ABSTRACT
Maize (Zea mays L.) is the most important cereal crop on which many smallholder farmers in sub-Sahara Africa (SSA) depend on as a sole source of calories, proteins, and vitamins. However, conventional maize is deficient in two essential amino acids, lysine, and tryptophan. Thereafter, Quality Protein Maize (QPM) developed to combat protein malnutrition. In SSA low soil nitrogen is also the most limiting factor in maize production and productivity. To determine the combining ability of QPM inbred lines for grain yield and other agronomic traits under low and optimum N environments, 121 genotypes generated using 11 QPM inbred lines in a complete diallel mating design with 5 checks were evaluated under both low and optimum N environments at 3 locations in the 2018 cropping season. Significant differences were observed among the genotypes for major characters under both low and optimum N environments. Under low and optimum N environments, non-additive gene actions were more important than additive gene action for grain yield, number of ears per plant, plant and ear height, ear length and diameter. Under low N environments, more contributions of reciprocal effects than GCA effects were observed for number of ears per plant, plant aspect, ear diameter. Parental lines TL156579, TL156583, and TL148288 were considered as a good general combiner for grain yield under low N environments. Parental line TL156612 showed higher positive GCA effects when used as female while parental line TL156612 showed higher negative GCA effects when used as a male parent for grain yield under low N environments.
Key words: Complete diallel, general combining ability, low soil nitrogen, lysine, optimum soil nitrogen, quality Protein maize, reciprocal effect, tryptophan.
Maize is one of the most important food crops and is widely grown all over the world on which the majority of developing countries depend on as a sole source of calories, proteins, vitamins and minerals Babu and Prasanna, 2014; Ranum et al., 2014; Badu-Apraku et al., 2015b; Farfan et al., 2015). However, in convemtional maize (normal maize), the concentration of lysine and tryptophan are low, which are more important for both humans and monogastric animals. In countries where maize is consumed as the primary or sole source of protein, malnutrition is common due to the lack of essential amino acids in maize kernel endosperm lysine and tryptophan.
Nitrogen (N) is an important nutrient for maize yield. It is a constituent of amino acids, proteins, hormones, and chlorophyll. Reduced seed production, leaf chlorosis, and reduced plant root branching occur due to a low soil N supply. Maize yield is more dependent on the availability of soil N, which is one of the major limiting factors in sub-Saharan Africa (SSA). Even if better yield of maize is dependent on the amount of N fertilizer supplied to the crop, many developing countries' farmers apply N fertilizer at sub-optimal levels due to the high cost of fertilizer, which leads to low yield. In addition to the high cost of N fertilizer, maize can only uptake 30% of applied N; and then, the rest 70% of the N applied to maize is lost due to different mechanisms like leaching, de-nitrification and surface runoff from the soil, which causes soil acidification and affect both plant and animal biodiversity (Erisman et al., 2013). To address both economic and environmental problems related to N fertilizer, development of genotypes that yield better under low N supply by improving N-use efficiency (NUE) of maize genotypes is the most important strategy for sustainable agriculture (Gelli et al., 2014). NUE is the capacity of the crop to produce biomass and grain yield per unit of available N in the soil (Moll et al., 1982); and it is the product of N-uptake efficiency and N-utilization efficiency (Banzinger et al., 2000). Improving NUE of a crop through breeding methods can improve crop yield.
In maize breeding strategy, information on combining ability, heterosis, and heterotic groups are the most important factors for the development of high-yielding maize hybrids. A heterotic group is a group of related or unrelated genotypes from the same or different populations, which display similar combining ability and heterotic response when crossed with genotypes from other genetically distinct germplasm groups. Analysis of combining ability for inbred lines indicates the type of gene action controlling quantitative characters, which are used for genetic diversity analysis, inbred selection, heterotic grouping, estimation of heterosis, and hybrid development (Fan et al., 2009; Hallauer et al., 2010). The advantage of grouping germplasms into divergent heterotic groups is advantageous because of higher mean heterosis and hybrid performance and a reduced specific combining ability (SCA) variance and a lower atio of SCA to GCA variance. Thus, early testing becomes more effective and superior hybrids can be identified and selected mainly based on their prediction from GCA effects (Reif et al., 2005). The most effective and direct way to solve the maize yield gap is to breed varieties with high yielding potential and wide adaptability. About 52.9% of maize yield increment was attributed to development of new varieties, and the rate of improvement was 89.1 kg ha-1 per year (Wang et al., 2009), which is attributed to heterosis.
Breeding strategies for maize under stress environments have been influenced by the breeding strategy for maize under non-stress environments, in which selection under high yielding environments may not be effective selection to increases yield under low yielding environments. This is due to the finding that plant traits that are less important under non-stressed environments, become more important for yield under stressed environments (Banzinger et al., 2000). Thus, the selection of germplasm under both low and optimum N environments can increase the chance to develop low N tolerant maize hybrids. Some organizations have developed QPM inbred lines with tolerances to low N; however, their adoptions in Sub-Saharan Africa countries were very low (Badaway, 2013).
