ABSTRACT
Phellinus sensu lato is a genus of polypores that are morphologically, biologically and phylogenetically highly diverse. This genus is composed of sessile and lignicolous species. Species belonging to this genus are found in all regions of the world where they decompose wood or live as tree parasites. In Burkina Faso, few studies have been conducted on this taxonomic group. Thus, these collections were carried out respectively in the classified forest of Kou (Bobo Dioulasso) and the Tin landscape (Orodara). These two sites have forest formations that provide a biotope favourable to the development of polypores. Data collection was carried out randomly along the 200 to 300 meter long transects. Basidiomes were collected from the trunk, branches or roots of forest trees using a machete. The geographic coordinates as well as the morphological characteristics of each sample were carefully noted in the field. Anatomo-morphological and ecological studies permitted to identify three (03) species. They are Phellinus cf. igniarius, Phellinus cf. leavigatus and Phellinus cf. robustus. All these species are perennial, tough and have a woody consistency. These species were collected for the first time on Parkia biglobosa, and Anogeissus leiocarpus in Burkina Faso.
Key words: Phellinus, polypores, ecology, morphology, Burkina Faso.
Hymenochaetaceae constitute a family of polypores belonging to the order Hymenochaetales. This group is highly diverse morphologically, biologically and phylogenetically (Ryvarden, 1991; Hibbett and Thorn, 2001). Species of the Hymenochaetaceae family arefungi responsible for white rot in wood ; they are either saprotrophs, facultative or obligate parasites of trees and most taxa are of tropical distribution (Hawksworth et al., 1995; Ryvarden, 1991; Ryvarden and Gilbertson, 1993). According to Ryvarden (1991), the Hymenochaetaceae constitutes a taxonomic family of polypores comprising nine genera namely ; Coltricia, Phellinus, Inonotus, Phylloporia, Pyrrhoderma, Coltriciella, Hymenochaete and Aurificaria. The first one, growing on roots are usually stipulated, while the following ones growing on trunks and branches are dimidiate to resupinated. The polypores belonging to this family are characterized by the presence or not in their hymenium or in their frame of very distinct cystids also called pointed bristles with thick wall, yellow or brown not encrusted (Wagner and Fischer, 2002).
These polypores develop on tree trunks and branches (Patouillard, 1900). They are characterized by resupinated, sessile Basidiome responsible for wood rot diseases, leading to desiccation of a wide range of tree species (Van der Kamp, 1991; Ryvarden and Gilbertson, 1993 ; Castello et al., 1995). The Hymenochaetaceae family contains 610 species belonging to 48 genera (Piepenbring, 2015). In this family, the two most famous genera are the genera Phellinus and Inonotus. At the generic level, especially in Phellinus species which represents more than half of the total number of Hymenochaetales species, the traditional systematics based on anatomo-morphological analyses of this group is imprecise or even incomplete (Corner, 1991). This genus includes several species complexes and is generally considered to be a polyphyletic group (Ryvarden, 1991; Fisher, 1996). The number of species in the genus Phellinus in the broad sense was estimated at around 154 species (Larsen and Cobb-Poulle, 1990). But this genus now has more than 180 known species (Piepenbring, 2015). The discrimination between Inonotus and Phellinus, initially based on the spore color and then on the hardness and durability of the carpophore, now rests on the structure of the hypha system; dimitic in Phellinus with generative septate and non-curly hyphae but generally monomitic in Inonotus with generative curly hyphae.
Species of the genus Phellinus are found in all parts of the world where they break down hardwood or live parasitically on trees (Piepenbring, 2015). They grow on the roots or on the trunks of trees up to a height of 10 m and the weight of the basidiocarp can exceed 50 kg (Dai and Cui, 2011). However, in Burkina Faso, limited data exist on polypores. The first inventories were carried out in 2005, 2013 and 2017 respectively (Guissou, 2005; Bicaba, 2013 ; Nankoné, 2017). These inventories made it possible to collect several specimens belonging to the genus Phellinus sensu lato all over the country. But the identification of the different species remains incomplete. The present study was carried out in the Tin landscape dominated by Parkia biglobosa (Jacq.) Benth and Mangifera indica (Linn.) and in the Kou classified forest in the part dominated by Anogeissus leiocarpus (DC) Guill. and Perr., in the western part of Burkina Faso. This study aims to contribute to the knowledge of species of Phellinus genus in Burkina Faso. This is a pioneering study because it constitutes the first anatomo-morphological characterization of the genus Phellinus sensu-lato in natural forests in western Burkina Faso.
