ABSTRACT
The effects of liquid nitrogen fertilizer (150% N) and benzyl amino purine (BAP 200 ppm) on senescence, yield and nutrients mobilization in two cowpea varieties were investigated in the rainy and dry seasons. The study was conducted in a screen house at the University of Lagos, Nigeria (6°27ʺ1, 3°45ʺ E). The duration of the study was between August 2012 and January 2013. The experimental design used was completely randomized block design. The four treatments (nitrogen fertilizer, BAP, nitrogen fertilizer and BAP combination in 3:1 ratio and the control) were applied through foliar spray at 3, 6 and 9 weeks after planting and data on time of senescence and yield was collected. The days to 50% senescence, 90% senescence and death of the plants was earlier in IT89KD-288 than in Kanannado, while senescence started earlier in plant treated with liquid nitrogen fertilizer (15%N). For the first harvest, variety IT89KD-288 had higher number of pods (10), number of seeds (13) and length of pods (15.8 cm) than kanannado that had 8.0, 12.0, 14.68 cm, 3.17 and 18.53 for number of pods, number of seeds, length of pods, weight of grains per pod and weight of grains per plant, respectively, while at the second harvest, kanannado had higher number of pods (5), number of seeds (10), length of pods (12.12 cm), weight of grains per pod (2.68) and weight of grains per plant (12.03) than IT89KD-288 that had 4.0, 9.0, 11.31 cm, 2.05 and 11.27 for number of pods, number of seeds, length of pods, weight of grains per pod and weight of grains per plant, respectively. The combined treatment induced significantly greater yield than the other treatments. Yields were higher in the rainy season than dry season. The longer the duration of senescence, the higher the grain yield at the second harvest; therefore, confirming a direct relationship between senescence, harvest time and yield among cowpea varieties.
Key words: Cowpea, benzyl amino purine, nitrogen, kanannado, fertilizer.
Cowpea, Vigna unguiculata (L.) walp, subspecies unguiculata is a dicotyledenous spermatophyte belonging to the family fabaceae. The plant is an annual crop grown virtually all over the world. There are several cultivars adapted to different world climatic regions. The crop is widely cultivated in Nigeria for the consumption of its leaves, green pod and grain. The herbage can also be used as green manure and animal feed (Steele, 1976). Cowpea is an important food grain legume for over 200 million people in the dry savanna of tropical Africa. It is particularly important in West Africa with over 9.3 million metric tonnes of annual production (Oritz, 1998). The grain is a good source of human protein, while the haulms are valuable source of livestock protein (Fatokun, 2015). It is also a source of income for many smallholder farmers in sub-Saharan Africa and contributes to the sustainability of cropping systems and soil fertility improvement in marginal lands through provision of ground cover and plant residue, nitrogen fixation and suppressing weed. However, despite its great importance, grain yield of cowpea crop is low, about 300 kg ha-1 (Cardoso et al., 2015; Leite et al., 2015).
When compared with many other crops, cowpea has received little attention from plant breeders and large efforts need to be made to break the yield barriers and for cowpea production to keep pace with other crops, especially cereals, its yield potential must be improved (Anonymous, 2014). In Nigeria, 80% of the cowpea produced mainly as grain is from the savanna zone of the country (FAO, 1999). A wide range of
seed yields has been recorded for cowpeas but is generally low. Among factors responsible for the low yields is low soil fertility, as most tropical soils are deficient in essential nutrients, particularly nitrogen and phosphorus (Jones and Wild, 2013). Traditionally, soil fertility in West Africa has been maintained through fallow. However, in Nigeria, intensive cropping is gradually replacing the traditional shifting cultivation that is associated with long fallow and hence low crop yield. The steady decline in food production due to reduced length of fallow on land has prompted farmers to amend soil with different materials (organic and inorganic) to enhance plant growth and increase yield (Adepetu, 2013).
It has been suggested that organic manure should be used in place of chemical fertilizer to avoid long-term negative effects of chemical fertilizer on the soil. However, organic manure is usually required in large quantity to sustain crop production and may not be available to the small-scale farmers (Nyathi and Campbell, 1995), hence, the need for inorganic fertilizer. The positive effect of the application of inorganic fertilizers on crop yields and yield improvement have been reported (Carsky and Iwuafor, 2013). Although, cowpea symbiotically fixes nitrogen, plant dependent on symbiotically fixed N may well suffer from temporary N deficiency during the seedling growth once the cotyledonary reserves have been exhausted. Usually, prior to the onset of symbiotic N fixation, cotyledonary reserves are mobilized during hypocotyl elongation in cowpea and cotyledons are usually shed one or two days from emergence. It has thus been recognized and demonstrated that application of a small quantity of nitrogen fertilizer enhances early vegetative growth (Dart et al., 2007).
Burris (2014) stated that nitrogen has a stimulating effect on root activity and rooting pattern of the crop. It has also been reported that available nitrogenous compound allowed seedlings to make a good start before nitrogen fixation has a chance to occur. Other workers have shown that plants given inorganic N during vegetative periods were much larger by the onset of flowering than those dependent on symbiotic N fixation (Minchin et al., 2014). Such plants also had more branches and produced many peduncles resulting in greater number of pods, seeds and significantly larger yields. There are many reported studies on the effects of P application on growth and yield of cowpea (Owolade et al., 2006; Kolawole et al., 2002; Okeleye and Okelana, 2013) there is dearth of information on the effects of N fertilizer on growth and yield of cowpea in Nigeria. However, it has been reported elsewhere that the main limiting nutrients for legume production in the tropics are N and P (Fox and Kang, 2009).