In Ethiopia, maize is ranked first in terms of total production and second in total area coverage; which accounts for 47.84% out of 73.35% of the land allocated to all cereals in the country (CSA, 2019). Maize can be grown in different agro-ecologies (highland, mid-altitude, and lowland) of the country. In Ethiopia, low maize yield production and productivity is associated with several limiting factors including, lack of high yielding and stable improved genotypes, drought, reduction in soil N fertility, inappropriate agronomic practices, low adoption of improved agricultural technologies including varieties by farming communities, foliar diseases, weeds, and high cost of inorganic fertilizer. Therefore, the objective of this study was to estimate the combining ability effects of QPM inbred lines under both low and optimum N environments for grain yield and other desirable traits.
Plant materials
Eleven inbred lines were selected based on their tolerance to a low N environment and high tryptophan content from a panel of 74 QPM inbred lines obtained from the maize breeding program at CIMMYT-Zimbabwe; and these were used in this study (Table 1). A complete diallel mating design was made among these 11 QPM inbred lines at Haramaya University research site (Raare), to generate 121 genotypes. The 121 genotypes with two non-QPM hybrid maize (BH547 and BH546) and three QPM hybrid (BHQPY548, BHQPY545 and MHQ138) checks were used in this study.
Experimental sites and design
The field experiments were conducted at three research sites (Raare, Fedis and Dire-Dawa) of Haramaya University, Ethiopia. The university is located in the Eastern part of the country, which represents the mid-altitude maize growing ecology of the country. The Raare research site lies between 9º40' N latitude and 42º03' E longitude at an altitude of 2020 m.a.s.l with an annual rainfall of 820 mm and average maximum and minimum temperature of 24.4ºC and 9.8ºC, respectively. Fedis lies between 8º49' N latitude and 42º00' E longitude at an altitude of 2,118 m.a.s.l with an annual rainfall of 750 mm and average maximum and minimum temperature of 30.4ºC and 13.6ºC, respectively. Dire-Dawa research site lies between 9º97' N latitude and 42º53' E longitude at an altitude of 1180 m.a.s.l with an annual rainfall of 637 mm and average maximum and minimum temperature of 33.0ºC and 18.9ºC, respectively. One hundred and twenty one hybrids generated from the complete diallel cross with 5 hybrid checks (126 genotypes) were arranged in 18 x 7 alpha lattice (0, 1) design with 2 replications in each location under both low N (30 kg ha-1) and optimum N (100 kg N ha-1) environments.

Depletion of N in experimental sites and soil analysis
Depletion of nitrogen in the soil was done by planting high population densities of maize plants without applying nitrogen fertilizer for two years and removing all biomass after harvest from the fields. The soil chemical properties of the sites were determined at two soil sample depths (0-30 cm and 30-60 cm) before planting at Haramaya university soil laboratory. From soil laboratory analysis results, the total N content at Raare was 0.070% and 0.056%, 0.098% and 0.084% at Fedis and 0.112% and 0.098% at Dire-Dawa for 0-30 cm and 30-60 cm soil depth, respectively; which were considered as low soil N at all locations.
Low soil N management
The field experiment was divided into two blocks (Low and optimum N blocks). Two seeds per hill on a one-row length of 4 m at 30 cm distance between plants and 75 cm between rows were planted. The plants were thinned to one plant per hill to give 44,444 plant populations per hectare. The recommended rate of N fertilizer for maize (100 kg N ha-1) was applied in the form of Urea for optimum N environment and 30 kg N ha-1 for low N environment. Low N environment of 30 kg N ha-1 was used by considering sub-Saharan Africa countries’ farmers apply 20-30% fertilizer below recommended rate because of the high cost of fertilizer and low access to fertilizer for their crops. In addition to N fertilizer, both experiments were supplied with 100 kg P2O5 ha-1 in the form of tricalcium phosphate. Split N fertilizer application was done on each plot for both low and optimum N environments. Under a low N environment, a mixture of 18 kg N ha-1 and 46 kg P2O5 ha-1 were applied at planting and 12 kg N ha-1, 45 days after planting. Under optimum N environment, a mixture of 50 kg N ha-1 in the form of Urea and 46 kg P2O5 ha-1 were applied at planting, and 50 kg N ha-1 was applied in the form of Urea 45 days after planting. Weeds were removed by hand weeding at each location.
Data collection
Based on the International Board for Plant Genetic Resources (IBPGR) descriptor list (IBPGR, 1991) phenotypic characters were measured from five randomly selected plants except for days to silking and anthesis, anthesis-silking interval, plant aspect, ear aspect, stay green characteristics which were recorded on a pilot basis (Table 2).
Data analysis
Analysis of variance per environment was performed on plot means for grain yield and other agronomic traits using PROC GLM procedure of SAS software, version 9.4 (SAS Institute, 2012).
