Full Length Research Paper

ABSTRACT
Previous biochemical and molecular sequence analyses of Xanthomonas campestris pathovar musacearum, the etiological agent of banana Xanthomonas wilt, suggest that it belongs within the species Xanthomonas vasicola (X. vasicola pv. vasculorum and X. vasicola pathovar holcicola. However, the X. vasicola pathovar names were considered invalid according to pathovar naming standards and placed as one X. vasicola species; this was also not helped by the lack of sufficient comparative pathogenicity studies. Hence the proposal to rename X. campestris pathovar musacearum was no longer further supported. This study therefore carried out large scale comparative pathogenicity trial studies on the X. vasicola strains and X. campestris pathovar musacearum on 112 plants for banana and maize, and 84 plants for sugarcane, to establish or support the proper X. vasicola pathovar designations. The study also included nine common plant pathogenic Xanthomonas pathovars and one non-Xanthomonas strain. The six strains of X. campestris pathovar musacearum used in the study caused disease in sugarcane and banana but not on maize. 2 and 4 strains of X. vasicola pathovar vasculorum and X. vasicola pathovar holcicola, respectively were not only pathogenic on maize and sugarcane but each also caused distinct symptoms on maize. X. vasicola pathovar vasculorum caused deformation of the plant while X. vasicola pathovar holcicola caused stunted growth.
Key words: Pathogenicity, X. axonopodis pv. vasculorum, X.campestris pv. musacearum, X vasicola pv. holcicola, X. vasicola pv. vasculorum, Xanthomonas wilt of bananas.
INTRODUCTION
Xanthomonas campestris pv. musacearum (Xcm) is the etiological agent of Banana Xanthomonas wilt and a major threat to the existence of Musa species (bananas and plantain) in East Africa and Ensete ventricosum (enset) in Ethiopia (Biruma et al., 2007). The disease was first discovered in Ethiopia on enset plants (Yirgou and Bradbury et al.,1968) and the first report in a major banana-growing area was in Mukono district, Uganda in 2001 (Tushemereirwe et al., 2004). It has since spread to the neighboring countries of Democratic Republic of Congo (Ndungo et al, 2006), Kenya, Tanzania and Burundi (Carter et al., 2010) and Rwanda (Reeder et al, 2007). Xanthomonas wilt causes severe losses to banana yields and production, thereby devastating livelihoods of millions in Uganda. Symptoms include premature ripening of the fruit, shriveling of male buds, progressive yellowing, and wilting of leaves. When the pseudostem is cut, pockets of yellow bacterial ooze are seen after 15 to 20 min, confirming the presence of the disease (Tinzaara et al., 2006). There are no known resistant cultivars of banana. The current control methods include complete destruction of infect plants/plant materials, use of sterile cutting or harvesting tools and removal of males buds (Tripathi et al., 2010; Biruma et al., 2007).
Recent characterization studies of Xcm isolates through Fatty Acid Methyl Ester (FAME) analysis, Rep-PCR and B gyrase sequencing (Aritua et al, 2008 and Parkinson et al., 2009) have revealed that Xcm is phylogenetically very closely related to strains of X. vasicola pathovars: X. vasicola pv. vasculorum (Xvv) and X. vasicola pv. holcicola (Xvh) that are pathogens on sugarcane and sorghum, respectively (Vauterin et al, 1995). Strains of Xvv were originally classified as X. campestris pv. vasculorum (Vauterin et al, 1995). Aritua et al. (2008) revealed that Xcm did not show much genetic similarity with Xanthomonas campestris pathovars. Aritua et al. (2008) further carried out pathogenicity studies with the X. vasicola pathovars on maize and banana and reported that Xcm not only caused disease in banana but in maize as well. The X. vasicola pathovars (Xvv and Xvh) only caused disease in maize. Maize was included in the study as it is a close relative of sorghum and also the source of isolation for Xvv 206 NCPPB (National Collection of Plant Pathogenic Bacteria). However the current proposed names of the X. vasicola pathovars were considered invalid according to the pathovar naming standards and were placed as one X.vasicola species (Garrity, 2005). The proposal to rename Xcm was no longer further supported. This was not helped by the lack of enough insufficient pathogenicity studies of the X.vasicola species.
The purpose of this study was to provide the much needed substantial data on pathogenicity of the X. vasicola species and Xcm on maize, sugarcane, and banana and to establish or support the current pathovar designations of these particular strains. It was assumed that Xvv and Xvh caused similar symptoms on maize and sugarcane and did not cause disease in banana while Xcm only caused disease in maize and banana. The green house trials also included other plant pathogenic Xanthomonas pathovars and non-Xanthomonas bacteria to bring out the distinct pathogenicity of the X. vasicola strains.