The term senescence is basically derived from the Latin verb senescere meaning “to grow old”; generally, the most obvious senescence in plants is foliar senescence (leaf senescence). In fact, the leaf senescence is the last stage of leaf development during which the leaf color changes from green to yellow (Keech et al., 2007). Normally, leaf senescence is initiated by yellowing of the margins of the leaf blade extending towards center of the leaf blade near the midrib, resulting in death of the leaf. Leaf senescence although deteriorative in nature has been recognized as the last phase of the organs development, a highly ordered process regulated by genes known as senescence associated genes (SAGs) (Pruitt, 1983). The leaf when young and mature accumulates nutrients and exports them to growing parts of the plant during senescence.
In cowpea, senescence causes substantial reduction in total grain yield because most cowpea plants die after producing the first flush of pods. The reduction in yield is most drastic in the local varieties. Delaying leaf senescence will most probably extend the reproductive period and increase the photosynthetic efficiency of the crop resulting in increased grain yield. The objective of the experiment is to investigate the combined effect of liquid nitrogen fertilizer with cytokinin (BAP) in regulating the onset of senescence in some cowpea varieties and also to determine the combined effect of nitrogen fertilizer and BAP on yield of two cowpea varieties.
Study area
This study was conducted in the screen house at the University of Lagos located in the south-western part of Nigeria, latitude 6°27ʺN, longitude 3° 45ʺ E and altitude 0 to 41 m (0-135 ft) above sea level with a tropical wet and dry climate of two peaks of rainy seasons, from April to July and between October and November. There is a brief relatively dry spell between August and September and a longer dry season from December to March. Monthly rainfall averages over 400 mm (16 in) between May and July, 200 mm (7.9 in) in August and September and as low as 25 mm (0.98 in) in December. The temperature ranged from a maximum of 37.3°C to a minimum of 13.9°C (World Weather Information Service Lagos, 2012).
Source of seeds
Kanannado, a local cowpea variety and IT 89KD–288, an improved variety which are photosensitive, were collected from IITA, Ibadan. In cowpea, senescence causes substantial reduction in total grain yield because most cowpea plants die after producing the first flush of pods. The reduction in yield is most drastic in the local varieties of which kanannado and IT89KD-288 are among the most affected hence their selection for the experiment.
Kanannado
This is a local variety that originated from Kano in the northern part of Nigeria. It is strongly photosensitive and late maturing, requiring about 80 to 114 days to flower during the rainy season when average minimum and maximum temperatures are between 19.6 and 32.5°C, respectively and day length range from 13.6 to 12.9 h day-1. It has an indeterminate growth habit with lateral branches growing up to 3 m long during the wet season. On the other hand, when such plants are grown where there is no rainfall and day length is short (about 12.2 to 12.7 h day-1) with minimum and maximum temperatures between 15.2 and 30.7°C respectively, their growth becomes stunted due to lack of elongation of the internodes. Branching habit is also affected in this case and the plants assume a rosette appearance. However, maturity occurs early (takes 40 to 45 days from sowing to flowering) (Singh, 1997). They have very dark green leaves and white flowers, pods are non-pigmented, coiled with thin pod walls. Seeds are large, rough and white with brown hilum.
IT89KD–288
IT89KD–288 is a cultivar derived from the cross between IT897F–1772 (Kanannado selection) and IT845–2246–4, like Kanannado, it is strongly photosensitive and late maturing requiring similar number of days to flower during the rainy season. Growth is indeterminate with lateral branches up to 3 m long. Because of its photosensitive nature, there is stunting growth in the dry season when day length is short; however, maturity is early, leaves are dark green, flowers are white, the pods are unpigmented, slightly curved and seeds are large, rough and white. It yields more fodder and grain than Kanannado. It combines resistance to aphids, bruchids and thrips.
Planting
Plastic pots of 250 mm diameter were used in this study. 240 pots were used for the two varieties, 120 pots for each variety. They were filled with fresh sandy loam top soil and watered well for 2 days before planting. The treatments were replicated 8 times in completely randomize block design. The treatments used include benzyl amino purine (BAP) and liquid N fertilizer (Boost Extra). There were also combined treatments of BAP and liquid N fertilizer. The 1st planting was done August 9th, 2012 for the rainy season and the 2nd planting was done January 20th, 2013 for the dry season. Seeds were directly sown in the prepared pots (after surface treatment with fungicide Apron plus) with 2 seeds per pot. They were labeled appropriately using white and green plastic tags according to the applied treatment.
Experimental treatments
Liquid nitrogen fertilizer
The type of liquid fertilizer used was Boost Extra, the composition of this fertilizer includes: nitrogen 20%, phosphate 20%, potassium 20%, magnesium 1.5%, iron EDTA 0.15%, manganese EDTA 0.075%, copper 0.075%, zinc 0.075%, boron 0.0315%, cobalt EDTA 0.0012% and molybdenum 0.0012%. The pH of the solution (10% solution) was 4.0 to 4.5. One hundred millilitres (100 ml) of the liquid fertilizer (Boost Extra) was added to 15 L of water, mixed together and applied to the plants using foliar application technique. The fertilizer was applied at 3, 6 and 9 weeks after planting.
Hormone treatment
The hormone used in this research was BAP at 200 ppm concentration and there were 15 replications per treatment. A foliar spraying technique was used to apply the treatment at three doses: at 3, 6 and 9 weeks after planting. The control plants were sprayed foliarly with water in 3 doses every week.
Combined treatments
The combined treatments comprise of a combination of the hormone and the liquid fertilizer, the liquid fertilizer was applied first and the plants were subsequently sprayed with cytokinin (BAP) at 3:1 ratio of liquid fertilizer and hormone, respectively.