Combining ability analysis
Analysis of variance for a complete diallel cross excluding checks was done using AGD-R (Analysis of Genetic Designs with R) version 5.0 (Rodríguez et al., 2018). Griffing’s method I (parents, F1s, and F1's reciprocals were included) and model I (fixed effect) of diallel analysis was used for computing GCA effects of the parents, SCA and reciprocal effects of the crosses. Analysis of variance was done for the individual environment, and combined analyses of variance over low N, Optimum N and across all N environments were done for those traits showed significant genotypes mean squares in an individual environment. The mean squares for ENV and crosses were tested against the mean squares for ENV x crosses as mean square of error, while ENV x crosses interactions mean squares were tested against pooled error. The significance of GCA, SCA and reciprocal source of variations were tested against F-tests while the significance of GCA, SCA and reciprocal effects were tested against t-test, with the standard error of GCA, SCA and reciprocal effects (Dabholkar, 1999; Griffing, 1956). The linear model for combining ability analysis for Griffing's method-I of the complete diallel cross for a single environment was computed as (Griffing, 1956):

The relative importance of combining ability
The ratio of combining ability variance components determines the type of gene action involved in the expression of traits and allows inferences about the optimum allocation of resources in hybrid breeding. The closer this ratio is to one, the greater the prediction of GCA alone, whereas a ratio with a value less than one shows SCA action. However, because in many cases only a few parents are used in crosses, the magnitude of GCA, SCA and reciprocal effects evaluated using the ratio of their sum of squares to the total sum of squares for crosses where they were involved were used to determine the importance of combining ability.
Combined analysis of variances and importance of combining ability effects of inbred lines under low, optimum, and across N environments
Under combined low N environments, environmental effects were significant for all traits except the plant aspect (Table 3). Mean squares for GCA and SCA were highly significant for all traits and E x crosses and E x GCA interactions were highly significant for all traits; while reciprocal and non-maternal effects were significant for all traits except the ear aspect. Mean squares for maternal effects were highly significant for grain yield, days to anthesis and silking, plant height, ear height, ear length and ear diameter. E x SCA and E x reciprocal interactions were highly significant for all traits except plant aspect and ear aspect. E x maternal interactions were significant for grain yield, days to anthesis, number of ears per plant, plant height, ear height and stay green characteristics. The mean square of E x non-maternal interaction was significant for all traits except days to anthesis and plant aspects.
Under combined low N environments, the percent of sum of squares for GCA ranged from 8.69% for ear length to 46.95% for days to silking; while percent of sum of squares for SCA ranged from 39.55% for days to silking to 73.54% for ear length. Percent of sum of squares for reciprocal effects ranged from 5.11% for ear height to 34.34% for number of ears per plant (Table 4). High GCA sum of squares accounted for days to anthesis (45.95%), days to silking (46.95%) and stay green characteristics (41.61%); while low GCA sum of squares were detected for number of ears per plant (13.02%), plant aspect (12.5%), ear length (8.65%) and ear diameter (10.88%) of a total variation among crosses. A higher SCA sum of squares than for the GCA sum of squares were recorded for all traits; while a lower reciprocal sum of squares than for the SCA sum of squares were recorded for all traits under low N environments.
Under optimum N environments, mean squares for environments were highly significant for all traits (Table 4). Variances of GCA, SCA, E x GCA, and E x SCA interactions were significant for all traits; but, E x maternal interactions were only significant for grain yield, plant height, and ear height. Ex non-maternal interaction was only significant for grain yield, number of ears per plant, plant height, and ear height. Mean squares for reciprocal effects were significant for all traits, except days to anthesis and silking; while E x reciprocal interactions were highly significant for grain yield, number of ears per plant, plant height and ear height. Maternal and Non-Maternal mean squares were highly significant for all traits, except days to anthesis and silking and ear aspect.
GCA sum of squares under optimum N environments ranged from 11.87% for plant aspect to 54.92% for days to silking; while sum of squares for SCA ranged from 36.86% for days to silking to 75.60% for plant aspect. Percent of sum of squares for reciprocal effects ranged from 8.22% for days to silking to 24.79% for number of ears per plant (Table 4). High GCA sum of squares accounted for days to anthesis (53.99%), days to silking (54.92%) and ear height (40.68%) of a total variation among crosses. A smaller SCA sum of squares than for the GCA sum of squares were accounted for days to antheis and silking of the total variation among crosses. A greater reciprocal sum of squares than for the GCA sum of squares were accounted for number of ears per plant and plant aspect of the total variation among the crosses under optimum N environments.
Combined mean squares for environmental effects were significant for all traits across all environments. Highly significant differences were also found among crosses for all measured traits (Table 5). Mean squares of GCA, SCA, reciprocal, non-maternal, and E x Crosses, E x GCA, and E x SCA interactions were highly significant for all traits. E x reciprocal and E x maternal interaction mean squares were significant for all traits except for days to anthesis and silking and ear aspect; while E x non-maternal interaction was highly significant for all traits except days to anthesis, days to silking and plant aspect.
