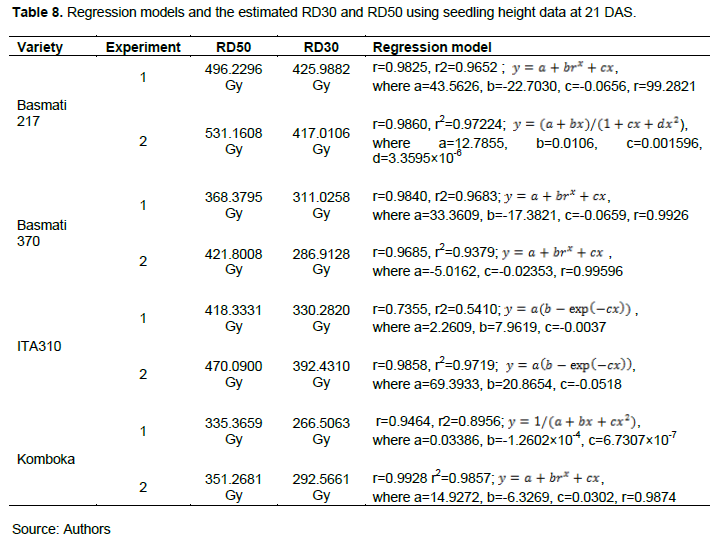
RD50 values in this study are consistent with the growth reduction 50 values on rice reported by FAO/IAEA (2018), which ranged between 350 and 500 Gy. LD50 and RD30/RD50 values differed slightly across environments consistent with similar studies on rice by Kant et al. (2020) where the researchers studied the effect of gamma radiation in vivo and in vitro.
CONCLUSION
In this study, gamma radiation on rice seeds did not have significant effect on seedling germination, but significantly affected seedling emergence, survival and growth. The effect of irradiation on these growth parameters varied across varieties and slightly across experiments/ environments. Basmati 370 and Komboka were the most sensitive to irradiation, ITA310 was moderate, while Basmati 217 was the least sensitive. At low doses of 50, 100, and 150 Gy, irradiation caused increased seedling vigor on Basmati 217, Basmati 370, and ITA310. Across varieties, seedling survival and height reduced with increasing doses from 300 and 350 Gy, respectively. The effective doses were determined for each variety as their individual LD50, RD30 and RD50 values. LD50 values ranged from 354 to 556 Gy. Basmati 370 had the lowest LD50 values at 354 and 451 Gy, followed by Komboka at 385 and 437 Gy, then ITA310 at 518 and 521 Gy, while Basmati 217 had the highest LD50 values at 522 and 556 Gy in the first and second experiments, respectively. RD30 values ranged from 267 to 426 Gy. Komboka had the lowest RD30 values at 267 and 293 Gy followed by Basmati 370 at 290 and 311 Gy, then, ITA310 at 330 and 392 Gy, while Basmati 217 had the highest values at 417 and 426 Gy in the first and second experiments, respectively. RD50 values ranged from 335 to 531 Gy. Komboka had the lowest RD50 values at 335 and 351 Gy, followed by Basmati 370 at 368 and 422 Gy, then ITA310 at 418 and 470 Gy, while Basmati 217 had the highest values at 496 and 531 Gy in the first and second experiments, respectively. Radio-sensitivity study was successful in determining the effective doses for selected varieties. In the rice mutation breeding program, it is recommended to adopt gamma ray doses between RD30 and LD50 in this study as the optimal doses for each variety to cause the desired mutations.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
This study was funded by the West Africa Center for Crop Improvement (WACCI) and German Academic Exchange Service (DAAD). Kenya Agricultural and Livestock Research organization (KALRO) Mwea, provided the facility where the study was conducted.
REFERENCES
|
Abdel-Hady MS, Okasha EM, Soliman SSA, Talaat M (2008). Effect of Gamma Radiation and Gibberellic Acid on Germination and Alkaloid Production in Atropa belladonna L. Available at: . |
|
|
Al-Azab KF (2013). Improving wheat for drought tolerance by using hybridization and mutation breeding procedures. Cairo University Egypt. Available at: |
|
|
Cheema AA, Atta BM (2003). Radiosensitivity studies in Basmati rice. Pakistan Journal of Botany 35(2):197-207. |
|
|
Chepkoech E (2018). Application of gamma induced mutation in breeding for bacteria Wilt (Ralstonia solanacearum) disease resistance in potato (Solanum tuberosum L.). University of Eldoret. Available at: |
|
|
Chepkoech E, Kinyua M, Ochuodho J, Kiplagat O, Bado S, Kinyua Z (2020). Application of Gamma Induced Mutation in Breeding Potato for Bacterial Wilt Disease Resistance. International Journal of Pathogen Research 5(3):28-38. |
|
|
FAO/IAEA (2018). Manual on mutation breeding. In Spencer-Lopes MM, Forster BP, Ljupcho Jankuloski (eds.), Food and Agriculture Organization of United Nations (3rd ed.). FAO/IAEA. Available at: |
|
|
FAO/IAEA (2021). Mutation Breeding. Joint FAO/IAEA Programme, Nuclear Techniques in Food and Agriculture. Available at: |
|
|
Food and Agriculture Organization (FAO) (2012). Trends in the crop sector. FAO Statistical Yearbook. pp. 182-197. Available at: |
|
|
Food and Agriculture Organization (FAO) (2018). FAO Rice Market Monitor. FAO, XXI(1). Available at: |
|
|
Food and Agriculture Organization (FAO) (2021). FAOSTAT 2021. FAO. Available at: |
|
|
Gupta R, Mofazzal Islam M, Nahar Begum S, Akram W, Shafiqul Islam M (2021). Determination of Lethal Dose of Rice (Oryza Sativa L.) Genotypes By Radio Sensitivity Test. Plant Archives 21(1):1912-1916. |
|
|