Data collection
Data was collected on senescence, the changes in leaves color were observed visually and the extent of chlorophyll loss was monitored. Days to onset of senescence were counted and recorded for each plant, likewise days to 50% of senescence, days to 90% of senescence and days to complete death of the plant. From the obtained data, the duration of senescence were estimated in order to determine their effects on senescence in the cowpea varieties. Data was also collected on yield as follows:
Number of pods
Number of pods for each treated and control plants were counted and recorded.
Length of pods
The length of each pod of the plants was measured with the aid of a meter rule and a thread and the values were recorded.
Number of grains per pod
The number of grains per pod for each plant was counted and recorded.
Weight of grains per pod
The weight of grains per pod of each treatment and control plants were weighed and recorded using a weighing balance. The weighing balance was adjusted to 0. The grains were then removed from the pods and placed on the weighing balance, and the weight the grains recorded.
Weight of grains per plant
The dry weight of the grains of the harvested fruits of all the cowpea varieties giving hormone and fertilizer treatments were recorded as mean weight of seeds per plant and taken as yield per treatment.
Statistical analysis
The data collected was subjected to one-way analysis of variance (ANOVA) using GENSTAT software and means with significant differences were separated using least significance difference test at P<0.05.
Senescence
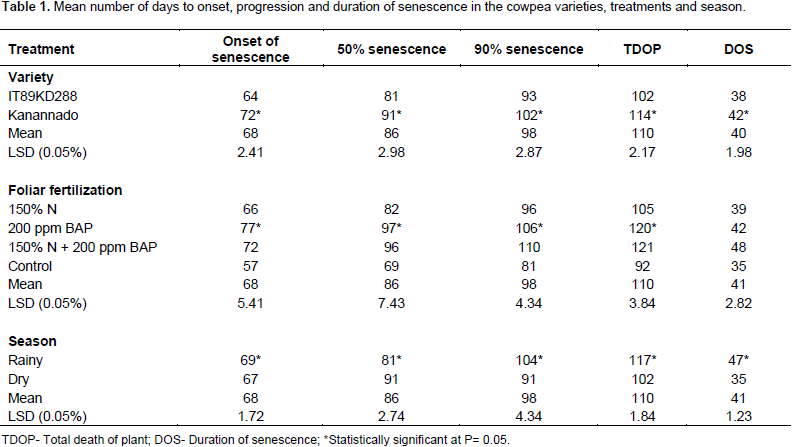
The result of onset, progression and duration of senescence in the two cowpea varieties, is presented in Table 1. Comparison of the varieties showed significant difference (P≤0.05) in all the parameters. In the variety Kanannado, there was a general delay in the number of days taken to the onset of senescence and days to 50% senescence, days to 90% senescence, and days to total death of the plant, when compared with IT89KD–288 (Table 1). The effect of the various treatments on senescence is also shown in Table 1. The treatments induced different responses for the number of days taken to the onset of senescence, days to 50% senescence, days to 90% senescence, and days to total death of the plants. Comparison between the treatments and the control showed that nitrogen fertilizer hastened senescence, while BAP treatment applied as a single treatment or in a combination with N fertilizer delayed senescence. Nitrogen fertilizer is not known to delay senescence.
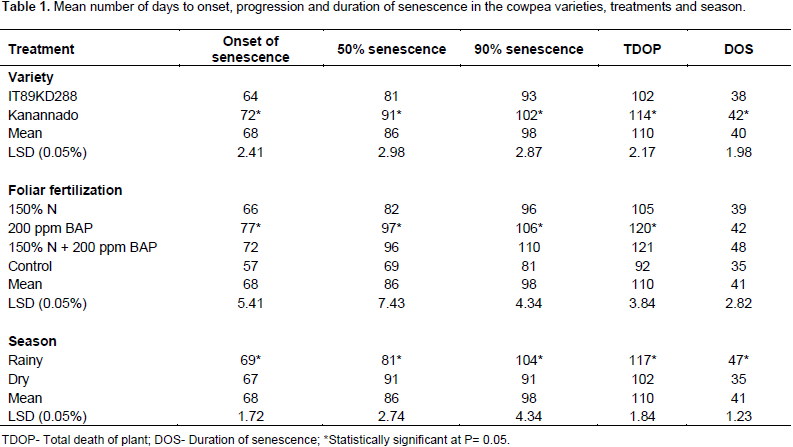
Several studies reported that N fertilizer is known to improve soil productivity and fertility which improved yield and quality of crops (Whalen, 2000; Maerere and Ishimine, 2001; Vanek, 2003). Soil treated with N fertilizer was found to be loose and this probably provided adequate aeration in the soil and improved microbial activities (Xio and Li, 2006). On the other hand, turmeric plant when treated with nitrogen fertilizer remained green for a longer time and resulted in a higher vegetative growth and yield (Mazid, 1993; Seobi, 2005; Anes and Johnson, 1980). Nitrogen treatments (150% N) hastened the onset of senescence by 66 days, days to 50% senescence by 82 days, 90% by 96 days and total death by 105 days. The combined treatment (150% N + 200 ppm BAP) induced delay in the onset of senescence by 72 days, 50% senescence by 96 days, 90% senescence by 110 days and total death of plant by 121 days (Table 1). Treatment with 200 ppm BAP also delayed the onset of senescence (77 days), and 50% senescence (97 days), 90% senescence (106 days) and total death of plant (120 days).