Across all N environments, the GCA sum of squares ranged from 12.78% for plant aspect to 56.22% for days to silking; while SCA sum of squares ranged from 34.19% for days to silking to 75.97% for plant aspect. The reciprocal sum of squares ranged from 5.15% for grain yield to 24.60% for number of ears per plant (Table 5). A higher contribution of GCA effects was found for days to silking (56.22%), and lower contribution for plant aspect (12.78%); while there was a high contribution of SCA effects for ear length (73.90%), with a low contribution for days to anthesis (35.48%). A greater reciprocal sum of squares than GCA sum of squares was accounted for number of ears per plant across all N environments.
Combining ability effects
General combining ability and maternal effects
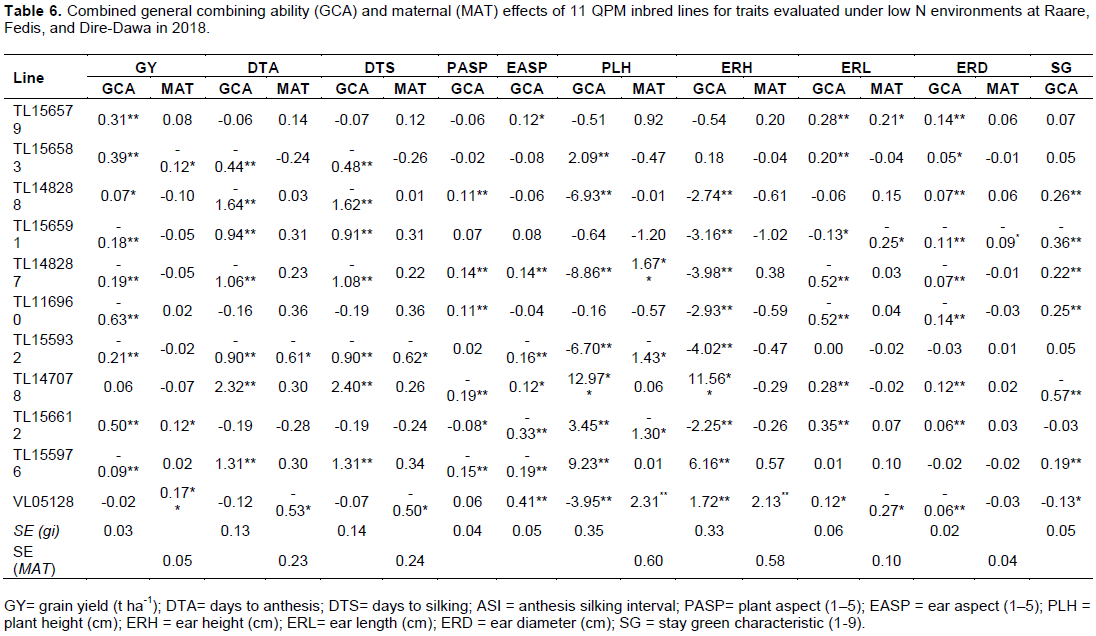
From combined analysis under a low N environment, inbred lines TL156579, TL156583, TL148288, and TL156612 had significant positive GCA effects for grain yield; while inbred lines TL156612 and VL05128 had significant positive MAT effects for grain yield (Table 6). TL148288, TL148287, and TL116960 inbred lines had positive and significant GCA effects for plant aspects. Positive and significant GCA effects were found in TL156579, TL147078, and VL05128 inbred lines for the ear aspect. Inbred lines TL156583, TL147078, and TL156612 had significant positive GCA effects for plant height, ear length, and ear diameter. Significant positive GCA and MAT effects were found for ear height in the VL05128 inbred line. GCA effects were found to be significant and positive in TL156579, TL156583, TL147078 and TL156612 inbred lines for ear length and diameter. Significant positive GCA effects with significant negative MAT effects were found in the VL05128 inbred line for ear length. Inbred lines TL148288, TL148287, TL155976, and TL116960 had significant positive GCA effects for stay green (Table 6).
Under optimum N environments for combined analysis, inbred lines TL156579, TL148288, TL147078, and TL156612 had positive significant GCA effects for grain yield. Inbred lines TL156583 and TL155976 had both positive and significant GCA and MAT effects for grain yield; while inbred lines TL156591 and VL05128 had negative significant GCA and MAT effects for grain yield. Positive and significant GCA effects, with negative significant MAT effects, were observed in TL147078 inbred lines for grain yield. The TL156583 inbred line had a positive and significant GCA effect for number of ears per plant. Two inbred lines (TL148287 and VL05128) had significant and positive GCA effects for plant and ear aspects. Inbred lines TL148287, TL155976 and VL05128 had significant positive MAT effects for plant height. GCA and MATeffects were found to have significant positive otucomes in the VL05128 inbred line for ear height.
Significant positive GCA and MAT effects were observed in the TL156579 inbred line for ear length. Inbred lines TL156579, TL155976, and VL05128 had significant positive GCA effects for ear diameter (Table 7).
