ABSTRACT
A greenhouse experiment was conducted to study the effect of inoculation of Rhizobium strain and sulphur fertilization on seed and straw yields, nutrients uptake and seedquality protein of two soybean (Glysine max L.) varieties. The experiment consisted of four levels of S (0, 20, 30 and 40 kgha-1), three Rhizobium strains (MAR-1495, SB-6-1-A2and TAL-379) and two soybean varieties (Belessa-95 and Wollo) combined factorially in complete randomized design (CRD) with three replications. Grain and straw yield and nutrients uptake increased by inoculation of Rhizobium strain whether used alone or in combination with S. Application of strain and S further increased seed and straw yield and nutrients uptake. Seed yields of Bellessa-95 inoculated with MAR-1495 at 30 and 40 kg S ha-1 were estimated to be 3864.1 and 3893.8 kg ha-1, corresponding to 112 and 114% increase; but Wollo was with seed yields of 3633.3 and 3709.2 kg ha-1, corresponding to 160 and 166% increase, respectively over control. Estimated soil N balance was maximum for S application along with inoculation of strains, ranged from -29.25 kg ha-1 in control to 80.74 kg ha-1 for Bellessa-95 and from -21.76 kg ha-1 to 84.66 kg N ha–1 for Wollo, respectively. Efficient seed nutrients uptake due to combined application further caused significant (p<0.001) increase in protein yield of soybean. With strain MAR-1495 protein yield varied from 26% in the control to 41.5% for Belessa-95 at 30 kg S ha–1, while from 28% in the control to 44% for Wollo at 40 kg S ha–1, respectively. The results clearly suggested that balanced application of S along with Rhizobium strain can affect grain and straw yield and nutrients uptake of soybean varieties, enhance their growth and improve grain quality protein as well as benefit on soil N balance in S-deficient soils.
Key words: Sulphur, strain, nutrients use efficiency, quality protein, soil nitrogen balance.
Soybean (Glycine max L.) is one of grain legumes and it is grown for its edible bean, an important source of inexpensive and high quality protein (40%) and oil (20%) around the world (Laswai et al., 2005). In Ethiopia, soybean is an important food crop widely produced in high rainfall areas, in west and southwestern parts (such as Assosa) and it is recently integrated into the cropping systems and serves as a cash crop for smallholder farmers of the area (Nigussie et al., 2009). Legumes including soybean are able to fix atmospheric N2 in association with rhizobia. In this symbiosis they partly supply their own N needs and also provide some nutrients left over to succeeding crops through decomposition of their nodule, roots and biomass (Chianu et al., 2011). The practices appeared to be very useful for smallholder farmers as it is cost effective to improve the N requirement of legumes and succeeding crops (Graham et al., 2004). Furthermore, N2 fixing soybean crop is of considerable interest for more sustainable agriculture and particularly in organic farming systems (Cazzato et al., 2012).In view of this, biological nitrogen fixation (BNF), a renewable N fertilizer source, holds great promise for smallholder farmers in sub-Saharan legume crops rank second after cereals, with their 12% contribution to national food production and occupy 18% of the total cultivated area in Ethiopia. In recent years, production of haricot bean (Phaseolus vulgaris L.) and soybean (Glycine max L.) has increased as they are exportable and cash earning commodities (Abera et al., 2013).
Declining soil fertility particularly low soil N availability is often the major factor resulting in decreased crop plant yields and recognized as a major problem to continue cereal cropping in soils of Ethiopia, specifically in soils of Assosa area (Zeleke et al., 2010). Due to low soil fertility status in the country grain legumes are generally grown in sever soil conditions which are inherently low in fertility including S and low soil pH especially in western Ethiopia (ATA, 2013). Fertility situation is further deteriorated by nutrient depletion by crops and other related processes, such as leaching and removal of crop residues in the area (Zeleke et al., 2010). The Ethiopian Soil Information Service is currently involved in mapping the entire country for all nutrients, and has found extensive areas of S, Zn, and B deficiency (Vanlauwe et al., 2015).