MATERIALS AND METHODS
Bacterial strains
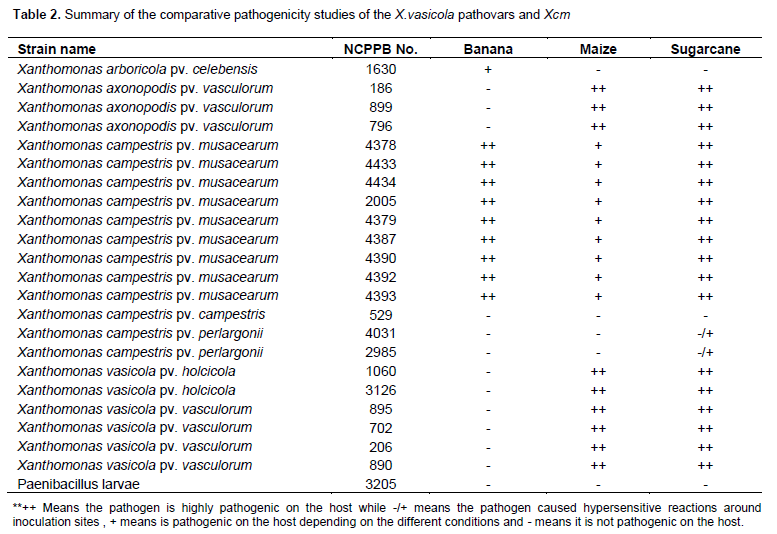
28 bacteria strains from NCPPB (National Collection of Plant Pathogenic Bacteria) were used in this study and are listed in Tables 1 and 2. 9 Xcm, 4 Xvv, 2 Xvh, 2 X.axonopodis pv. vasculorum, one X. arboricola pv. celebensis, 2 X. campestris pv. perlargonii, 3 X. campestris pv. campestris strains and one non Xanthomonas strain Paenibacillus larvae. These strains were also tested and evaluated by PCR using the different available BXW primers (Hodgetts et al., 2014). The bacteria were cultured on YDC (yeast dextrose chalk; (Bacto Agar 15 g/l, yeast extract 10 g/l, CaCO3 20 g/l, D-glucose 20 g/l (dextrose) and distilled/de-ionized water 1000 ml, autoclaved at 121°C for 15 min) media and incubated at 28°C (optimum temperature for Xanthomonas) for 48 h. Bacteria culturing was done 48 h before inoculation of the plants. After 48 h, 10 µl loop of the bacteria was then resuspended in sterile water and the concentration adjusted to 107 CFU/ml, using the spectrophotometer by addition of water or bacteria. The suspension was then inoculated into the plants using sterile 1 ml syringes (Sinclair and Dhingra, 1995).



Pathogenicity trials
Pilot trials
Pilot trials on banana (Dwarf Tropicana and Dwarf Cavendish) and maize were carried out to determine the level/concentration of bacteria inoculums that would enable pathogenicity and to know what symptoms to expect and when to expect them. Four (4) Tropicana and three of Cavendish dwarf banana were also inoculated with the 200 µl of Xcm suspension (NCPPB 2005, 4392) while two Tropicana and three Cavendish dwarf bananas were inoculated with Xvh (NCPPB 1060). Two (2) Tropicana and three Cavendish dwarf bananas were left as controls.
Eight (8) maize plants for each pathovar (Xcm, Xvv, and Xvh) were inoculated with 200 ul of bacterial suspension containing 107CFU/ml of one of eight bacterial strains: Xcm (NCPPB 4344, NCPPB 4378), Xvv (NCPPB 702, NCPPB 795) and Xvh (NCPPB 1060), Xanthomonas axonopodis pv. vasculorum (Xav, NCPPB 796, NCPPB 899. Four maize plants were controls; two treated with sterile water and two were left untreated.
Eight (8) sorghum seedlings were also inoculated with 200 µl of bacterial suspensions of Xcm (NCPPB 4434, NCPPB 4378) and eight with Xvh (NCPPB 1060). Four (4) sorghum seedlings were control plants. Control plants were either left untreated or were inoculated with sterile water. Photographic evidence of symptoms was taken. A pilot study on sugarcane was not attempted due to a shortage of sugarcane plants.