Sample collection sites
The work of collecting polypores took place in the passage of Tin and in the classified forest of Kou.
The landscape of Tin (Figure 1) is located 12 km from Orodara on the Orodara-Djigouèra axis. This site does not appear to be the subject of scientific study, hence the lack of published scientific data. This landscape is located between latitudes 11° 04'-11°06'N and longitudes 04° 55-04°58'W. The average annual rainfall varies from 900 to 1100 mm. The vegetation is dominated by wooded and wooded savannas. Forest formations are for the most part linked to the presence of permanent to semi-permanent watercourses, the banks of which are severely degraded by human activity. The most frequent woody species were: Parkia biglobosa, Khaya senegalensis, Acacia albida, but also orchards of Mangifera indica, Anacardium occidentale, etc. The richness and diversity of the Tin landscape in woody species offer chances of finding polypores which are subservient there.
On the other hand, the classified forest of Kou located 15 km North-west of the city of Bobo-Dioulasso covers an area of ​​114 ha. The term Kou refers to the river that flows through the forest. The Kou forest presents an essentially flat relief. It belongs to the Sudanese phytogeographic domain (Guinko, 1984) ; it has an important character due to its floristic diversity. The forest inventory carried out by the PAFDK (Coulibaly, 2003) reveals nearly 200 species distributed in the different formations, are mainly savannas and gallery forests.
Sampling
The sampling was carried out following a modified version of the methodology used by Balezi (2013) which consists of collecting data along transects, inside plots and collecting along road axes. To do this, investigations were carried out along the axes leading to the sites. Prospecting and collecting missions were carried out from July to September between 2016 and 2019. At each site, samples were collected along the transects over 200 to 300 meters. Basidiomata belonging to the genus Phellinus were collected from forest species or wood with a knife (Small basidiome) or with a machete (large and leathery basidiome). The characteristics of each basidiome were noted. The health status of each host plant as well as the stage of decomposition of the substrate (wood) was also noted. Then, the geographical coordinates of the place of harvest were taken using the Global Positioning System (GPS) and a photograph of the basidiome on the substrate was taken. The samples are carefully stored in a harvest basket after wrapping in aluminum foil. A technical photograph was taken on the base camp on each sample bearing a label. The description of the macroscopic characters consisted of describing the morphological characters of the basidiome in the fresh state. It took into account the mode of growth, the habit, the shape, the consistency, the margin of the carpophore, the coating of the carpophore as well as its dimensions (diameter, thickness, projection), the characteristics of the hymeneal surface (tubes, lamellae, prickles and pores).
This description was made according to the description sheet for macrofungi from De Kesel et al. (2002) which has been simplified and adapted for the description of polypores. The characters described allow a first discrimination between the genera. The specimens collected in the field were dried using an electric desiccator (Dorrex brand) for 24 h to serve as a database for further studies. The dried basidiomata were placed in hermetically sealed mini-grip bags. Each dried sample was stored in a cabinet according to the fungal genus to which it belongs. The microscopic studies were carried out on exsiccata. An optical microscope equipped with a drawing tube of the brand NIKKON H 550 S was used for this purpose. Different cutting techniques have been used depending on the type of carpophore. A longitudinal cut was made at the level of the hymenium or at the level of the weft for the search for bristles and hyphae and basidia. A scraping of the hymeneal surface was done for basidiospores.
The sections obtained were placed in a drop of 10% KOH in order to re-swell the structures and finally to check the coloring reaction. Congo Red Ammoniacal in 1% dilute ammonia solution was used to stain the cell walls. A few drops of Melzer's reagent were used to check for spore amyloidy. Anatomical structures (connective hyphae, skeletal hyphae, hymenial and / or weft setae, basidiospores, etc.), were drawn. All the anatomical structures drawn were measured using a micrometer (µm) ruler. Thus, the length (L) and width (W) of the basidiospores, setae, and basidia were measured. From measurements of the length and width of the basidiospores, the ratio (Q) of the spore (Q = L / l) was calculated using an Excel spreadsheet, thus highlighting the differences in measurements. This made it possible to determine the shape of the basidiospore, setae, cystidia, and basidia. The drawings produced were enlarged on A3 size paper and then traced on scalp paper on which the cystidia, basidiospores, basidia and the hyphal system were carefully grouped. Each scalp paper was scanned and stored in a photographic format in JPG format then the images obtained were processed to make them clearer.