Sulphur is an essential nutrient for plant growth accounting to about 10% of the total N content (Anandham et al., 2007) and legume crops such as soybean generally require it in a similar quantity or more than that of phosphorus for high yield and quality (Jemal et al., 2010). But reductions in S sources from organic matter and less S returned with inadequate use of crop residues and rare addition of manure which often deplete soil organic matter (Habtemichial et al., 2007). In other words, in countries like Ethiopia where subsistence farming is practiced, the turnover of S through SOM is usually insufficient even to meet the small requirement associated with the small yields. Organic S pool which is the large proportion of soil S highly affected by long term cultivation in the tropics (Solomon et al., 2001) and further aggravated through removing plant residue. In intensive crop rotation S uptake can be very high, especially when the crop residue is removed from field along with the product (Fismes et al., 2002). In addition, less S returned with inadequate use of crop residues and rare addition of manure which often deplete soil organic matter in Ethiopia (Habtemichial et al., 2007).
In addition, using of S-free mineral fertilizers is decreasing soil S levels and threatens the adequate fertilization of most crops (Khalid et al., 2011). Application of N containing fertilizers year after year on S deficient soils can make its deficiency worse because of widen N:S ratio. Mineral fertilizers that are used in Ethiopia such as di-ammonium phosphate (DAP) and urea contain no S. Despite the important roles of S in agriculture, research pertaining to its status in soils and its response in crops are almost nonexistent in Ethiopia (Habtemichial et al., 2007). Sulphur fertilization of soil has significant potential of increasing the amount of N fixed by legumes and their grain yield, thus improving fertility status of soil (Cazzato et al., 2012). Nitrogen fixing capacity of leguminous plants can be increased by the supply of adequate amounts of nutrients such as S. S is a vital part of the ferredoxin, an iron-S protein occurring in the chloroplasts. Ferredoxin has a significant role in NO2 and sulphate reduction, the assimilation of N by root nodule bacteria and frees living N-fixing soil bacteria (Scherer et al., 2008). A study revealed lower N accumulation and a yield reduction of legumes when S was limiting (Scherer et al., 2006) and also recognized as a limiting factor not only for crop growth and seed yield but also for quality of products (Jemal et al., 2010). Because S is a main component of the amino acids methionine, cysteine, and cystine, coenzymes, thioredoxine and sulfolipids and hence improves protein quality (Jemal et al., 2005). Sharma and Sharma (2014) also reported S-containing amino acids such as methionine and cysteine increased significantly by combined application of N and S and indicated improvement in soybean nutritional value. This is because N and S are both involved in plant protein synthesis, a process that may determines yield of crops (Habtemicheal et al., 2013). Therefore, the most important constraints to soybean crop growth may be those caused by the shortage of plant nutrients such as S and N. In addition, there are not many studies available about the effect of S in presence of seed inoculation with effective strains of Rhizobium for successful soybean cultivation in Ethiopia. Therefore, a greenhouse trial was conducted to evaluate the effect of S fertilization and inoculation of Rhizobium strain on yield and nutrients uptake of grain and straw, grain quality protein content and estimated soil N Soybean (Glycine max L.) is one of grain legumes and it is grown for its edible bean, an important source of inexpensive and high quality protein (40%) and oil (20%) around the world (Laswai et al., 2005). In Ethiopia, soybean is an important food crop widely produced in high rainfall areas, in west and southwestern parts (such as Assosa) and it is recently integrated into the cropping systems and serves as a cash crop for smallholder farmers of the area (Nigussie et al., 2009). Legumes including soybean are able to fix atmospheric N2 in association with rhizobia. In this symbiosis they partly supply their own N needs and also provide some nutrients left over to succeeding crops through decomposition of their nodule, roots and biomass (Chianu et al., 2011). The practices appeared to be very useful for smallholder farmers as it is cost effective to improve the N requirement of legumes and succeeding crops (Graham et al., 2004). Furthermore, N2 fixing soybean crop is of considerable interest for more sustainable agriculture and particularly in organic farming systems (Cazzato et al., 2012).In view of this, biological nitrogen fixation (BNF), a renewable N fertilizer source, holds great promise for smallholder farmers in sub-Saharan legume crops rank second after cereals, with their 12% contribution to national food production and occupy 18% of the total cultivated area in Ethiopia. In recent years, production of haricot bean (Phaseolus vulgaris L.) and soybean (Glycine max L.) has increased as they are exportable and cash earning commodities (Abera et al., 2013).