There was no significant difference between 200 ppm BAP treated plants and the combined treatment of 150% nitrogen fertilizer + 200 ppm BAP with respect to 90% senescence and total death of plants. Several workers such as Nooden (1978) and Richmond and Lang (1994), reported that BAP and gibberellins retard senescence, while abcissic acid and ethylene tend to act as accelerators. Leaf senescence can be retarded locally by the application of BAP (Schuphan, 1974). Physiological studies suggest that BAP can regulate leaf senescence and that the internal BAP level drops with the progression of leaf senescence (Schuphan, 1974). Senescence is the result of complex changes in basic plant metabolism. In higher plants, various degradative phenomena associated with free radicals (FRs) have been implicated in the senescence process (Leshem et al., 1986; Thompson et al., 1987). Onset of senescence in the control plants occurred at 57 days after planting, 50% at 69 days, and 90% at 81 days and total death at 92 days after planting (Table 1).
The duration of senescence from its onset to total death of plants ranged from 35 days in the control to 39 days in 150% N to 42 days in 200 ppm BAP to 48 days in 150% N + 200 ppm BAP (Table 1). This shows that reproductive period in the combined treatment of 150% N + 200 ppm BAP was most extended by about 13 days. Effect of the two planting seasons (rainy and dry) on senescence is presented in Table 1. Onset of senescence, days to 50 and 90% senescence and total death of the plants were earlier for the dry season planting when compared with the rainy season planting with significant difference (P≤ 0.05). In the rainy season, senescence commenced from 69 days after planting (DAP) and total death of the plants occurred by 117 DAP, whereas onset of senescence in the dry season was recorded at 67 DAP and total death at 102 DAP (Table 1). The duration of senescence from onset to total death were 35 days in the dry season and 47 days in the rainy season (Table 1) suggesting that reproductive period was extended by about 12 days in the rainy season.
Yield
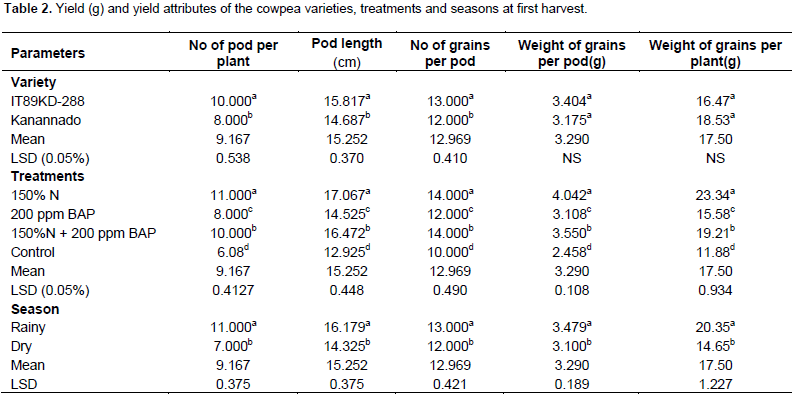
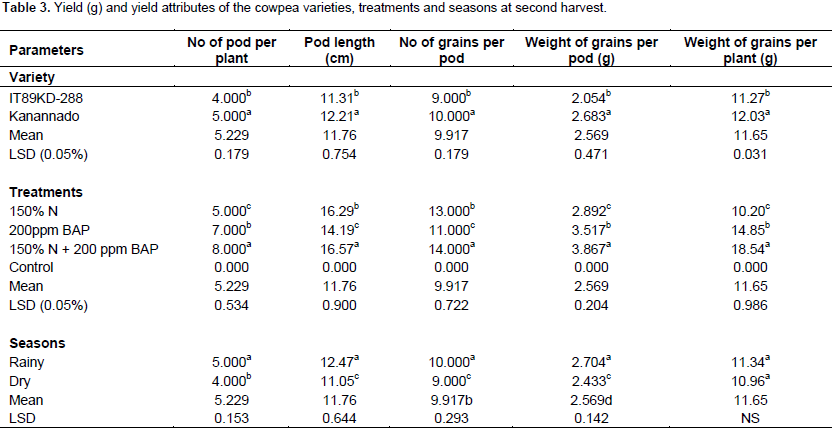
There was significant difference between the two varieties of cowpea used in this experiment with respect to yield parameters. In the first harvest, variety IT89KD – 288 had greater number of pods, number of seeds and length of pods. There was exception in weight of grains per pod and weight of grains per plant in which no significant difference was observed (Table 2), but during the second harvest, kanannado variety had greater number of pods, number of seeds, length of pods, weight of grains per pod and weight of grains per plant when compared with IT89KD-288 (Table 3). This might be due to the early commencement of senescence in IT89KD-288 that led to the death of most of the plants before the2nd harvest. However, IT89KD-288 produced less yield when compared with kanannado that had a delay in the onset of senescence and thus most of its plants were alive before the 2nd harvest and therefore produced more yield in the 2nd harvest.