Significant and positive GCA effects were found for grain yield in TL156579, TL156583, TL148288, TL147078, and TL156612 inbred lines across all N environments (Table 8). A significant positive GCA effect with a significant negative MAT effect was found in TL147078 for grain yield. A significant negative GCA effect with a significant positive MAT effect was observed in TL155976 inbred line for grain yield. Inbred lines TL156583 and TL156612 had significant positive GCA effects. For plant and ear aspects, inbred lines TL148287 and VL05128 had significant positive GCA effects.
Significant negative GCA effects with significant positive MAT effects were observed in TL148287 and VL05128 inbred lines, but positive and significant GCA effects with negative and significant MAT effects were observed in TL156612 inbred lines for plant height. Positive and significant GCA and MAT effects were found in the TL156579 inbred line for ear length. Lines TL156579 and TL147078 had significant positive GCA effects for ear diameter (Table 8).
Under both low and optimum N environments, significant differences were found among genotypes for grain yield, days to anthesis, days to silking, plant aspect, ear aspect, plant height, ear height and length and eaer diameter. This indicates genetic variability among genotypes, which creates a great opportunity for a maize breeder to improve genotypes for yield and other agronomic traits under both soil environments. Different authors also reported similar results for QPM hybrids both under low and optimum N environments (Badu-Apraku et al., 2015b; Wegary et al., 2014).
Significant differences among genotypes for traits of interest at a specific location under specific N environments indicate that each environment was unique for each genotype; and each genotype responded differentially under low and optimum N environments. Similar findings were reported by Badu-Apraku et al. (2010)who reported the effects of stress environments on the performance of tropical early-maturing maize cultivars in multiple stress environments.
Single cross hybrids VL05128 x TL156612 (4.89 t ha-1) and TL156612 x TL155932 (4.74 t ha-1) were selected as high yielding hybrids which out-yielded by 16.76% and 14.14%, respectively, than the best check (4.07 t ha-1) under low N environments due to better tolerance effects of the genotypes to stress environments than the checks. Under low N environments, a 20% grain yield increment over local commercial check was reported by Worku et al. (2001)for normal maize varieties. Overall, mean percentage of relative grain yield reduction for hybrids was 32% under low N environments as compared to optimum environments, which are within a range of 25% to35% yield reduction as recommended by Banzinger et al. (2000)to be selected as resistant/ tolerant cultivars under low N environments. This level of yield reduction is closer to that of Below (1997) and Presterl et al. (2003), who reported yield reduction under low N stress of 35% and 37%, respectively, for normal maize hybrids. Yield reduction of 42% for tropical early maturing cultivars was reported by Badu-Apraku et al. (2010)and 40% of grain yield reduction for QPM hybrids by Badu-Apraku et al. (2015b)under low N environments. Different authors have reported different percentages of relative yield reduction under low N environments because of differences in the severity of low N environments and the germplasm used for the study.
From the analysis of genetic designs, GCA effects provide an estimation of additive gene action, while SCA effect provides non-additive gene action (Baker, 1978); and reciprocal effects provide an estimation of additive gene action through maternal (MAT) effects and non-additive gene action through non-maternal (NMAT) effects (Fan et al., 2013; Mahgoub, 2011). Significant differences were found among QPM hybrids for almost all traits both under low and optimum N environments, indicating the available genetic variability among the hybrids under low and optimum N environments, which would allow good progress to the selection of the hybrids under the target environments. Similar results were reported by different authors for QPM and non-QPM hybrids for grain yield, days to anthesis and silking, plant and ear height, plant and ear aspect under low and optimum N environments (Badu-Apraku et al., 2015a; Njeri et al., 2017; Wegary et al., 2014).
Under combined low, optimum, and across all N environments, significant mean squares of GCA, SCA and reciprocal effects were found for grain yield and other agronomic traits indicating the importance of additive, non-additive gene action and maternal cytoplasmic genes under these target environments. This implies that in addition to additive and non-additive gene actions, the inheritance of the traits was controlled by maternal effects that created differences in the performance of grain yield and other traits between F1 hybrids and their F1 reciprocals, which allow selection of superior hybrid under the target environments. Fan et al. (2013) and Mahgoub (2011)reported the effects of reciprocal crosses for maize grain yield under non-stress environments.
Highly significant E x crosses interactions were observed for all traits under low, optimum, and across all environments indicates that crosses reacted differently to different testing environments; thus, the breeder should test the hybrids in a wide range of environments for the selection of best performing and stable crosses. This finding is consistent with the result reported by Badu-Apraku et al. (2015a)and Wegary et al. (2014)for QPM hybrids under multiple stress environments.