Anatomo-morphological description
Phellinus cf. igniarius
Phellinus igniarius (L. Fr.) Quél. 1886, Ench. Fung., P.172
Synonyms : Polyporus igniarius, Boletus igniarius, Fomes igniarius, Phellinus igniarius
Morphological structures
Perennial basidiome is crusted, robust and ungulate with a more less smooth appearance. The old part is black, cracked in places and the rest of the basidiome is greyish, with woody pulpit and gray-orange color turning black in contact with KOH. The lining of the basidiome is corky and thin. The margin of the basidiome is circular and well delimited by a greyish band upwards and whitish towards the hymenium. Hymenium is porous with fine, tight pores. The hymeneal surface is rusty ocher to yellowish in color. The diameter of the basidiome is 17-20 cm, on a projection from the substrate of 10-16 cm and a thickness of 6-13 cm (Figure 2A and B)
Microscopic structure
The hypha system is dimictic, consisting of generative, compartmentalized, thin-walled hyphae and fairly thick-walled skeletal hyphae. The basidia: 25-40×10-18 µm, are cylindrical with keyed-out and a little pot-bellied. The setae: 90-65 ×15-20 µm are more or less long and have a somewhat thick wall. The bristles appear quite long and pointed at their apex under an electron microscope. The lining of the basidiome consists of thin skeletal hyphae with branched apices. Basidiospores : 8-5×6-4 µm, Q= (1.50-1.28-1.00 µm ; n= 60/2) are smooth, subglobose to globose (Figure 3a-f).
Ecology: The specimen was collected from the trunk of Anogeissus leiocarpus (African Job) at a height of about 2 meters. It is a parasitic species frequently found on this plant in the classified forest of Kou.
Material examined in Burkina Faso, Houet Province, Bobo Dioulasso. Samples no. NKS0152 (Holotype), coordinates: Latitude: 11°11'14.7''N., Longitude: 004°26'46.9''W., Altitude: 336 m. Harvested by NANKONE Samson, on 08/05/2018 in the classified forest of Kou.
This perennial species is characterized by a large, robust, ungulate and truncated, greyish basidioma with black and cracked old part. Hyaline Melzer spores are rare and small in size, turning orange to yellowish when in contact with KOH. The hymeneal setae are very long and tapered. It is a species collected for the first time in 2016 and then in 2018 on Anogeissus leiocarpus in the classified forest of Kou in western Burkina Faso.
Phellinus cf. leavigatus
Phellinus leavigatus (P. Karst.) Bourdot & Galzin 1928, Hym. France, p.624
Synonym : Polyporus leavigatus (Fr.), 1874
Morphological structure
Perennial basidioma is resupinated with a rigid flesh, brownish in color against the trunk of Parkia biglobosa at a height of nearly 2 m above the ground. The basidioma diameter is 8 cm, its basidioma thickness does not exceed 2 cm, and its spread is 16 cm2. The hymenium is porous and concolorous at the basidioma. The pores line the surface of the basidioma, with approximately 10-12 pores / cm. The hymeneal surface is smooth and the pores are oval (Figure 4).
Microscopic structures
The hypha system is dimictic made up of generative hyphae and skeletal hyphae. Generative hyphae are septate with dichotomous branching, and skeletal hyphae are thick-walled and parallel in the weft. The basidia : 8-12×19-39µm are slightly keyed and cylindrical. Hymeneal setae : are not very thick, short with a more or less pointed and forked apex. Basidiospores : 8-5×6-4µm, Q=(1.60-1.30-1.00, n= 30/1) are smooth and globular to subglobose (Figures 5 a-e).
Ecology: Perennial basidioma is collected from the trunk of Parkia biglobosa 2 m from the ground. It is responsible for the brown and white rot in wood.
Distribution: It is a cosmopolitan, pantropical species reported from Europe, Central Siberia.
Material examined: Burkina Faso. Province of Kénédougou (PTIN), no. NKS042 (holotype), coordinates : Latitude 10° 53'54.4''N, Longitude 004° 50'53.0''W and Altitude 457 m collected on 08/22/2016.
Phellinus cf. leavigatus was collected only once in 2016 in the Tin Landscape. It is a resupinated, olive-colored basidioma that is thin and less spreading. The spores are smooth globose to subglobose, becoming amyloid in Melzer. Phellinus leavigatus was collected only from Parkia biglobosa.