Large-scale pathogenicity trials
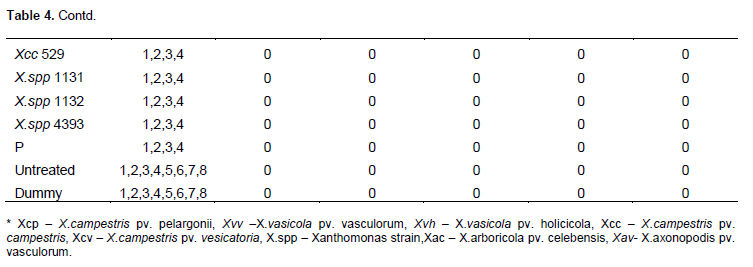
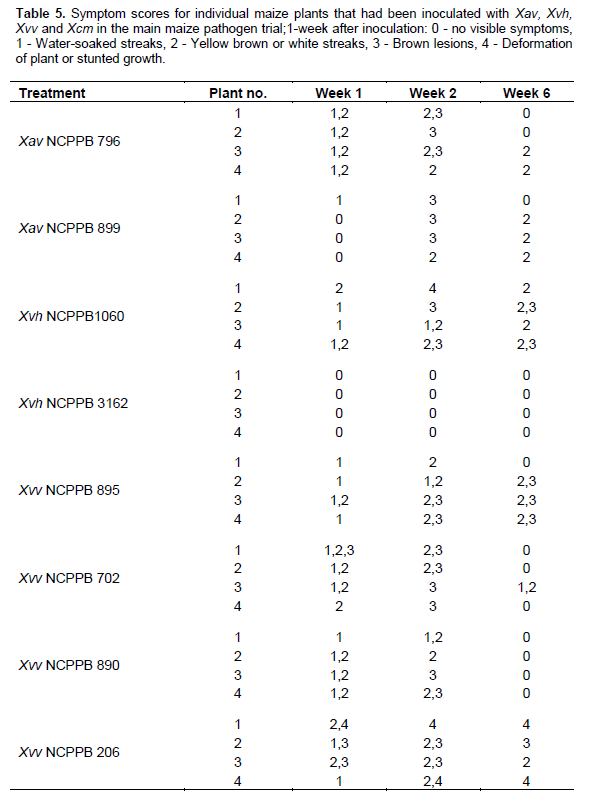
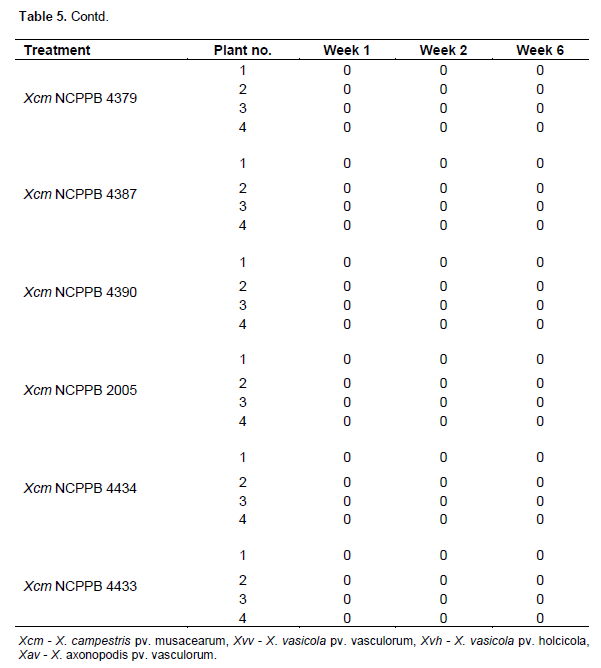
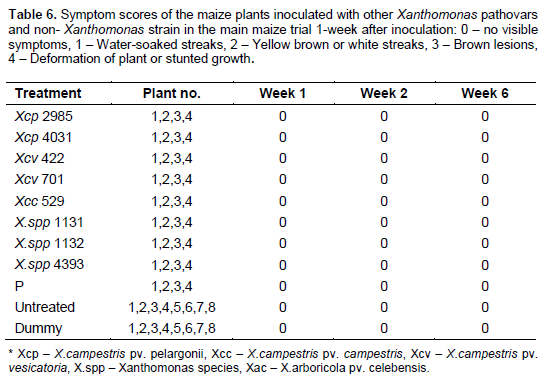
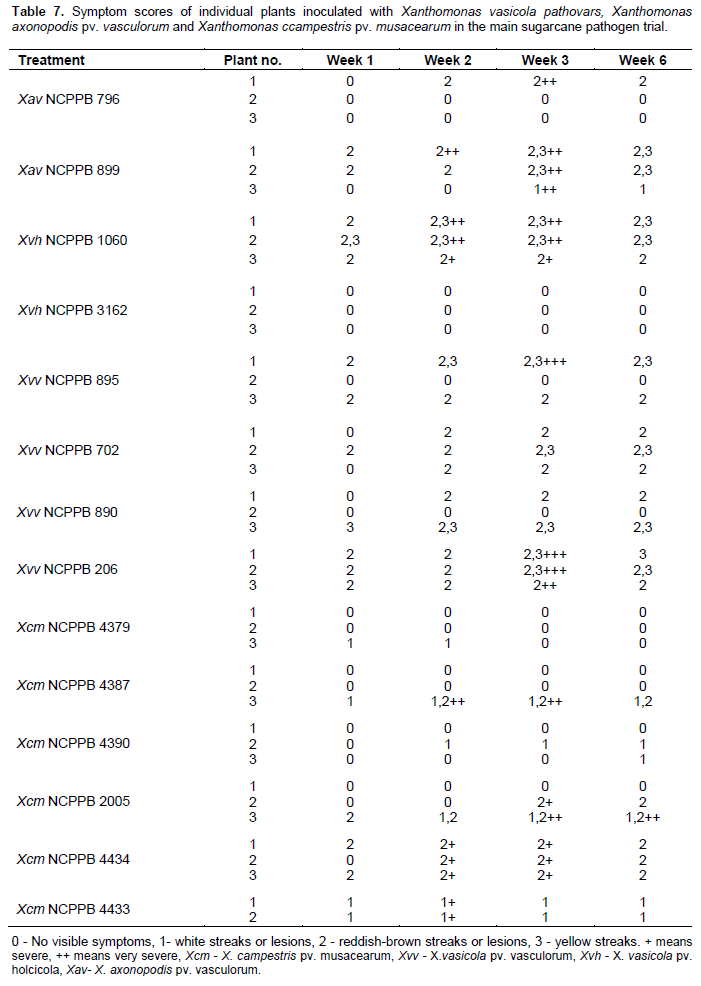
One hundred and twelve (112) plants of maize (var. cisko) and 112 of banana (Dwarf Cavendish) were used in the pathogenicity trial (Table 1 for experiment design). 8 maize plants and 8 banana plants were left untreated and 8 plants of each were inoculated with sterile water (16 control plants) for each trial. For the sugarcane trial, we were only able to acquire 84 plants and therefore included 12 control plants rather than 16. As in the pilot trial, the plants were inoculated with 200 µl of bacterial suspension adjusted to 107 CFU/ml, using sterile 1ml syringes and hypodermic needles. 100 µl of bacterial suspension was also spread on Yeast Dextrose Chalk (YDC) media, plates incubated at 28°C for 48 h to confirm the viability of the inoculums. The greenhouse temperature was set between 28 to 30°C. The inoculated plants were assessed every day for the appearance of symptoms and photographs were taken of any suspected visible symptoms. For the banana pathogenicity trial, symptom severity was scored as follows; 0 - no visible symptoms, 1 - slight wilting/folding of lower leaves, 2 - pronounced wilting/yellowing of most leaves, 3 - pronounced necrosis of the whole plant, 4 - complete death, rotting of the plant. For the maize pathogenicity trial, symptom severity was scored as follows of disease; 0 - no visible symptoms, 1 - water-soaked like streaks, 2 - yellow or brown or white streaks, 3 - brown lesions and 4 - deformation of plant or stunted growth. For sugarcane main trial symptom scores of the leaves were scored according to the type of symptom rather than severity of the disease: 0 - no visible symptoms, 1 - white streaks or lesions, 2 - reddish-brown streaks or lesions, 3 - yellow streaks.
Re-isolation of the pathogens was done from plants inoculated with Xcm, Xvv and Xvh irrespective of whether the plants exhibited symptoms or not. One or two leaves or leaf stalks of each treatment were picked, and the part of the leaf bordering between the diseased and healthy was cut up in to pieces (0.1 to 0.3 g). The leaf pieces were then soaked, crushed in 1 ml of phosphate buffered saline solution (PBS) and left to stand for at least 10 min to allow bacteria to ooze out. 100 µl of the crushed leaf-PBS solution was then placed and spread on YDC media, incubated at 28 to 30°C for 48 to 72 h.