Harding S (2012). Effect of Gamma Rays on Seed Germination, Seedling Height, Survival Percentage and Tiller Production in Some Rice Varieties Cultivated in Sierra Leone. American Journal of Experimental Agriculture 2(2):247-255. |
|
|
Kadhimi AA, ALhasnawi AN, Isahak A, Ashraf MF, Mohamad A, Yusoff WMW, Zain CRCM (2016). Gamma radiosensitivity study on MRQ74 and MR269, two elite varieties of rice (Oryza Sativa L.). Life Science Journal 13(2):85-91. Available at: Gamma radiosensitivity study on MRQ74 and MR269, two elite varieties of rice (Oryza Sativa L.). Available at: |
|
|
Kang R, Seo E, Kim G, Park A, Kim WJ, Kang SY, Ha BK (2020). Radio sensitivity of cowpea plants after gamma-ray and proton-beam irradiation. Plant Breeding and Biotechnology 8(3):281-292. |
|
|
Kant A, Chakraborty NR, Das BK (2020). Immediate radiation effects and determination of optimal dose of gamma rays on non-basmati aromatic rice (Oryza sativa L.) of eastern India. Journal of Experimental Biology and Agricultural Sciences 8(5):586-604. |
|
|
Kega MV, Gikonyo WE, Catherine WM, Macharia MKJ, Lucy M (2015). Rice cultivation manual (Issue 978). Available at: |
|
|
Kim JS, Baek MH, Lee YG, Jung KH (2000). Effects of low dose gamma-ray radiation on the seed germination and physiological activity of vegetable crops. Proceedings of the KNS Autumn Meeting. Available at: |
|
|
Kinyua MG, Okwaro H (2021). Effect of Gamma Irradiation Mutagenesis On Diversity of Cassava Tissue Culture Plantlets. International Journal of Scientific and Engineering Research 12(2):809-817. Available at: |
|
|
Kumar DP, Chaturvedi A, Sreedhar M, Aparna M, Venu-Babu P, Singhal RK (2013). Gamma radiosensitivity study on rice (Oryza sativa L.). Pelagia Research Library 3(1):54-68. Available at: View |
|
|
Lee SI, Jae-Wan P, Jin-Baek K, Si-Yong K, Hong-II C, Sung-Min H (2019). Evaluation of Radiosensitivity of Rice (Oryza sativa L.) Seeds Exposed to Proton Beams and Gamma Rays and Investigation of Optimal Doses for Mutation Induction. Journal of Radiation Industry 13(3):173-178 Available at: . |
|
|
Mba C, Afza R, Bado S, Jain SM (2010). Induced Mutagenesis in Plants Using Physical and Chemical Agents. Plant Cell Culture: Essential Methods 20:111-130. Available at: https://doi.org/10.1002/9780470686522.ch7 |
|
|
Mondal S, Petwal VC, Badigannavar AM, Bhad PG, Verma VP, Goswami SG, Dwivedi J (2017). Electron beam irradiation revealed genetic differences in radio-sensitivity and generated mutants in groundnut (Arachis hypogaea L.). Applied Radiation and Isotopes 122:78-83. |
|
|
Njau PN, Kinyua MG, Kimurto PK, Okwaro HK, Maluszynski M (2005). Drought Tolerant Wheat Varieties Developed Through Mutation Breeding Technique. Journal of Agriculture, Science and Technology 7(1):18-29. |
|
|
PHILRICE (2007). Seed soaking, incubation, and sowing. Philippine Rice Research Institute, Handout series. |
|
|
Ramchander S, Ushakumari R, Arumugam Pillai M (2015). Lethal dose fixation and sensitivity of rice varieties to gamma radiation. Indian Journal of Agricultural Research 49(1):24-31. |
|
|
Sasikala R, Kalaiyarasi R (2010). Sensitivity of rice varieties to gamma irradiation. Electronic Journal of Plant Breeding 1(4):845-889. Available at: |
|
|
Singh S, Richharia AK, Joshi AK (1998). An Assessment of Gamma Ray Induced Mutations in Rice (Oryza sativa L.). Indian Journal of Genetics and Plant Breeding 58(4):455-463. Available at: |
|
|
Solim MH, Rahayu S (2021). Radiosensitivity of rice varieties of Mira-1 and Bestari mutants using gamma rays irradiation. IOP Conference Series: Earth and Environmental Science 911(2021):012014. |
|
|
Suliartini NWS, Wangiyana W, Aryana IGPM, Sudharmawan AAK (2020). Radiosensitivity and Seedling Growth of Several Genotypes of Paddy Rice Mutants Irradiated with Gamma Rays at Different Doses. International Journal of Horticulture, Agriculture and Food Science 4(6):242-247. |
|
|
Till BJ, Amos EK, Kinyua MG, Bado S, Jankowicz-cieslak J, Huynh OA, Henry I, Comai L, Lagoda P (2014). Mutation induction and reverse-genetics for functional genomics and breeding pp. 5-6. Available at: |
|
|
Toker C, Uzun B, Canci H, Ceylan FO (2005). Effects of gamma irradiation on the shoot length of Cicer seeds. Radiation Physics and Chemistry 73:365-367. |
|
|
Ulukapi K, Ozmen SF (2018). Study of the effect of irradiation (60 Co) on M 1 plants of common bean ( Phaseolus vulgaris L.) cultivars and determined of proper doses for mutation breeding. Journal of Radiation Research and Applied Sciences 11(2):157-161. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