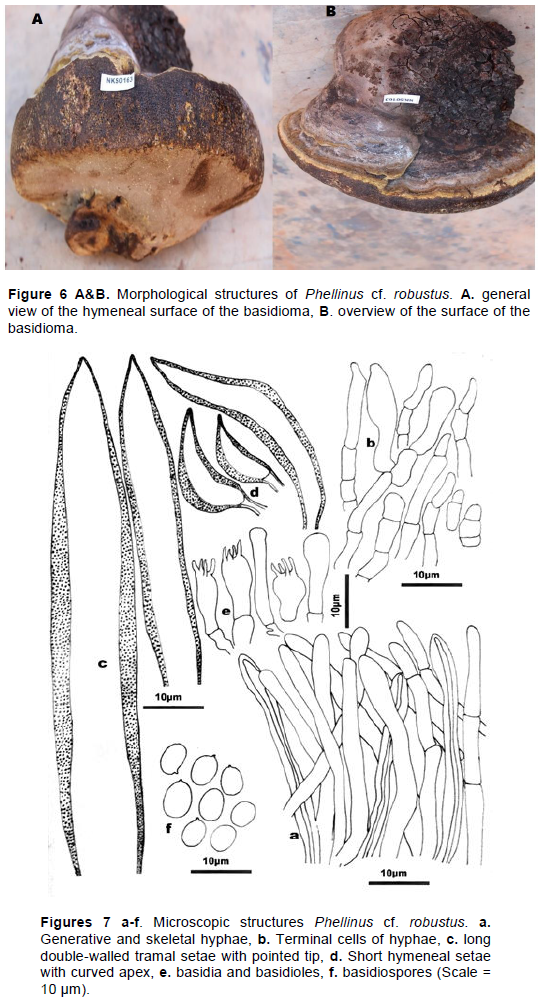
Phellinus cf. robustus
Phellinus robustus (Karsten) Bourdot and Galzin, 1925.
Bull. Soc. Mycol. France, 41 : 188.
Synonym Fomes robustus (Karst.) 1889 (Figures 6 and 7).
Morphological structures
Large, perennial basidioma, oblong, greyish in color, the old part of which is black, cracked, cracked and pre-senting bulges. The margin of the basidioma is flattened and thick with a greenish color which turns brown over time and when touched or blackens in contact with KOH. The margin secretes a limpid exudate which darkens to the touch. The porous hymenium is rusty ocher in color consisting of fine and circular pores, the tubes are woody stratified and very sclerified with a length of up to 2-5 cm. The part in contact with the substrate is covered by a clearly visible whitish to yellowish (rhizomorph) down. Its diameter is 20-30cm, its projection is 15-30 cm, and its thickness is 10-15 cm (Figure 6A and B).
Microscopic structures
The hyphae system is a dimitic hyphae system made up of generative septate hyphae, and thick-walled skeletal hyphae. Generative hyphae are quite numerous with a rounded top. The basidia : 21-15×10-5 μm are stocky with rather long sterigmas. The setae : 170-25 × 24-9 μm are numerous and have a thick double wall. The tramal setae are long while the hymeneal setae are shorter with a curved apex. The basidiospores: 8-5 × 6-5 µm, Q = (1.60-1.31-1.00; n = 60/2) are smooth, subglobose to globose (Figure 7a-e).
Ecology: Perennial fruiting bodies collected living from Parkia biglobosa.
Material reviewed: Burkina Faso. Province of Kénédougou, Landscape of Tin (PTIN), sample number NKS0163, collected on 5/08/2018, Coordinate: 11° 05'01.2''North; 004° 57'02.5'' West, Altitude 527 m.
Phellinus cf. robustus is a perennial species, very robust, ungulate with a thick margin and the old part of which is black and cracked. The basidioma is greyish in color with a brownish flesh that turns black in contact with KOH. The basidiospores are smooth, small and hyaline. This specimen has only been found on Parkia biglobosa in Burkina Faso.