Significant mean squares for E x GCA interaction effects were observed for grain yield and all other agronomic traits under combined low, optimum, and across all N environments; which indicated that GCA effects of the parental lines exhibited variations under the considered environments of this study. Similar results were reported by different authors (Badu-Apraku et al., 2015; Badu-Apraku et al., 2010, 2017; Njeri et al., 2017; Wegary et al., 2014). This finding also points out that, for the development of high yielding and stable hybrids across a range of environments for commercial purposes, there is the need to test parental lines for their GCA and SCA effects across a range of environments, and/or different parental lines need to be selected for each environment for hybrid development. Significant mean squares of E x SCA interaction effects were observed for grain yield and other measured traits under combined low, optimum and across all N environments, except for plant and ear aspects under combined low N environments. This indicates that except for plant and ear aspects under combined low N environments, all traits were significantly different across environments of this study. This finding is in disagreement with the finding of Wegary et al. (2014), who reported non-significant E x SCA interaction effects of QPM hybrids for grain yield and most of the other agronomic traits evaluated under drought, low and optimum N environments. However, this finding is in agreement with the finding of Badu-Apraku et al. (2015a), who reported significant E x SCA interaction effects of QPM hybrids for grain yield and most of the other measured traits evaluated under drought, low and
optimum N environments.
Under combined low, optimum, and across all N environments, greater contributions of GCA sum of squares than SCA sum of squares were observed for days to anthesis and silking, indicating the greater importance of additive gene action than non-additive gene actions in the inheritance of the traits. A greater reciprocal sum of squares than GCA sum of squares were observed for number of ears per plant, plant aspect, ear length and diameter under combined low N environments. This indicates that the inheritances of the traits were more affected by the additive gene actions of the maternal cytoplasm genes than the additive gene actions of the crosses nuclear genes for ears per plant, plant aspect, ear length, and diameter under combined low N environments. Different authors have also reported different results for contributions of the GCA and SCA sum of squares for QPM and non- QPM maize hybrids under multiple stress environments (Badu-Apraku et al., 2015a and Njeri et al., 2017)who reported greater magnitude of SCA sum of squares than GCA sum of squares of QPM germplasm for grain yield under drought stress. Wegary et al. (2014)also reported a high contribution of non-additive gene actions for the inheritance of grain yield, ears per plant and plant height; while there was a greater importance of additive gene actions for the inheritance of days to anthesis and silking under low N environment for QPM hybrids. More importance of non-additive gene actions than additive gene action for grain yield under low N environments for non-QPM hybrids was reported by Worku et al. (2008).
Significant and positive GCA effects were observed in TL156579, TL156583, TL148288, and TL156612 parental lines, for grain yield across N environments, indicating that the inbred lines are good general combiners for grain yield across N environments; and the inbred lines can be used as donor parents for the target trait under the target environments. Different effects of SCA were observed in hybrids under low and optimum N environments, indicating that selection of hybrids based on only SCA effects is not a guarantee for the selection of the best grain yielding hybrids under target environments. Menkir et al. (2004)recommended that for better hybrid development, parental lines should have to be grouped into heterotic groups based on the grain yield performance and SCA effects of the inbred lines.
For effective maize hybrid development, the classification of maize inbred lines into appropriate groups could be essential in a maize breeding program that maximizes the importance of inbred lines (Badu-Apraku et al., 2015b). In this study, classification of inbred lines into heterotic groups was done based on SCA effects for grain yield and grain yield performance of the crosses as suggested by Fan et al. (2013). Because, asper se performance of inbred lines is not a good indicator of the hybrids grain yield performance for maize due to the strong dominance effects in maize (Badu-Apraku and Akinwale, 2011). Inbred lines were classified into 3 heterotic groups under low, optimum and across all N environments, which maximize the development of superior maize hybrids under stress and non-stress soil environments. Different researchers have used different methods for inbred lines classification into heterotic groups to maximize the potential importance of inbred lines for the development of productive hybrids. Badu-Apraku et al. (2015a)used heterotic grouping based on GCA of multiple traits (HGCAMT) methods for QPM inbred lines using a classification into 3 heterotic groups under a low N environment. Fan et al. (2009)classified 23 normal maize inbred lines into 4 known maize heterotic groups by using the heterotic group’s specific and general combining ability (HSGCA) method. Non-QPM inbred lines were classified into 4 heterotic groups based on SCA effects for yield and yield performances of the crosses (Fan et al., 2013).
Mean squares for GCA, SCA and reciprocal effects showed significant for grain yield and other major phenotypic traits both under low and optimum N environments. Under low and optimum N environments, additive gene actions were more important than non-additive gene action for days to anthesis and silking; whereas, non-additive gene actions were more important than additive gene action for grain yield, the number of ears per plant, plant and ear aspects, plant and ear heights, ear length and diameter.
Single cross hybrids VL05128 x TL156612 and TL156612 x TL155932 were selected as high yielding hybrids, which out-yielded 16.76% and 14.14%, respectively, more than the best check (4.07 t ha-1) under low N environments; while hybrids TL156583 x TL156612, TL156612 x TL148288, TL148287 x TL156583, TL156612 x TL156583, and TL156583 x TL148287 were selected as single cross hybrids; and these out-yielded the best check under optimum N environments.