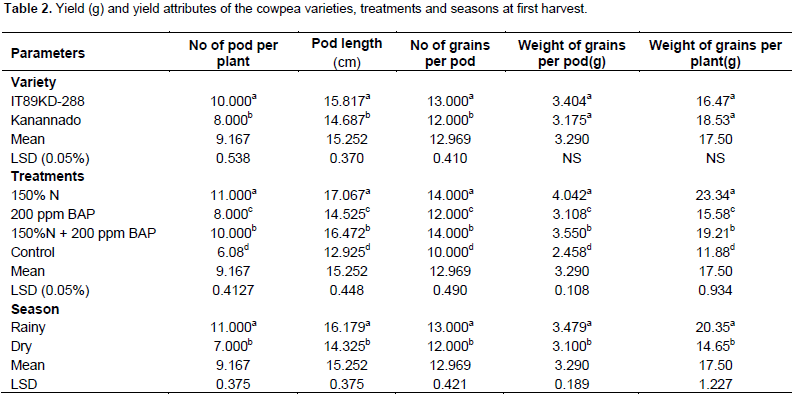
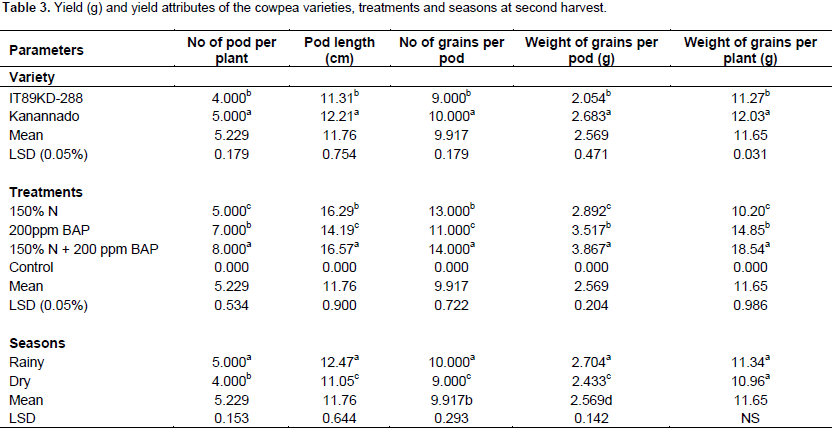
Fertilizer and hormonal treatments increased yield in the cowpea varieties grown in the different seasons with significant difference when compared with the control plants. Combined treatment of 150% N fertilizer and 200 ppm BAP treatment induced significantly higher yield in the two harvests (1
st and 2
nd harvests) as compared to the rest of the treatments. This might be due to the early commencement of senescence in the other treatments that led to the death of the plants before the 2
nd harvest which subsequently led to low yield. The control plants did not have any yield as a result of the death of all the plants before the 2
nd harvest (Table 3). It was reported that in cowpea, many plants often die after producing the 1
st flush of pods. This causes substantial reduction in total grain yield in that 2
nd flush yield is proportional to the number of plants surviving to produce the 2
nd flush (Ismail and Hall, 1998.) Several authors have reported the same observations on some cowpea varieties (
Ferry and Singh, 1997;
Singh, 2002;
Boukar et al., 2015).
The delayed leaf senescence result from a higher proportion of plants surviving after the production of the 1st flush of pods and probably results from the maintenance of root viability (Gwathmey and Madore, 1992; Fatokun et al., 2013) which could enhance nitrogen fixation. The single hormone treatments, that is, 200 ppm BAP did not show significant effect on yield when compared with the control plants. Certain hormones have been found to increase yield in many crops, while some other growth substances do not have significant effect on yield. For instance, the auxin, b–naphthoacetic acid (NAA) sprayed on the open flowers of Solanum melongena either singly or in combination were observed to increase fruit set as well as total weights of the fruits (Olympios, 1976). Subramanian and Kende (1985) observed that growth substances influenced seed yield in cowpea even though the yield component were not significantly affected.
Kaul et al. (1976) reported that an increase in number of pods/plant by seed treatment with 200 ppm planotix increased yield by 33%. Thomas and Stoddart (1976) reported that auxins have been used for many years to increase fruit set of tomatoes. Several workers showed the importance of N fertilizer, in increasing grain yield and its components. Nour (1998) and El-Kholy et al. (1999) found that application of fertilizer significantly increased grain yield in rice when compared with the control. Ebaid and Ghanem (2000) reported that panicle weight, 1000-grain weight and grain yield in rice were significantly increased as the N fertilizer increased up to 30 tons/ha, while 70 tons/ha was adequate for the highest values of panicle length and number of grain/panicle. Mazid (1993) reported that N fertilizer should be applied in a particular ratio, for higher growth and yield of a specific plant species. El-Batal et al. (2004) showed that increasing N fertilizer rate from 50 to 80 kg N/feed significantly increased plant height and yield. Vanek (2003) reported that regular application of N fertilizer to root crops leads to higher yield.
Comparing the 2 seasons during the 1st harvest, greater yield was obtained in rainy season with significant difference (Table 2). In the 2nd harvest, rainy season differed significantly from the dry season in the number of pods, length of pods, number of grains per pod, weight of grains per pod but no significant difference with regards to weight of grains per plant (Table 3). Wallace (1985) and Summerfield et al. (1975) demonstrated positive relationship between yield and photoperiod temperature response; the increase in yield is due to increase in number of nodes at which pods could be set, indicating that the nodes arose from new branches and continued elongation of existing indeterminate stem and branches. They also reported that plant size at flowering and the number of nodes produced has a great influence on subsequent yield in indeterminate genotypes.
They noted that stunted plants due to adverse conditions gave poor yield. Grain yield in cowpea is dependent on both vegetative and reproductive component that are in turn governed by environmental factors such as day length, temperature and soil moisture (Chaudhry and Ogo, 1985). Economic yield is expected to show close positive relation with total plant dry weight, 50% of which to a large extent is dependent on number of leaves, plant height and number and length of branches (Summerfield et al., 1975). As recent examples,
Souleymane et al. (2013) and
Huynh et al. (2015) confirmed that with the improved variety, IT97K-556-6.
Grain yield at 1st and 2nd harvest and the total yield
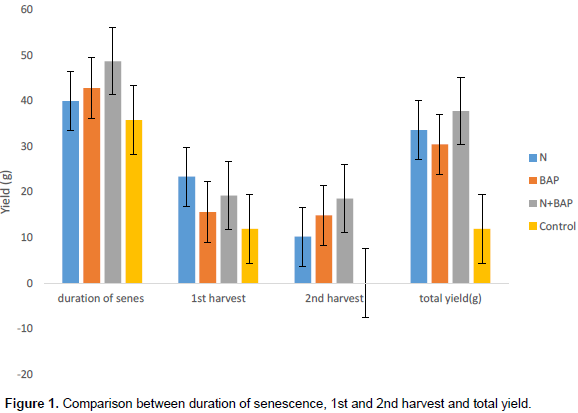
Table 4 shows the grain yield at 1st and 2nd harvest and the total yield with respect to the treatments. At 1st harvest, 150% N treated produced greater yield than the combined treatment of 150% N + 200 ppm and then the 200 ppm BAP treatments, and the least were the control plants. In the 2nd yield, 150% N + 200 ppm produced greater yield than the rest of the treatments. Therefore, at the end when the 1st and 2nd harvest were added together, the total yield was greater in 150% N + 200 ppm followed by 150% N then 200 ppm BAP, the least in the total yield were the control plants (Table 4).