Confirmation of the isolates was done by extracting DNA using the QiAamp DNA Mini Kit following the manufacturer’s protocol and then testing them with the PCR assays specific for Xcm (GspDmFR), (Adriko et al., 2011) and those that are able to detect Xvv and Xvh strains (BXW1F/3R, Lewis Ivey et al., 2010 and Xcm44FR (Adikini et al., 2011). Re-isolation from plants that had been inoculated with other Xanthomonas strains was also done and identified visually; colonies that looked like Xanthomonas were assumed to be of the same strain that had been inoculated into that particular plant, since no Xanthomonas was isolated from control plants.
RESULTS
Banana and maize pilot trial
Two hundred microliters (200 µl) of bacterial inoculum at a concentration of 107 CFU/ml was enough to cause virulence in maize and banana. All control plants; untreated and those inoculated with sterile water remained healthy, that is no leaf wilting, streaks, necrosis, or yellowing. The 3 Dwarf Cavendish and 4 Dwarf Tropicana banana plants that had been inoculated with Xcm (NCPPB 4392 and NCPPB 2005) developed necrotic leaf symptoms, wilting 3 weeks after inoculation and eventually necrosis of whole plant by the end of trial (5 weeks after inoculation).The 2 Tropicana and 3 Cavendish dwarf banana plants that were inoculated with the Xanthomonas vasicola pathovar Xvh (NCPPB 1060) had no visible symptoms and remained healthy.
The 4 maize control plants remained healthy, that is the leaves showed no signs of lesions, chlorosis, or streaks and no folding of stems. The 8 maize plants that had been inoculated for each Xanthomonas vasicola pathovars Xvv (NCPPB 702, NCPPB 795) and Xvh (NCPPB 1060) and Xanthomonas axonopodis pv. vasculorum (NCPPB 796, NCPPB 899) strains exhibited leaf chlorosis, yellow-brown or water soaked streaks or lesions on the leaves, usually beginning in the center of the leaf, 1 week after inoculation. The most severe symptoms included deformation of the plant and retarded growth by Xvv and Xvh (NCPPB1060) 5 weeks after inoculation respectively. Out of the 8 maize plants inoculated with Xcm (NCPPB 4434, NCPPB 4378), only one plant (inoculated with Xcm NCPPB 4378) showed identical symptoms as those seen on plants inoculated with Xvv; the leaves only had yellow-brown streaks. All the maize controls remained healthy and did exhibit any leaf lesions or wilting. The pilot trial for sorghum was unsuccessful as all the plants including those that had been inoculated with Xcm and Xvh did not show any signs of disease. Unfortunately, fresh sorghum seedlings were still not available in time for the main large scale trial.