Declining soil fertility particularly low soil N availability is often the major factor resulting in decreased crop plant yields and recognized as a major problem to continue cereal cropping in soils of Ethiopia, specifically in soils of Assosa area (Zeleke et al., 2010). Due to low soil fertility status in the country grain legumes are generally grown in sever soil conditions which are inherently low in fertility including S and low soil pH especially in western Ethiopia (ATA, 2013). Fertility situation is further deteriorated by nutrient depletion by crops and other related processes, such as leaching and removal of crop residues in the area (Zeleke et al., 2010). The Ethiopian Soil Information Service is currently involved in mapping the entire country for all nutrients, and has found extensive areas of S, Zn, and B deficiency (Vanlauwe et al., 2015).
Sulphur is an essential nutrient for plant growth accounting to about 10% of the total N content (Anandham et al., 2007) and legume crops such as soybean generally require it in a similar quantity or more than that of phosphorus for high yield and quality (Jemal et al., 2010). But reductions in S sources from organic matter and less S returned with inadequate use of crop residues and rare addition of manure which often deplete soil organic matter (Habtemichial et al., 2007). In other words, in countries like Ethiopia where subsistence farming is practiced, the turnover of S through SOM is usually insufficient even to meet the small requirement associated with the small yields. Organic S pool which is the large proportion of soil S highly affected by long term cultivation in the tropics (Solomon et al., 2001) and further aggravated through removing plant residue. In intensive crop rotation S uptake can be very high, especially when the crop residue is removed from field along with the product (Fismes et al., 2002). In addition, less S returned with inadequate use of crop residues and rare addition of manure which often deplete soil organic matter in Ethiopia (Habtemichial et al., 2007).
In addition, using of S-free mineral fertilizers is decreasing soil S levels and threatens the adequate fertilization of most crops (Khalid et al., 2011). Application of N containing fertilizers year after year on S deficient soils can make its deficiency worse because of widen N:S ratio. Mineral fertilizers that are used in Ethiopia such as di-ammonium phosphate (DAP) and urea contain no S. Despite the important roles of S in agriculture, research pertaining to its status in soils and its response in crops are almost nonexistent in Ethiopia (Habtemichial et al., 2007). Sulphur fertilization of soil has significant potential of increasing the amount of N fixed by legumes and their grain yield, thus improving fertility status of soil (Cazzato et al., 2012). Nitrogen fixing capacity of leguminous plants can be increased by the supply of adequate amounts of nutrients such as S. S is a vital part of the ferredoxin, an iron-S protein occurring in the chloroplasts. Ferredoxin has a significant role in NO2 and sulphate reduction, the assimilation of N by root nodule bacteria and frees living N-fixing soil bacteria (Scherer et al., 2008). A study revealed lower N accumulation and a yield reduction of legumes when S was limiting (Scherer et al., 2006) and also recognized as a limiting factor not only for crop growth and seed yield but also for quality of products (Jemal et al., 2010). Because S is a main component of the amino acids methionine, cysteine, and cystine, coenzymes, thioredoxine and sulfolipids and hence improves protein quality (Jemal et al., 2005). Sharma and Sharma (2014) also reported S-containing amino acids such as methionine and cysteine increased significantly by combined application of N and S and indicated improvement in soybean nutritional value. This is because N and S are both involved in plant protein synthesis, a process that may determines yield of crops (Habtemicheal et al., 2013). Therefore, the most important constraints to soybean crop growth may be those caused by the shortage of plant nutrients such as S and N. In addition, there are not many studies available about the effect of S in presence of seed inoculation with effective strains of Rhizobium for successful soybean cultivation in Ethiopia. Therefore, a greenhouse trial was conducted to evaluate the effect of S fertilization and inoculation of Rhizobium strain on yield and nutrients uptake of grain and straw, grain quality protein content and estimated soil N balance of two soybean varieties.