Relationship between duration of senescence, 1st and 2nd harvest and the total yield in relation to treatments
The relationship between senescence, 1st and 2nd harvests and the total yield showed that the combined treatments of 150% N + 200 ppm BAP took longer days from the onset of senescence to the total death of the plants followed by the 200 ppm BAP treatment, then 150% N. Therefore, due to the delayed senescence in the 150% N + 200 ppm BAP and single 200 ppm BAP treatments, they were able to produce 2nd flush of pods which led to greater yield in the 2nd harvest and the total grain yield when compared with 150% N treatment that took lesser days from the onset of senescence to the total death of the plants, and therefore produced less yield in the 2nd harvest and total yield, and this shows that the longer the days the plant takes to senesces, the greater the yield (Figure 1). A relationship between crop yield and senescence has been postulated for many years, Thomas et al. (1996) assumed that an extended period of maximal photosynthetic activity, that is, delayed senescence, should lead to higher yields.
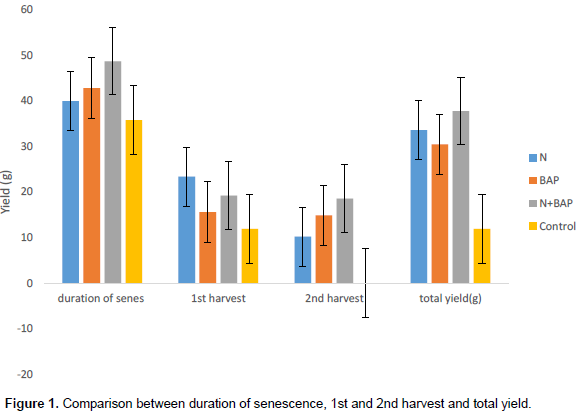
Whereas a positive correlation between leaf senescence duration and yield may be valid for most crops with regard to total biomass production and for tuber crops; the relationship is more complicated with respect to seed yields. In particular for cereal crops, there has been a long discussion on whether grain yield is determined by sink (the developing grain) or by source (the photosynthesizing vegetative tissues). The predominant current view is that sink strength is the main limiting factor for yields, especially in small-grain cereals such as wheat (Fischer, 2008; Boukar et al., 2013) and, hence, physiological events particularly in the period around seed setting are crucial for determining yield levels. The genetically determined senescence program can indeed have bearings on the productivity of crop plants including seed yields (Borra´s et al., 2004; Huynh et al., 2016).
The association among senescence parameters and yield are found in sorghum and in particular corn. A classical study on 10 short season maize hybrids showed a positive correlation between leaf senescence duration and grain yield (Tollenaar and Daynard, 1978). Similar relationship between leaf area duration/stay-green and yield parameters were reported for sorghum hybrid lines (Borrell et al., 2000) and also for oilseed rape (Hunkova et al., 2011). In addition to these strong examples, there are many studies involving a number of species which demonstrates that genetic modulation of senescence parameters can indeed affect the yield levels, even though the effects are variable and highly influenced by environmental conditions.
In conclusion, the combined liquid nitrogen fertilizer and BAP regulated the onset of senescence and had higher overall yield when compared with the other treatments for the two cowpea varieties. This therefore implies that the longer the days the plant takes to senesces, the greater the yield. From the findings of this study, the following can be recommended:
1. The use of liquid N fertilizer could be recommended to farmers to improve growth and yield in photosensitive cowpeas.
2. N fertilizer in combination with BAP could be used to extend the reproductive period and the photosynthetic
efficiency of the plant, leading to increase in yield as well as delay senescence in the photosensitive cowpea studied.
The authors have not declared any conflict of interests.
REFERENCES
|
Adepetu JA (2013). Soil and Nigeria food security. Inaugural Lecture Series 119. Obafemi Awolowo University, Ile-Ife, Nigeria. P 19.
|
|
|
|
Ames M, Johnson WS (1980). Analysis of chemical and physical compositions of soil. A guide for the hydroponic and soil lessons. pp. 1-124.
|
|
|
|
Anonymous (2014). Vigna Germplasm: Current Status and Future Needs. A report prepared by the Vigna Crop Germplasm Committee. pp. 1-11.
|
|
|
|
Boukar O, Fatokun CA, Roberts PA, Abberton M, Huynh BL, Close TJ, Kyei-Boahen S, Higgins TJV, Ehlers JD (2015). Cowpea, In. Grain Legumes (Handbook of Plant Breeding 10), Ed. A. M. De Ron (New York, NY: Springer-Verlag). pp. 219-250.
Crossref
|
|
|
|
Boukar O, Massawe F, Muranaka S, Franco J, Maziya-Dixon B, Singh B, Fatokun C (2013). Evaluation of cowpea germplasm lines for protein and mineral concentrations in grains. Plant Genet. Res. 9(4):515-522.