Systematic
In traditional taxonomy, the genus Phellinus sensu lato differs from other genera of the Hymenochaetaceae family only by a dimictic hyphal system and the consistency of the basidioma. However, the systematics based solely on anatomo-morphological analyses of this kind is imprecise (Corner, 1991; Fiasson and Niemelä, 1984; Wagner and Fisher, 2001, 2002). Nevertheless, this study is essential and for that, it constitutes the first characterization of fungal species and which is completed by molecular analyses. Based on the anatomo-morphological characters, the genus Phellinus Quél. is defined as a group of tawny, cinnamon, rusty polypores, etc., whose caps or basidioma are in console or sometimes resupinated sessile without distinct rind, perennial. These mushrooms have a porous hymenium, with layers of tubes, often without a distinct layer of flesh. These polypores have woody, rufous, reddish-brown flesh, with a dimictic hypha system, non-curly yellowish-brown hyphae, and usually bristles present in the hymenium. The spore of these polypores is white to rusty, the spores smooth, rounded to elliptical, non-amyloid, sometimes a little dextrinoid or cyanophilic and are responsible for the white rot of the wood. Polypores of the genus Phellinus can be found all year round but are only fertile in the right season (Patouillard, 1900). The morphological characters that allowed the discrimination of species of the genus Phellinus in this study are mainly the shape of the basidioma, the dimensions of the basidioma, the hyphal system, the length of the bristles, the shape and the size of the basidiospores. Three species have been described in the context of this study: Phellinus cf. igniarius, Phellinus cf. leavigatus and Phellinus cf. robustus. These three species are all characterized by a sessile basidioma of woody consistency and have a dimictic hypha system. We noted the presence of hymeneal setae and smooth spores in all three species.
Phellinus cf. igniarius is characterized by a robust ungulate and truncated, greyish basidioma with black and cracked old part, with globose to subglobose spores. The same species, Phellinus igniarius had previously been described as having an effusive-reflexed basidioma with basidiospores (4.5-6 × 5-6.5 µm) Niemelä (1975), Fisher and Binder (2004). It should be noted that P. igniarius groups together with a polyphyletic complex of species of the genus Phellinus whose main characteristics are: a dimictic hyphae system, setae always present in the weft and in the hymenium of non-dextrinoid and hyaline basidiospores (Lamrood and Goes-Neto, 2006). In the present study, the basidiospores (8-5×6-4 µm) appear relatively larger than those previously described by Niemelä (1975), Fisher and Binder (2004). Our results did not agree with these authors. Phellinus cf. igniarius has long, tapering setae. Such a remark was made by Tomsovsky et al. (2010) who showed that P. igniarius is characterized by long and tubular hymeneal setae.
Phellinus cf. leavigatus has a resupinated, perennial basidioma of thin olive color and less spreading, perennial with basidiospores : 8-5×6-4 µm, smooth and globular. This species had been described as a resupinated species having a dimictic hypha system with basidiospores, 3-4 × 4-5 μm small and smooth (Niemelä, 1972; Ryvarden and Gilberson, 1994). According to Patouillard (1900), the spores of Phellinus leavigatus have a thick wall, smooth, hyaline, sub-globular, 4-5×3.5-4 μm. This differs from our results which present larger basidiospores. Phellinus cf. leavigatus has thin hymeneal setae with a more or less pointed and forked apex. This presents a similarity to the description made by Robert (2011) that showed that Phellinus leavigatus is marked by an absence of setae in the weft but has rather short hymeneal setae with pointed tips often split. But this differs slightly from the description made by Patouillard (1900) according to which the hymeneal setae dark brown with thick walls, swollen base, top in "halberd", short, 10-20 × 4-8 μm.
Phellinus cf. robustus is also a perennial, very robust, ungulate to ungulate-reflexed species with a thick margin and the old part of which is black and cracked. The basidiospores of Phellinus cf. robustus are smooth, small and hyaline. However, Phellinus robustus is said to have dextrinoid and sub-globular basidiospores (Rajchenberg and Wright, 1987; Decock et al., 2005). Thus these basidiospores would have a strong affinity with the spores of Phellinus elegans (Robledo et al., (2006). According to Fiasson and Niemela (1984), Phellinus robustus would have globular, cyanophilic and dextrinoid spores. Our described specimen shows setae having a thinner double wall with a pointed apex. This aspect of the silks of this species had been mentioned by Karadelev et al. (2006) who showed that the Phellinus robustus hymeneal setae are distinct with slender and elongated apices and are apparently characteristic of all species of the Phellinus robustus complex.
Ecology and distribution
The genus Phellinus contains ubiquitous lignicolous, parasitic or saprophytic species of wood and trees and causing white rots of wood (Van der Kamp, 1991; Ryvarden and Gilberson, 1993; Castello et al., 1995; Yombiyeni, 2014). They are pantropical species present in all regions of the world. But given the wide morphological variability of species of this genus, precise data on their distribution in tropical Africa remain insufficient.