Description of the study area
A greenhouse experiment was conducted using Dystric Nitiosols (AsARC, 2007) at Assosa Agricultural Research Center (EIAR) to investigate the effects of integrated application of S and Rhizobium strains on grain and straw nutrients yield and grain quality protein content of two soybean varieties (Belessa-95 and Wollo). Assosa is located about 670 km west of Addis Ababa, the capital city of Ethiopia. It is a capital city of ‘Benishangul Gumuz’ Region State of Ethiopia and lies on altitude of 1,480 m above sea level, and located at 09°58’41.7” N, 034°38’09.5” E coordinates. The study soil was slity clay loam in texture with clay 33%; silt 25%; sand 42%, acidic in pH (5.30), medium in soil organic carbon (OC,1.90%), medium in soil nitrogen content (TN, 0.12%), very low in available P (14.55 ppm) and low in CEC (14.7, milliequivalents per 100 g soil).Available S was low (2.97 ppm) can be categorized under S-deficient soil.
Experimental set up
Prior to the experiment, 32 surface (0-20 cm) random sub samples were collected in a zigzag walk from different villages of Assosa district. The measurement of soil pH was performed to identify and select the experimental soil with acidic soils pH range of 5.1 to 5.5. Thirty-two farmers’ fields were considered from sampling villages with known soil acidity problems based on past and present management and production of soybean and with no previous history of Rhizobium inoculation. Surface soil samples (0-20 cm) from Megele-32 was air dried, passed through a 0.5 cm sieve and filled in 5 kg soil pots containing holes at the bottom to ensure free drainage with saucers placed under the pots to prevent losses of nutrients. The experiment consisted of four levels of S (0, 20, 30, and 40 kg ha-1 S) and three strains of Rhizobium (MAR-1495, SB-6-1-A2, and TAL-379) along with uninoculated control and two soybean varieties (Belessa-95 and Wollo) that was arranged in a Completely Randomized Design (CRD) with three replications in greenhouse conditions. For the purpose of assessing BNF, a non-N fixing reference crop (wheat, variety called Digalu) was grown in similar environmental condition with soybean.
Soil fertilization and sowing
Sulphur was applied as potassium sulfate (K2SO4) in solution form. In addition to sulphur, other nutrients, such as starter dose of N fertilizer at 18 kg N ha-1 as Urea (Solomon et al., 2012) and phosphorus fertilizer as TSP at recommended dose of 23 kg P ha-1 were applied at sowing for each treatment pots (5 kg of soil). Since sulphur fertilizer was applied as K2SO4, the disproportionate addition of K in different treatments was counter balanced by the addition of proportionate amount of potassium chloride. Basal nutrients were also added to each pot to prevent deficiency of other nutrients. These include 5 kg ha-1 Mg as MgCl2, 10 kg ha-1 Zn as ZnCl2, 1 kg ha-1 Mo as Na2MoO24.2H2O. S and other basal nutrients were dissolved in deionized water and applied to each pot in required amounts. Soils were then mixed thoroughly, and deionized water was added to raise the soil moisture to pF 2.5.
Seed treatment with Bradyrhizobuium strains
Soybean seeds were selected based on size and healthiness (able to shoot). Then the seeds were weighed and surface sterilized by soaking them first with 70% (v/v) ethanol for 10 s and 4% (v/v) sodium hypochlorite (NaOCl) solution for five minutes and later washed five times with sterilized water as indicated in Somasegaran and Hoben (1994). Each strain was applied at the rate of 10 g peat-based powder inocula per 100 g of seed. In order to ensure that all the applied inoculum stick to the seed, the required quantity of inoculants was suspended in 1:1 10% sugar solution. The sugar slurry was gently mixed with dry seed and then with Carrier-based inoculant so that all the seeds received a thin coating of the inoculant. Then Bradhyrhizobium inoculum was mixed thoroughly with these seeds. For each inoculation, separate plastic bag was used and care was taken to avoid contamination of the inoculated and uninoculated seeds. Seeds were allowed to air dry for a few minutes and were then sown at the required rate and spacing. Pots with uninoculated seeds were planted first to avoid contamination. Seeds were sown at 3-4 cm depth of soil. Five seeds per pot were sown, and plant populations was maintained by thinning at four to six leaf stages (that is, 15 days after germination) into three plants per pot and maintain plant distance of 5 cm. Soil N balance (Nba) considering the aboveground biomass (straw) was calculated by subtracting N output from N input using Equation 1 (Habtemichial et al., 2007). Roots were not removed from the soil, and hence the calculated potential N benefits are conservative estimates, as they do not include root N.