Crossref
|
|
|
|
Burris RH (2014). Nitrogen nutrition. Annu. Rev. Plant Physiol. 20:301-308.
|
|
|
|
Cardoso MJ, Andrade FB, Prota AB (2015). Avaliacao agroeconomica da producao de sementes de caupi Sob irrigacao. Teresina: Embrapa-CPAMN (Communicado Tecnico, 62).
|
|
|
|
Carsky RJ, Iwuafor ENO (2013). Contribution of Soil Fertility Research and Maintenance to Improve Maize Production and Productivity in Sub-Saharan African. In. Strategy for Sustainable Maize Production in West and Central Africa, Badu-Apraku B, Fakorede MAB, Ouedraogo M and Quin FM (Eds.). International Institute for Tropical Agriculture, Cotonou, Benin Republic. pp. 3-20.
|
|
|
|
Chaudhry AB, Ogo RC (1985).The seasonal effects on the performance of ten cowpea varieties in the Kainji lake Basin of Nigeria Savannah. Trop. Grain Legume Bull. 31:9-14.
|
|
|
|
Dart PJ, Day RA, Dobereiner J (2007). Some Effects of Temperature and Composition of the Rooting Medium in Symbiotic Nitrogen Fixation in Plants Synthesis. In. Tropical Grain Legume, Nutman RS (Ed.). Cambridge University Press. pp. 361-383.
|
|
|
|
Ebaid RA, Ghanem SA (2000). Productivity of Giza 177 rice variety grown after different winter crops. Egypt. J. Agric. Res. 78:717-731.
|
|
|
|
El-Batal MA, Abdel-Gawad FH, Fatima A, Abdo YMY (2004). Application as antilodging for rice plants fertilized with high nitrogen rate. Zagazig J. Agric. 31:473-490.
|
|
|
|
El-Kholy MH, Abido YMY, Nassar KE (1999). Effect of organic manure and nitrogen fertilization on efficiency of blue green algae. Inoculation in paddy fields of the international symposium on biological Nitrogen fixation and crop production,Cairo Egypt. pp. 169-179.
|
|
|
|
FAO (1999). Production Year Book. FAO, Rome.
|
|
|
|
Fatokun AC (2015). Breeding Cowpea for Resistance to Insects Pests, Attempted Crosses between Cowpea and Vigna vexillata. In. Challenges and Opportunities for Enhancing Sustainable Cowpea Production, Fatokun CA, Tarawali SA, Singh B.B, Kormawa PM, Tamo M (Eds.). International Institute for Tropical Agriculture (IITA) Ibadan, Nigeria. P 52.
|
|
|
|
Fatokun CA, Boukar O, Kamara A, Coulibaly O, Alene A, Boahen S, Young ND, Myers GO (2013). Enhancing cowpea productivity and production in drought-prone areas of Sub-Saharan Africa, In. Four Seasons of Learning and Engaging Smallholder Farmers: Progress of Phase 1, ed T. Abate (Nairobi: International Crops Research Institute for the Semi-Arid Tropics), pp. 81-112.
|
|
|
|
Ferry RL, Singh BB (1997). Cowpea genetics: a review of the recent literature, In. Advances in cowpea research, eds B. B. Singh, D. R. Mohan-Raj, K. E. Dashiell, and L. E. N. Jackai (Ibadan: Co-publication of International Institute of Tropical Agriculture (IITA); Japan Int. Research Center for Agric. Sci. (JIRCAS). Pp.13-29.
|
|
|
|
Fox RL, Kang BT (2014). Exploiting the legume-rhyzobium symbiosis in tropical agriculture. Vicent, J.M., A.S. Whitney and J. Bose (Eds.). University of Hawaii, Publication No, 145:183-210.
|
|
|
|
Gwathmey CO, Madore MA (1992). Pod removal effects on cowpea Genotypes contrasting in monocarpic senescence traits. Crop Sci. 32(4):1003-1009
Crossref
|
|
|
|
Huynh B L, Matthews W, Ehlers JD, Lucas M, Santos JP, Ndeve A et al (2016). A major QTL corresponding to the Rk locus for resistance to root-knot nematodes in cowpea (Vigna unguiculata L. Walp.). Theor. Appl. Genet. 129:87-95.
Crossref
|
|
|
|
Huynh BL, Ehlers JD, Ndeve A, Wanamaker S, Lucas MR, Close TJ, Roberts PA (2015). Genetic mapping and legume synteny of aphid resistance in African cowpea (Vigna unguiculata L. Walp.) grown in California. Mol. Breed. 35(1):36.
Crossref
|
|
|
|
Ismail AM, Hall AE (1998). Positive and potential negative effects of heat-tolerance genes in cowpea. Crop Sci. 38:381-390.
Crossref
|
|
|
|
Jones MI, Wild A (2013). Soils of West African savanna. The maintenance and improvement of their fertility. Technical Communication No 55 of the Commonwealth Bureau of Soils, Harpenden, UK. Commonwealth Agricultural Bureau (CAB), Farnham Royal, UK. P 246.
|
|
|
|
Kaul JN, Sekhon HS, Brar JS (1976). Effects of Hormones grain yield of cowpea. Trop. Grain Legume Bull. 6:3-4.
|
|
|
|
Keech O, Pesquet E, Ahad A, Askne A, Nordvall D, Vodnala SM, Tuominen H, Hurry V, Dizengremel P, Gardestrom P (2007) The different fates of mitochondria and chloroplasts during dark-induced senescence in Arabidopsis leaves. Plant Cell Environ. J. 30:1523-1534.
|
|
|
|
Kolawole GO, Tian G, Singh BB (2002). Differential Response of Cowpea Lines to Application of Phosphorus Fertilizer. In. Challenges and Opportunities of Enhancing Sustainable Cowpea Production, International Institute for Tropical Agriculture (IITA), Ibadan, Nigeria, pp. 319-328.