In tropical Africa studies of the genus Phellinus sensu lato have been carried out and published by European mycologists. Among these researchers, we have Leif Ryvarden through publications Ryvarden and Johansen (1980), Masuka and Ryvarden (1993), Ryvarden (1998), Roberts and Ryvarden (2006) and Decock in publications Decock and Mossebo (2001, 2002), Decock et al. (2005), Decock and Bitew (2012). However, in Central Africa, this fungal genus has been mainly studied by Alphonse Balezi in Congo Balezi and Decock (2009), Balezi (2013) and Prudence Yombiyeni in Gabon (Yombiyeni et al., 2011 ; Yombiyeni, 2014). In West Africa, the only work devoted to polypores was carried out in Benin by Boris Olou, in 2020. This work comprehensively dealt with taxonomic studies based on the molecular phylogeny of polypores in tropical Africa (Olou, 2020). However, work in West Africa by local researchers has not specifically addressed the distribution of species in the genus Phellinus. The three species : Phellinus cf. igniarius, Phellinus cf. leavigatus and Phellinus cf. robustus which were the subject of this study were found in Burkina Faso on Parkia biglobosa, Mangifera indica and Anogeissus leocarpus. They are cosmopolitan species, present in both tropical and temperate climates (Ryvarden and Johansen, 1980). Phellinus cf. igniarius and Phellinus cf. robustus have been found in East Africa in two countries. Ethiopia and Tanzania Larsen and Cobb-Poulle (1990). Phellinus igniarius, Phellinus leavigatus and Phellinus robustus have been reported in Europe as parasitic species of deciduous trees such as Quercus, Betula, Castanea, Robinia, Salix, Alnus, and Carpinus etc (Karadelev et al., 2006). Phellinus leavigatus has been reported in Russia where it exhibited strong specialization on host genus Betula (Park et al., 2020).
Prospecting and collecting research of polypores in the classified forest of Kou and in the landscape of Tin permetted to collect three species of the genus Phellinus. They are: Phellinus cf. igniarius, Phellinus cf. leavigatus and Phellinus cf. robustus. These three species are all characterized by sessile basidiome with a dimictic hyphal system, with the presence of hymeneal setae and smooth spores. Phellinus cf. igniarius, a parasitic species of Anogeissus leiocarpus, is characterized by a robust ungulate and truncated, greyish basidioma with black and cracked old part. Its woody flesh is very rigid and of woody consistency. Its bristles are long, straight and tapered. However, Phellinus cf. leavigatus is a species collected from Parkia biglobosa. It has a resupinated, olive-colored basidioma that is thin and less spreading. Its basidiospores are smooth globular, becoming more or less orange on KOH. Finally, Phellinus cf. robustus has a perennial, oblong basidiome with a flattened and thick margin of greenish color turning brown with time and to the touch and bearing bulges. Its coating is greyish and turns black, cracked, with age. Its thick margin secretes a limpid exudate which turns black when touched. These three species were described for the first time in Burkina Faso. Therefore, their complete and exact identification requires molecular studies.
The authors have not declared any conflict of interests.
REFERENCES
|
Balezi A, Decock C (2009). Inonotus rwenzorianus (Basidiomycetes, Hymenochaetales) : An undescribed species from the rwenzori Mountain range. Cryptogamie Mycologie 30:225-232.
|
|
|
|
Balezi AZ (2013). Taxonomie et écologie des Hymenochaetales dans les forêts de montagne de l'Est de la République Démocratique du Congo : Cas du Parc National de Kahuzi-Biega. Thèse de doctorat, Université catholique de Louvain 174p.
|
|
|
|
|
Bicaba M (2013). Inventaire et importance des macromycètes phytopathogènes de la Forêt classée du Kou au Burkina Faso. Mémoire de D.E.A., Université de Ouagadougou, Burkina Faso. 75p.
|
|
|
|
|
Castello JD, Leopold DJ, Smallidge PJ (1995). Pathogens, patterns, process in forest ecosystems. Biosciences 1:16-24.
Crossref
|
|
|
|
|
Corner EJH (1991). Ad Polyporaceas VII. The xanthochroic polypores. Beihefte Nova Hedwigia 101:1-175.
|
|
|
|
|
Coulibaly S (2003). Résultats du traitement des données de l'inventaire forestier réalisé dans la Forêt Classée du Kou, rapport. BKF07-PAFDK, Burkina Faso.
|
|
|
|
|
Dai YC, Cui BK (2011). Fomitiporia ellipsoidea has the largest fruiting body among the fungi. Fungi Biology 115:813-814.