Banana large scale pathogenicity trial
The 16 banana control plants remained healthy. Out of the 24 plants that had been inoculated with Xcm, 20 showed severe typical symptoms of Xanthomonas wilt of bananas (Figure 1 a-c) and refer to Table 3). The 4 plants that were inoculated with Xanthomonas arboricola pv. celebensis (Xac) that causes Banana blood disease remained healthy and re-isolation of the bacteria was unsuccessful. This strain may have lost its ability to cause disease in banana as it had been got from -80°C storage. The 24 plants that had been inoculated with the Xanthomonas vasicola pathovars (16 with Xvv and 8 with Xvh) also remained healthy and re-isolation of these strains was successful. The 8 plants that had also been inoculated with Xanthomonas species NCPPB 1131 and 1132 originally isolated from banana plants also remained healthy. All 112 banana plants had their older leaves yellowing with scorched appearance throughout the trial most likely due to natural ageing rather than disease infection.



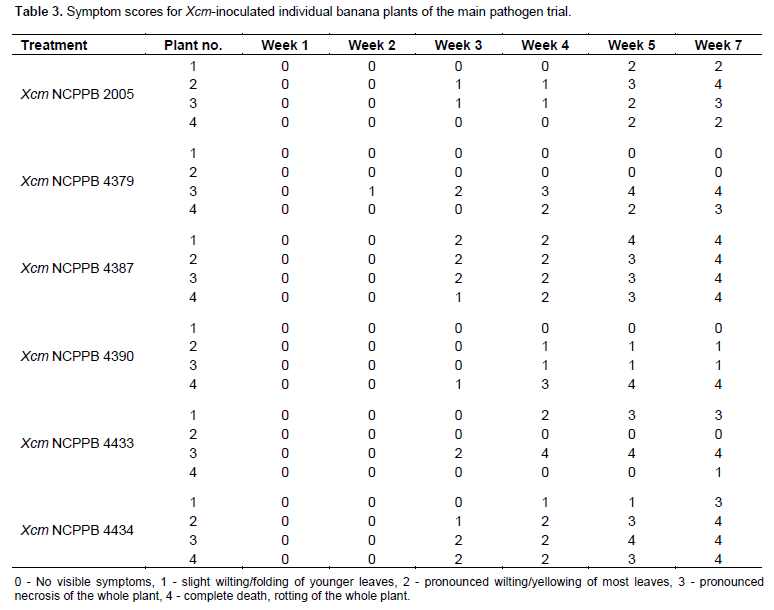
The 20 plants inoculated with Xcm showed severe typical symptoms of BXW 3 weeks after inoculation. The disease affected the younger leaves first, beginning with dull green coloring of the lamina, folding of the two halves of the midrib touching each other, yellowing of the leaves, reddish brown streaks on the leaf, eventually all the leaves wilted and entire plant rotted away. Most of the Xcm inoculated plants were dead by the 7th week of the trial. Re-isolation of the Xcm from most Xcm-inoculated plants was successful but rather difficult from plants that had already died as it was apparent that other microbes had already invaded the plants.
Other pathovars such as X. axonopodis pv. vasculorum, X. campestris pv. pelagonii, X. campestris pv. campestris and X.campestris pv. vesicatoria did not cause disease in banana but were successfully-re- isolated. The non-Xanthomonas strain Paenibacillus larvae were not successfully re-isolated (Table 4).
.png)
Maize large scale pathogenicity trial
The 16 control plants; eight inoculated with sterile water and eight left untreated remained healthy throughout the trial. Out of the 24 plants inoculated with Xanthomonas vasicola pathovars (X.vasicola pv. vasculorum and X.vasicola pv. holcicola), 4 maize plants inoculated with Xvh NCPPB 3129 remained healthy (Table 5). The other 20 plants and the eight inoculated with Xanthomonas axonopodis pv. vasculorum displayed symptoms consistent with the known pathogen profiles of Xvv, Xvh and Xav in the pilot studies. Symptoms appeared 6 days after inoculation, these included; yellow, brown, white or water soaked streaks as well as brown lesions on the leaves, usually beginning in the center of the leaf. The most severe symptoms were deformation of the plant and retarded growth by X. vasicola pv. vasculorum and X. vasicola pv. holcicola respectively (Figure 2a-b). These symptoms separated Xvv and Xvh The 24 plants that had been inoculated with X. campestris pv. musacearum did not exhibit any of the symptoms throughout the trial, suggesting that Xcm did not affect maize under these conditions and re-isolation from a few of these plants was successful even five weeks after inoculation. Plants inoculated with other common plant pathovars; Xanthomonas campestris pv. campestris, Xanthomonas campestris pv. vesicatoria and X. campestris pv. perlargonii did not show any signs of disease (Table 6).