Parental lines TL156579, TL156583, and TL148288 were considered as a good general combiner for grain yield under low N environments. Parental line TL156612 showed higher positive GCA effects when used as female; while parental line TL156612 showed higher negative GCA effects when used as the male parent for grain yield under low N environments. Under low N environments, hybrids TL156591 x TL155976, TL116960 x TL147078 and TL156583 x TL155932 were recorded as showing high SCA effects for grain yield; while hybrids TL156583 x TL148287, TL148288 x TL155976 and TL148287 x TL155932 were recoded as showing high SCA effects for grain yield under optimum N environments.
CSA, Central Statistical Agency; FAO, Food and Agricultural Organization; GCA, general Combining Ability; IBPGR, International Board for Plant Genetic Resources; Low N, Low N environment; MAT, Maternal effect; Optimum N, optimum N environment; QPM, Quality protein Maize; SCA, Specific Combining ability.
The authors have not declared any conflicts of interests.
The authors highly thank the community of Haramaya University and West Africa Center for Crop Improvement, University of Ghana for their continued support during the study and field work, and also thank CIMMYT for providing QPM lines for the study. This work was supported by DAAD and ECONET through the West Africa Center for Crop Improvement.
REFERENCES
|
Babu R, Prasanna BM (2014). Molecular Breeding for Quality Protein Maize ( QPM ). International Maize and Wheat Improvement Center pp. 489-505.
Crossref
|
|
|
|
Badu-Apraku B, Menkir A, Ajala SO, Akinwale RO, Oyekunle M, Obeng-Antwi K (2010). Performance of tropical early-maturing maize cultivars in multiple stress environments. Canadian Journal of Plant Science 90(6):831-852.
Crossref
|
|
|
|
|
Badu-Apraku B, Akinwale RO (2011). Identification of early-maturing maize inbred lines based on multiple traits under drought and low N environments for hybrid development and population improvement. Canadian Journal of Plant Science 91(5):931-942.
Crossref
|
|
|
|
|
Badu-Apraku B, Fakorede MAB, Talabi AO, Akaogu IC, Annor B, Melaku G, Aderounmu, M (2015a). Gene Action and Heterotic Groups of Early White Quality Protein Maize Inbreds under Multiple Stress Environments. Crop Science 56:183-199.
Crossref
|
|
|
|
|
Badu-Apraku B, Annor B, Oyekunle M, Akinwale RO, Fakorede MAB, Talabi AO, FasanmadeY (2015b). Grouping of early maturing quality protein maize inbreds based on SNP markers and combining ability under multiple environments. Field Crops Research 183:169-183.
Crossref
|
|
|
|
|
Badu-Apraku B, Yallou CG, Obeng-Antwi K, Alidu H, Talabi AO, Annor B, Aderounmu M (2017). Yield Gains in Extra-Early Maize Cultivars of Breeding Eras under Multiple Environments. Agronomy Journal 109:418-431.
Crossref
|
|
|
|
|
Badaway MM (2013). Heterosis and Combining ability in maize using Dialleel Crosses among Seven new Inred Lines. Asian Journal of Crop Science 5(1):1-13.
Crossref
|
|
|
|
|
Below FE (1997). Growth and productivity of maize under stress. In: Edmeades GO, Banziger M, MIckelson HR, Pena-Valdivia CB (Eds.). Developing Drought and Low N-Tolerant Maize. Proceedings of a Symposium. 25-29 March 1996, CIMMYT, Mexico, D.F, Mexico pp. 235-240.
|
|
|
|
|
Baker RJ (1978). Issues in the diallel analysis. Crop Science 18(4):533-536.
Crossref
|
|
|
|
|
Banzinger M, Edmeades GO, Beck D, Bellon M (2000). Breeding for Drought and Nitrogen Stress Tolerance in Maize. From Theory to Practice. Mexico, D.F. CIMMYT.
|
|
|
|
|
CSA (2019). The federal democratic republic of Ethiopia's central statistical agency reports on area, production and farm management.
View
|
|
|
|
|
Dabholkar AR (1999). Elements of Biometrical Genetics. Ashok Kumar Mittal Concept Publishing Company, New Delhi, India 244 p.
|
|
|
|
|
Erisman JW, Galloway JN, Seitzinger S, Bleeker A, Dise NB, Petrescu AMR, de Vries W (2013). Consequences of human modification of the global nitrogen cycle. Philosophical Transactions of the Royal Society. 368, 20130116. Facette MR, Shen Z, Bjönsdót.
Crossref
|
|
|
|
|
Fan XM, Zhang YD, Bi YQ, Liu L, Chen HM, Kang MS, Chen HM (2013). Reciprocal Diallel Crosses Impact Combining Ability, Variance Estimation, and Heterotic Group Classification. Crop Science 54(1):89-97.
Crossref
|
|
|
|
|
Fan XM, Zhang YM, Yao WH, Chen HM, Tan J, Xu CX, Kang MS (2009). Classifying Maize Inbred Lines into Heterotic Groups using a Factorial Mating Design. Agronomy Journal 101:106-112.