Crossref
|
|
|
|
|
De Kesel A, Codjia JTC, Yourou SN (2002). Guide des champignons comestibles du Bénin. Cotonou, République du Bénin, Jardin Botanique National de Belgique et Centre International d'Ecodéveloppement Intégré (CECODI). Imprimerie Coco-Multimédia. 275p.
|
|
|
|
|
Decock C, Mossebo DC (2001). Studies in Perenniporia (Basidiomycetes, Aphyllophorales): African taxa II Perenniporia centrali-aficana a new species from Cameroon. Systematic and Geography of Plants 71:607-612.
Crossref
|
|
|
|
|
Decock C, Mossebo D (2002). Studies in Perenniporia (Basidiomycetes, Polyporaceae) : African taxa III. The new species Perenniporia djaensis and some records of Perenniporia for the Dja Biosphere reserve, Cameroon. Systematic and Geography of Plants 72:55-62.
|
|
|
|
|
Decock C, Bitew A, Castillo G (2005). Fomitiporia tenuis and Fomitiporia aethiopica (Basidiomycetes, Hymenochaetales), two undescribed species from the Ethiopian highlands : taxonomy and phylogeny. Mycologia 97(1):121-129.
Crossref
|
|
|
|
|
Decock C, Bitew A (2012). Studies in Perenniporia (Basidiomycota): African taxa VI. A new species and a new record of Perenniporia from the Ethiopian Afromontain forests. Plant Ecology and Evolution 145:272-278.
Crossref
|
|
|
|
|
Fiasson JL, Niemelä T (1984). The Hymenochaetales: arevision the European poroid taxa. Karstenia 24:14-28.
Crossref
|
|
|
|
|
Fisher M (1996). On the species complexes within Phellinus : Fomitiporia revisited. Mycological Research 100:1459-1467.
Crossref
|
|
|
|
|
Fischer M, Binder M (2004). Species recognition, geography, distribution and host-pathogen relationships : a case study in a group of lignicolous basidiomycetes, Phellinus s.l. Mycologia 96:799-811.
Crossref
|
|
|
|
|
Guinko S (1984). Végétation de la Haute-Volta. Thèse de Doctoratd'Etat ès Sciences. Université de Bordeaux III. 394p.
|
|
|
|
|
Guissou KML (2005). Les macromycètes du Burkina Faso : Inventaire, ethnomycologie, valeurs nutritionnelle et thérapeutique de quelques espèces. Thèse de doctorat, Université de Ouagadougou 187p.
|
|
|
|
|
Hawksworth DL, Kirk PM, Sutton BC, Pegler DN (1995). Ainsworth and Bisby΄ s Dictionary of the Fungi. 8th edit. Wallingford: CAB International.
|
|
|
|
|
Hibbett DS, Thorn RG (2001). Basidiomycata : homobasidiomycetes. In McLaughlin DJ, McLaughlin EG, Lemke PA (eds). The Mycota VIIB: systematic and evolution. Springer, Heidelberg. pp. 121-168.
|
|
|
|
|
Karadelev M, Rusevska K, Stojanovska S (2008). Ecology and distribution of genus Phellinus (Hy¬menochaetaceae) in the Republic of Macedonia. Proceedings of the III Congress of Ecologists of the Republic of Macedonia with International Participation, Struga. Special issues of Macedonian Ecological So¬ciety 8:197-207.
|
|
|
|
|
Lamrood P, Goes-Neto A (2006). Taxonomic studies on Indian Phellinus s.l. species: Parsimony analysis using morphological characters. Mycotaxon 95:117-131.
|
|
|
|
|
Larsen MJ, Cobb-Poulle LA (1990). Phellinus (Hymenochaetaceae) A survey of the world Taxa. Synopsis Fungorum 3:1-206.
|
|
|
|
|
Masuka A, Ryvarden L (1993). Aphyllophorales (basidiomycetes) of Zimbabwe. African Journal of Mycology and Biotechnology 1:24-32.
|
|
|
|
|
Nankoné S (2017). Inventaire et enquêtes ethnomycologiques des macromycètes parasites des essences ligneuses de la forêt galerie de Dan et du paysage de Tin à l'Ouest du Burkina Faso. Mémoire de Master 2, Université Joseph KI-ZERBO. 77p.