.png)
However by the 5th week of the trial, most of the diseased plants seemed healthier than they had been 1 week after inoculation; the affected leaves were fewer or either drying or falling off. The leaf symptoms such as the streaks and lesions were not exhibited on leaves that had inoculation sites thereby confirming the symptoms were not as a result of HR. Some of the plants inoculated with Xvv and Xav that had earlier had their leaves with lesions and streaks (one to two weeks after inoculation) later became healthy, showing no more symptoms on any other leaves or on leaves that had the inoculation sites or any other parts of the plant by the 5th week. Four out of the eight plants inoculated with Xvh (these were inoculated with Xvh NCPPB 3126) remained healthy and throughout the trial (asymptomatic) and did not exhibit any visible symptoms even one week after inoculation, and re-isolation of the Xvh bacteria from these plants was successful.
Sugarcane large scale pathogenicity trial
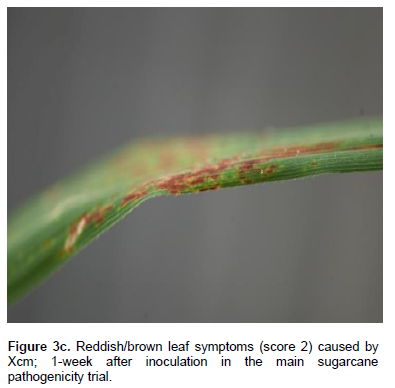
All control plants (inoculated with sterile water and those left untreated) remained healthy throughout the trial. Xvv, Xvh, Xcm, and Xav caused foliar symptoms such as reddish-brown streaks and white streak spots (Figure 3a-d, Table 7). Xcm also caused white and yellow streaks on the leaves). Xav also caused white streaks on the leaves. These symptoms appeared 1 week after inoculation. Out of the 6 plants inoculated with Xvh, 3 of them (these had been inoculated with Xvh NCPPB 3126) remained healthy. Out of the six plants inoculated with Xav, 2 of them remained health. Out of the 12 plants inoculated with Xvv, 2 of them remained health. Out of 21 plants inoculated with Xcm, 7 remained healthy.


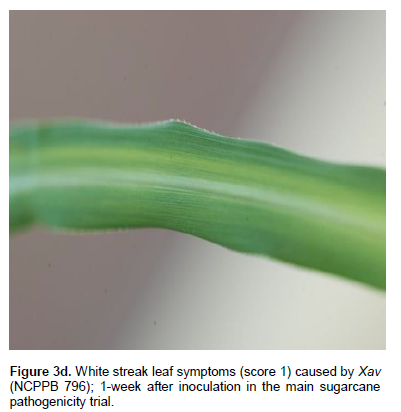

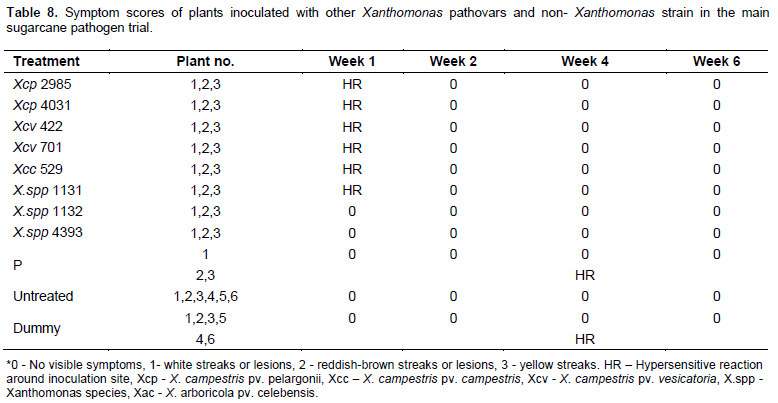
Common plant pathogenic Xanthomonas pathovars; X. campestris pv. pelargonii, X.campestris pv. campestris, X. campestris pv. vesicatoria and the Xanthomonas species (NCPPB 1131) originally isolated from banana caused white lesions and patches around the inoculation site on the leaves suspected to be more of a hypersensitive reaction (HR) to the pathogens in sugarcane, one week after inoculation (Figure 3e and Table 8). The other common plant Xanthomonas pathovar; X.arboricola pv. celebensis as well as the non Xanthomonas strain Paenibacillus larvae did not affect sugarcane.
Viability of the bacterial inoculum
The 100 µl of bacteria inoculum that was plated before each inoculation of a plant did grow within 48 h at 28°C incubation, confirming that the inoculum was viable.
DISCUSSION
This study has been able to provide substantial data on the comparative pathogenicity of the Xanthomonas vasicola pathovars and Xcm (Table 2). This study alsowhile Xvh naturally affects sorghum (Navi et al., 2002). Even though the two pathogens caused similar symptoms such as lesions or streaks on leaves of maize and sugarcane, they caused distinct symptoms in maize: Xvv causes deformation of the plant while Xvh causes stunted growth. This then separates them as two different pathovars or warrants different pathovar designations based on the differences in symptomology on the same host (Young et al., 2001). Differences in the draft genomes of Xvv isolates by Wasukira et al. (2014) may also attempt to explain the possible cause of the difference in symptomology by Xvv and Xvh. Wasukira’s study revealed that an Xvv isolate from maize had lost the virulence factor xopAF which was present in five other Xvv isolates from sugarcane. As Xvv and Xvh are also closely related, Xvh may be lacking this virulence factor xopAF.