Crossref
|
|
|
|
|
Farfan IDB, Fuente GND, La Murray SC, Isakeit, T, Huang C, Warburton M, Kolomiets M (2015). Genome-Wide Association Study for Drought, Aflatoxin Resistance, and Important Agronomic Traits of Maize Hybrids in the. PLoS ONE 10:1-30.
Crossref
|
|
|
|
|
Gelli M, Duo Y, Konda AR, Zhang C, Holding DR, Dweikat IM (2014). Identification of differentially expressed genes between sorghum genotypes with contrasting nitrogen stress tolerance by genome-wide transcriptional profiling. Agronomy and Horticulture 15:1471-2164.
Crossref
|
|
|
|
|
Griffing B (1956). Concept of General and Specific Combining Ability in Relation to diallel crossing systems. Australian Journal of Biological Sciences 9(4):463-493.
Crossref
|
|
|
|
|
Hallauer AR, Carena MJ, Filho JBM (2010). Quantitative genetics in Maize breeding 2nd Ed. Springer New York, USA.
Crossref
|
|
|
|
|
IBPGR (1991). Descriptors for maize. International Maize and Wheat Improvement Center, Mexico City/international Board for Plant Genetic Resources. Rome pp. 29-56.
|
|
|
|
|
Mahgoub GMA (2011). Partitioning of General and Specific Combining Ability Effects for Estimating Maternal and Reciprocal Effects. Journal of Agricultural Science 3(2):213-222.
Crossref
|
|
|
|
|
Menkir A, Melake-Berhan A, The C, Ingelbrecht I, Adepoju A (2004). Grouping of tropical mid-altitude maize inbred lines based on yield data and molecular markers. Theoretical and Applied Genetics 108(8):1582-1590.
Crossref
|
|
|
|
|
Moll RH, Kamprath EJ, Jackson WA (1982). Analysis and interpretation of factors that contribute to the efficiency of nitrogen utilization. Agronomy Journal 74(3):562-564.
Crossref
|
|
|
|
|
Njeri SG, Makumbi D, Warburton ML, Diallo A, Jumbo MB, Chemining G (2017). Genetic analysis of tropical quality protein maize (Zea mays L.) germplasm. Euphytica 213(11):1-19.
Crossref
|
|
|
|
|
Presterl T, Seitz G, Landbeck M, Thiemt EM, Schmidt W, Geiger HH (2003). Improving nitrogen-use efficiency in European maize: Estimation of quantitative genetic parameters. Crop Science 43(4):1259-1265.
Crossref
|
|
|
|
|
Ranum P, Pena-Rosas JP, Garcia-casal MN (2014). Global maize production, utilization, and consumption. New York Academy of Science 1312(1):105-112.
Crossref
|
|
|
|
|
Reif JC, Hallauer AR, Melchinger AE (2005). Heterosis and heterotic patterns in maize. AGRIS 50:215-223.
|
|
|
|
|
Rodríguez F, Alvarado G, Pacheco A, Burgueno J, Crossa J (2018). AGD-R (Analysis of Genetic Designs with R for Windows) Version 5.0. CIMMYT, Km. 45 Carretera Mexico-Veracruz, El Batán, Texcoco, Estado de México, México, C.P. 56237.
|
|
|
|
|
SAS Institute, Inc. (2012). Statistical analysis software (SAS) user's guide. SAS Institute, Inc., Cary, NC.
|
|
|
|
|
Wang B, Wang L, Li D, Özkan N, Li S, Mao Z (2009). Rheological properties of waxy maize starch and xanthan gum mixtures in the presence of sucrose. Carbohydrate Polymers 77(2):472-481.
Crossref
|
|
|
|
|
Wegary D, Vivek BS, Labuschagne MT (2014). Combining Ability of Certain Agronomic Traits in Quality Protein Maize under Stress and Nonstress Environments in Eastern ... Combining Ability of Certain Agronomic Traits in Quality Protein Maize under Stress and Nonstress Environments in Eastern and South Africa. Crop Science 54(3):1004-1014.
Crossref
|
|
|
|
|
Worku M, Bänziger M, Friesen,D, Schulte G, Horst WJ, Vivek BS (2008). Relative Importance Of General Combining Ability and Specific Combining Ability Among Tropical Maize (Zea Mays L.) Inbreds Under Contrasting Nitrogen Environments. Maydica 53:279-288.
|
|
|
|
|
Worku Mosisa, Tuna H, Abera W, Wolde L, Diallo A, Afriyie ST, Guta A (2001). Developing Low-N Tolerant Maize Varieties for Mid-Altitude Sub- Humid Agro-Ecology of Ethiopia. Seventh Eastern and Southern Africa Regional Maize Conference 11th - 15th February, pp. 197-201.
|
|
|
|
|
Yao WH, Zhang YD, Kang MS, Chen HM, Liu L (2013). Diallel Analysis Models?. A Comparison of Certain Genetic Statistics. Crop Science 53(4):1480-1490.
Crossref
|
|