|
|
|
|
Leite ML, Rodrigues JD, Vigens Filho JS (2015). Avaliacao de cultivares de coupi (Vigna unguiculata (L.) Walp) quanto a productividade e compomentes de productividade, sob condicoes de estufa plastica. Rev. Agric. 72:375-385.
|
|
|
|
Leshem YY, Halevy AH, Frenkel C (1986).Free radicals and senescence. In. Processesand control of plant senescence (Eds. Leshem YY, Halevy HA, Frenkel C) Elsevier, New York, Pp. 100-166.
|
|
|
|
Maerere AP, Ishimine Y (2001). Effect of organic manure and planting pattern on growth and yield of turmeric. Afr. J. Sci. Technol. 1:14-21.
|
|
|
|
Mazid MA (1993). Exploring nitrogen remobilization for seed filling using natural variation in Arabidopsis thaliana. Ph.D dissertation.University of the Phillippines at Los Banos, 232p.
|
|
|
|
Minchin FR, Summerfield RJ, Neves MC (2014). Nitrogen nutrition of cowpea (Vigna unguiculata (L.), Walp): Effects of timing of inorganic nitrogen application. Trop. Agric. J. (Trinidad) 58:1-12.
|
|
|
|
Nooden LD (1978). Senescence and aging in plants. Sandiego, CA. Academic Press, Inc. pp. 391-439
|
|
|
|
Nour MA (1998). Effect of farmyard manure and Water regime on production of rice. Egypt. J. Agric. Res. 76:1533-1547.
|
|
|
|
Nyathi P, Campbell BM (1995). The effect of tree leaf litter, manure, inorganic fertilizer and their combination on above ground production and grain yield of maize. Afr. J. Crop Sci. 3:451-456.
|
|
|
|
Okeleye KA, Okelana MAO (2013). Effect of phosphorus fertilizer on nodulation, growth and yield of cowpea (Vigna unguiculata) varieties. Indian J. Agric. Sci. 67:10-12.
|
|
|
|
Olympios CM (1976).Effect of growth regulators on fruits set and fruit development of the eggplant (Solanum mellangena L.). Hortic. Res. 16(2):67-70.
|
|
|
|
Oritz R (1998). Cowpeas from Nigeria: A silent food revolution. Outlook Agric. 27:125-128.
Crossref
|
|
|
|
Owolade OF, Adediran JA, Akande MA, Alabi BS (2006). Effects of application of phosphorus fertilizer on brown blotch disease of cowpea. Afr. J. Biotechnol. 5:343-347.
|
|
|
|
Pruitt PD (1983). The physiological causes of senescence in annual plants. J. Biol. 279:1050-1059.
|
|
|
|
Richmond TA, Lang A (1994). The fate of infloresnece meristem is controlled by developing fruits in Arabidopsis. Plant Physiol. 106:863-876.
Crossref
|
|
|
|
Schuphan W (1974). Nutritional value of crops as influenced by organic and inorganic fertilizer treatments. Results of 12 years' experiments with vegetables (1960-1972). Qual. Plant. 23(4): 333-358.
Crossref
|
|
|
|
Seobi T (2005).Effect of farmyard manure on growth and yield of turmeric. Soil Sci. Am. J. 69:893-901.
Crossref
|
|
|
|
Singh BB (1997). Advances in Cowpea Research. Co. Publication of International Institute of Tropical Agriculture (IITA) and Japan International Research Center for Agricultural Scientist (JIRCAS), IITA, Ibadan – Nigeria. Estimating the Broad – sensitivity Heritability of early growth of cowpea. pp.79-100.
|
|
|
|
Singh BB, Mohan RD, Dashiell KE, Jackai LEN (2002). (Eds), Advances in cowpea research. IITA, JIRCAS, Devon. Pp. 326-332.
|
|
|
|
Souleymane A, Aken'Ova ME, Fatokun CA, Alabi OY (2013). Screening for resistance to cowpea aphid(Aphis craccivora Koch) in wild and cultivated cowpea (Vigna unguiculata L. Walp.) accessions. Int. J. Sci. Environ.Technol. 2:611-621.
|
|
|
|
Steele WM (1976). Cowpeas (Vigna unguiculata). In. evolution of crop plants simmonds, N.W. (Ed.) Longman London. pp. 183-185.
|
|
|
|
Subramanian JC, Kende H (1985). Ethylene action and loss of membrane integrity during senescence Tradescantia. Plant Physiol. 65:1067-1072.
|
|
|
|
Summerfield RJ, Cassman KG, Matson PA, Naylor R, Polasky S (1975). Agricultural sustainability and intensive production practices. Nature 418:671-677.
|
|
|
|
Thomas H, Schellenberg M, Vicentini F, Matile P (1996). Gregor Mendel's green and yellow pea seeds. Plant Biol. 109(1):3-4.
Crossref
|
|
|
|
Thomas H, Stoddart JL (1976). Leaf senescence: a review. Annu. Rev. Plant Physiol. 31:83-111.
Crossref
|
|
|
|
Thompson JE, Legge RL, Barber RF (1987). The role of free radicals in senescence and wounding. New Phytol. 105:317-344.
Crossref
|
|
|
|
Vanek V (2003).Farm yard manure application on rootcrops for higher yield. Plant Soil Environ. J. 49:197-202.
|
|
|
|
Wallace DH (1985). Physiological genetics of plant maturity adaptations and yield. Plant Breed. Rev. 3:21-167.
Crossref
|
|
|
|
Whalen JK (2000). The usage of chemical fertilizer, improving economy in agriculture and minimizing environmental problems. Sci. Am. J.64:962-966.
|
|
|
|
World Weather information service Lagos (2012). World Meteorological Organization.
|
|
|
|
Xio O, Li K (2006). Turmeric as a medicinal plant. Plant Prod. Sci. 6:3-89.
|