|
|
|
|
|
Niemelä T (1972). On Fennoscandian polypores II. Phellinus leavigatus (Fr.) Bourd and Galz. And Phellinus lundellii Niemelä, n. sp. Annales Botanici Fennici 9:41-59.
|
|
|
|
|
Niemelä T (1975). On Fennoscandian polypores. IV. Phellinus igniarius, P. nigricans and P. populicola, n. sp. Annales Botanici Fennici 12:93-122.
|
|
|
|
|
Olou BA (2020). Diversity, molecular systematics, ecology and distribution of tropical African Polypores, PhD in Mycology, University of Abomey-Calavi, Benin. 187p.
|
|
|
|
|
Patouillard NT (1900). Essai taxonomique sur les familles et les genres des Hyménomycètes. Lons-le-Saunier, France. 184p.
Crossref
|
|
|
|
|
Park JH, Pavlov IN, Kim MJ, Park MS, Oh SY, Park KH, Fong JJ, Lim YW (2020). Investigating Wood Decaying Fungi Diversity in Central Siberia, Russia Using ITS Sequence Analysis and Interaction with Host Trees. Sustainability 12(6):2535.
Crossref
|
|
|
|
|
Piepenbring M (2015). Introduction to Mycology in the Tropics. APS Press, American Phytopathological Society.
Crossref
|
|
|
|
|
Rajchenberg M, Wright JE (1987). Type studies of Corticiaceae and Polyporaceae (Aphyllophorales) described by C. Spegazzini. Mycologia 79:246-264.
Crossref
|
|
|
|
|
Robert J (2011). Phellinus leavigatus (Hymenochaetaceae) de France, Fiche descriptive.
|
|
|
|
|
Roberts P, Ryvarden L (2006). Poroid fungi from Korup National Park, Cameroon. Kew Bulletin 61:55-78.
Crossref
|
|
|
|
|
Robledo G, Urcelay C, Dominguez L and Rajchenberg M (2006). Taxonomy, ecology and biogeography of polypôres (Basidiomycetes) from Argentinian polylepsis woodlands. Canadian Journal of Botanic 84:1561-1572.
Crossref
|
|
|
|
|
Ryvarden L, Gilbertson RL (1993). European polypores. Part 1.6th ed. Synopsis Fungorum. pp. 1-387.
|
|
|
|
|
Ryvarden L, Gilbertson RL (1994). European polypores 2. Meripilus-Tyromyces. Synopsis Fungorum 7. Fungiflora Oslo Norway. pp. 392-743.
|
|
|
|
|
Ryvarden L, Johansen I (1980). A preliminary polypore flora of East Africa. Funflora Oslo Norway 636p.
|
|
|
|
|
Ryvarden L (1991). Genera of polypores. Nomenclature and taxonomy. Fungiflora Oslo Norway 363p.
|
|
|
|
|
Ryvarden L (1998). African polypores-a review. Belgium Journal of Botanic 131:150-155.
|
|
|
|
|
Tomsovsky M, Vampola P, Sedlák P, Byrtusová Z and Jankovský L (2010). Delimitation of central and northern European species of the Phellinus igniarius group (Basidiomycota, Hymenochaetales) based on analysis of ITS and translation elongation factor 1 alpha DNA sequences. Mycological Progress 9:431-445.
Crossref
|
|
|
|
|
Van der Kamp BJ (1991). Pathogens as agents of diversity in forested landscapes. Forestry Chronicle 67:353-354.
Crossref
|
|
|
|
|
Wagner T, Fisher M (2001). Natural groups and a revised system for the European poroid Hymenochaetales (Basidiomycota) supported by nLSU rDNA sequence data. Mycological Research 105:773-782.
Crossref
|
|
|
|
|
Wagner T, Fisher M (2002). Proceeding towards a natural classification of the worldwide taxa Phellinus s.l. and Inonotus s.l., and phylogenetic relationships of allied genera. Mycologia 94:998-1016.
Crossref
|
|
|
|
|
Yombiyeni P, Douanla-Meli C, Amalfi M, Decock C (2011). Poroid Hymenochaetaceae from guineo-Congolian rainforest: Phellinus gabonensis sp. nov. From Gabon- taxonomy and phyllogenetic relationships. Mycological Progress 10:351-362.
Crossref
|
|
|
|
|
Yombiyeni P (2014). Contribution à l'étude de la diversité taxonomique et approche écologique des polypores en forêt guinéo-congolaise au Gabon. Thèse de Doctorat Université catholique de Louvain. 341p.
|
|