Unfortunately we were not able to conduct pathogenicity tests on sorghum, to further confirm this symptom difference between Xvv and Xvh. The pilot trial for sorghum was unsuccessful and this could probably be attributed to either the age of the seedlings or it could be that artificial inoculation of sorghum is usually unsuccess-ful (Navi et al., 2002). However, some of the maize plants that had been affected one week after inoculation later recovered 5 weeks after inoculation and showed no more signs of disease suggesting that maize may be less susceptible to both pathogens.
A previous study has shown Xanthomonas campestris pv. musacearum (Xcm) to be able to cause disease in maize (Aritua et al., 2008), however this has been weakly supported in our study. Possible reasons as to why maize plants in the main trial that had been inoculated with X. campestris pv. musacearum remained healthy; the maize sub-species used in the two studies may have been different, the growth stage of the maize seedlings (in both studies), the greenhouse conditions of the two trials were different. However, based on the three possible reasons, these conditions may have caused a latency period for Xcm in maize. Latency period can be defined as period before the pathogen induces symptoms (Verhoeff et al., 1974) or when the symptoms appear due to the changes in the environmental and nutritional stage of maturity in the host or pathogen (Agrios, 1988).This suggests that most likely the conditions of the green house or the maize varieties used in both studies were critical factors.
The successful re-isolation of Xcm from the healthy maize plants also further suggests that maize can be a reservoir for Xcm strains. This is highly significant and should contribute to the control methods currently used for Xanthomonas wilt of bananas especially since maize are among the crops that usually intercropped with bananas in Uganda.
Pathogenicity of Xvv, Xvh, and Xcm on sugarcane
Symptoms caused by both X. vasicola pathovars and Xcm on sugarcane were similar, that is reddish-brown streaks or lesions on the leaves which may be explained by the genetic closeness the three strains share (Parkinson et al., 2009). Though one strain of Xvh NCPPB 3126 used in all the main pathogenicity trials was successfully re-isolated, it did not cause any symptoms in both maize and sugarcane. This strain may have lost its ability to cause disease after being kept in -80ËšC and may just have been surviving as an asymptomatic endophyte.
Pathogenicity of Xvv, Xvh, and Xcm on banana
The study has also shown the X. vasicola pathovars to be non-pathogenic on banana but the strains still able to survive within the plant asymptomatically. This suggests banana can be a host to other Xanthomonas strains (apart from Xcm) including Xvv and Xvh strains. X. axonopodis pv. vasculorum (Xav) is a pathovar that though does not fall within the X. vasicola species, causes similar symptoms apart from deformation and stunted growth. This separates Xav from the X. vasicola pathovars. The study demonstrates that Xav is patho-genic on maize and sugarcane but not banana. This suggests that Xav may share a few genetic similarities with the X. vasicola pathovars in terms of its ability to cause disease both in maize and sugarcane.
Xcm was also clearly pathogenic on sugarcane and the symptoms on sugarcane were also very similar to those caused by the X. vasicola pathovars. Only Xcm was distinctly pathogenic on banana while the X. vasicola pathovars did not affect banana. According to Studholme et al., 2010, the draft genomes of Xcm and Xvv are significantly similar; however differences in Type III secretion system (T3SS) effectors may explain their differences in host adaptations. Xcm encodes two predicted YopJ-like C55 cysteine proteases that are absent from Xvv. Previous studies (Aritua et al., 2008, Parkinson et al., 2009) have shown Xcm to fall within the X. vasicola species and the pathogenicity trials have revealed that there are clearly interspecific pathovar differences within the species. It is still unclear as why some Xcm-inoculated banana plants remained healthy; however such a phenomenon is not new. It has been shown that latency infection of BXW does normally occur in bananas (Ocimati et al., 2015) and is the cause of recurring BXW incidences.
Other Xanthomonas strains on banana, maize, and sugarcane
Re-isolation of other Xanthomonas bacteria from banana was successful but not from maize or sugarcane, suggesting that banana can be reservoir to common Xanthomonas plant pathogens; X. campestris pv. campestris, X.campestris pv. perlargonii, X.campestris pv. vesicatoria and the X. vasicola pathovars. Xanthomonas species NCPPB 1131 and 1132 originally isolated from bananas (Studholme et al., 2011) were shown to be non-pathogenic on banana.
CONCLUSION
The pathogenicity trials have shown that the current X. vasicola species (Xvv and Xvh) can cause disease in similar hosts, and there are differences in symptomatology. The genome studies of Xvh may explain the differences in symptomology caused by Xvv and Xvh. Xcm causes disease in the similar hosts (maize and sugarcane), causing similar foliar symptoms which further supports the genetic closeness Xvv, Xvh and Xcm share. Only Xcm causes disease in banana. This data also provides additional information on the pathogenicity of the X. vasicola pathovars and should also further support the proposal to rename Xcm to Xanthomonas vasicola pv. musacearum.
CONFLICT OF INTEREST
The authors have not declared any conflict of interest.
ACKNOWLEDGEMENTS
This study was supported by the Mcknight Foundation, Bioversity International and Uganda government through the National Banana Research Program. The authors wish to thank the staff at FERA (The Food and Environment Research Agency) for their invaluable support in providing the plants used in the trial and proper maintenance of the glass house conditions.
ABBREVIATIONS
BXW, Banana Xanthomonas Wilt; NCPPB, National Collection of Plant Pathogenic Bacteria; pv., pathovar; Xcm, Xanthomonas campestris pv. Musacearum; Xvh, Xanthomonas vasicola pv. Holcicola; Xvv, Xanthomonasvasicola pv. Vasculorum.
REFERENCES
| Agrios GN (1988). Plant Pathology. 3rd ed. Academic Press, New York.803pp. | ||||
| Aritua V, Parkinson N, Thwaites R (2008). Characterization of the Xanthomonas sp. causing wilt of enset and banana and its proposed reclassification as a strain of X. vasicola. Plant Pathol. 57:170-177. | ||||
| Aritua V, Parkinson N, Thwaites R, Tushemereirwe W, Smith J (2008). Molecular epidemiology of Xanthomonas campestris pv. musacearum, the causal agent of Xanthomonas wilt of Banana and Enset. Acta Horticulturae, 828:219-226. | ||||
|
Biruma M, Michael P, Leena T, Guy B, Steffen A, Maina M, Ranajit B, Perez M, Sadik K, Moses NL (2007). Banana Xanthomonas wilt: a review of the disease, management strategies, and future research directions. Afr. J. Biotechnol. 6(8):953-962. |
||||
|
Carter BA, Reeder R, Mgenzi SR, Kinyua ZM, Mbaka JN, Doyle K, Nakato V, Mwangi M, Beed F, Aritua V, Lewis Ivey ML, Miller SA and Smith JJ (2010). Identification of Xanthomonas vasicola (formerly X. campestris pv. musacearum), causative organism of Xanthomonas wilt in Tanzania, Kenya and Burundi. Plant Pathol. 59(2):403. Crossref |
||||
|
Dookun A, Stead DE, Autrey LJ (2000). Variation among the strains of Xanthomonas campestris pv. vasculorum from Mauristius and other countries based on fatty acid analysis. Syst. Appl. Microbiol. 23(1):148-155. Crossref |
||||
| Garrity GM (2005). "Genus Xanthomonas." In Bergey's Manual of Systematic Bacteriology; the Proteobacteria 2nd edition. 2:87. | ||||
|
Hodgetts J, Karamura G, Johnson G, Hall J, Perkins K, Beed F, Nakato V, Grant M, Studholme DJ, Boonham N, Smith J (2014). Development of a lateral flow device for in-field detection and evaluation of PCR based diagnostic methods for Xanthomonas campestris pathovar musacearum, the causal agent of banana Xanthomonas wilt. Plant Pathol. 64(3):559-567. Crossref |
||||
| Navi SS, Bandyopadhyay R, Devi KT, Reddy DVR (2002). Bacterial Leaf Streak on Sorghum – a New Report from India. 43:61-63. | ||||
|
Ndungo V, Eden-Green S, Blomme G, Crozier J, Smith JJ (2006). "Presence of Banana Xanthomonas wilt (Xanthomonas campestris pv. musacearum) in the Democratic Republic of Congo (DRC)." Plant Pathol. 55(2):294. Crossref |
||||
|
Ocimati W, Nakato GV, Fiaboe KM, Beed F, Blomme G (2015). Incomplete systemic movement of Xanthomonas campestris pv. musacearum and the occurrence of latent infections in Xanthomonas wilt-infected banana mats. Plant Pathol. 64:81-90. Crossref |
||||
|
Parkinson N, Cowie C, Heeney J, Stead D (2009). Phylogenetic structure of Xanthomonas determined by comparison of gyrB sequences. Int. J. Syst. Evol. Microbiol. 59:264-274. Crossref |
||||
| Sinclair BJ, Dhingra DO (1995). Basic Plant Pathology Methods, 2nd edition, CBC Press. | ||||
|
Tripathi L, Mwaka H, Tripathi JN, Wilberforce T (2010). Expression of sweet pepper Hrap gene in banana enhances resistance to Xanthomonas campestris pv. musacearum. Mol. Pathol. 11(6):721-731. Crossref |
||||
| Yirgou D, Bradbury JF (1968). Bacterial wilt of enset (Ensete ventricosum) incited by Xanthomonas musacearum sp. Phytopathol. 58:111-112. | ||||
|
Verhoeff K (1974). Latent infections by fungi. Ann. Rev. Phytopathol. 12:99-107. Crossref |
||||
|
Wasukira A, Coulter M, Al-Sowayeh N, Thwaites R, Paszkiewicz K, Kubiriba J, Smith J, Grant M, Studholme, DJ (2014). Genome Sequencing of Xanthomonas vasicola Pathovar vasculorum Reveals Variation in Plasmids and Genes Encoding Lipopolysaccharide Synthesis, Type-IV Pilus and Type-III Secretion Effectors. Pathog. 3:211-237. Crossref |
||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