Nba= (Nf + N2 fixed) - Ng 1
Where: Nf = Applied N fertilizer (kg ha-1), Ng = N removed by soybean grain.
Chemical analysis
At physiological maturity, plants were harvested and partitioned into grain and straw and samples from each treatment were collected for analysis of N, P, K and S. Each plant part was dried in an oven at 70°C for 48 h, ground and sieved with 0.5 mm mesh for analysis of nitrogen, sulphur, phosphorus and potassium. N was determined by Micro Kjeldahl’s method (Nelson and Sommers, 1973). After samples were digested with di-acid mixture (HNO3 and HClO4), P was determined using spectrophotometric vanadium phospho-molybdate method, K using digested solution on a flame photometer and S by turbidimetric, Barium sulfate precipitation by turbidimetric, Barium sulfate precipitation method of estimating available S adapted from Motsara and Roy (2008). N, P, K and S uptakes in the grain and straw was determined quantitatively by multiplying N, P, K and S content of the seed and straw with that of seed and straw yield, respectively.
Statistical analysis
The analysis of variance was carried out using SAS statistical software version 9.00 (SAS, 2004) after parameters taken were converted into kg ha-1. Three-factor analysis of variance also were performed to evaluate the effects of treatments (Rhizobium strain, S rate and variety) and their interactions on grain and straw yield and N, P, K and S uptake. Means were separated using Tukey’s procedure (P < 0.05). A correlation analysis between grain and straw yield and nutrients uptake were also carried out.
Seed and straw yields and nutrients uptake of soybean
Seed and straw yields, and nutrient uptake (N, P, K and S) were significantly (P<0.05) varied between the two soybean varieties. The straw and seed yields and their respective N, P, K and S uptakes of soybean were significantly (P<0.001) improved by S fertilization and Rhizobium inoculation (Tables 1 and 3). Seed yield of soybean was increased by 28.8 and 25.2%, when 30 and 40 kg ha-1 S was applied as compared to control (no S application, S0) (Table 1).
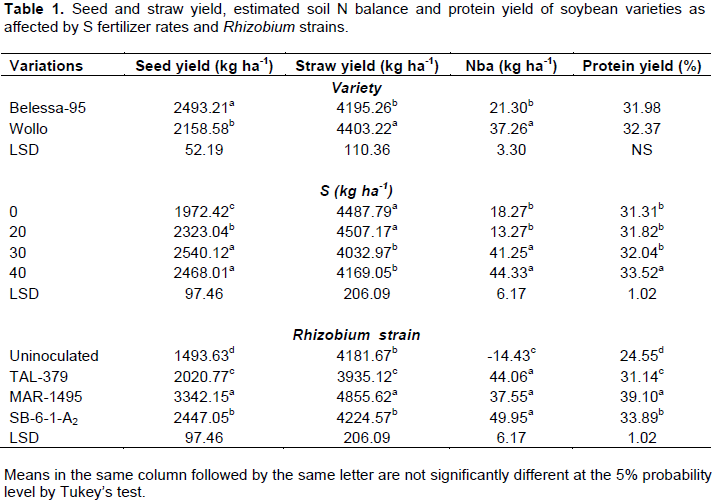
Even though most parameters were significantly (P < 0.05) affected by main effect and two-way interaction of the factors, but three-way interactions significantly (P<0.001) affected all seed and straw yields, and nutrients (N, P, K and S) uptake more than their respective two way interaction and/or main effects (Tables 2 and 4). The combined application of S (at 30 and 40 kg S ha-1) along with inoculation of MAR-1495 further showed highest seed yield for Bellessa-95 and Wollo, respectively (Table 2). For Bellessa-95, seed yields of 3864.1 and 3893.8 kg ha-1 were obtained, corresponding to 112% and 114% increases at 30 and 40 kg S ha-1, respectively with strain MAR-1495. For Wollo, seed yields of 3633.3 and 3709.2 kg ha-1 were obtained, corresponding to 160 and 166% increases for S at 30 and 40 kg ha-1, respectively with strain MAR-1495. While, S application at the rate of 30 kg S ha-1 was significantly at par with 40 kg S ha-1 for both varieties.
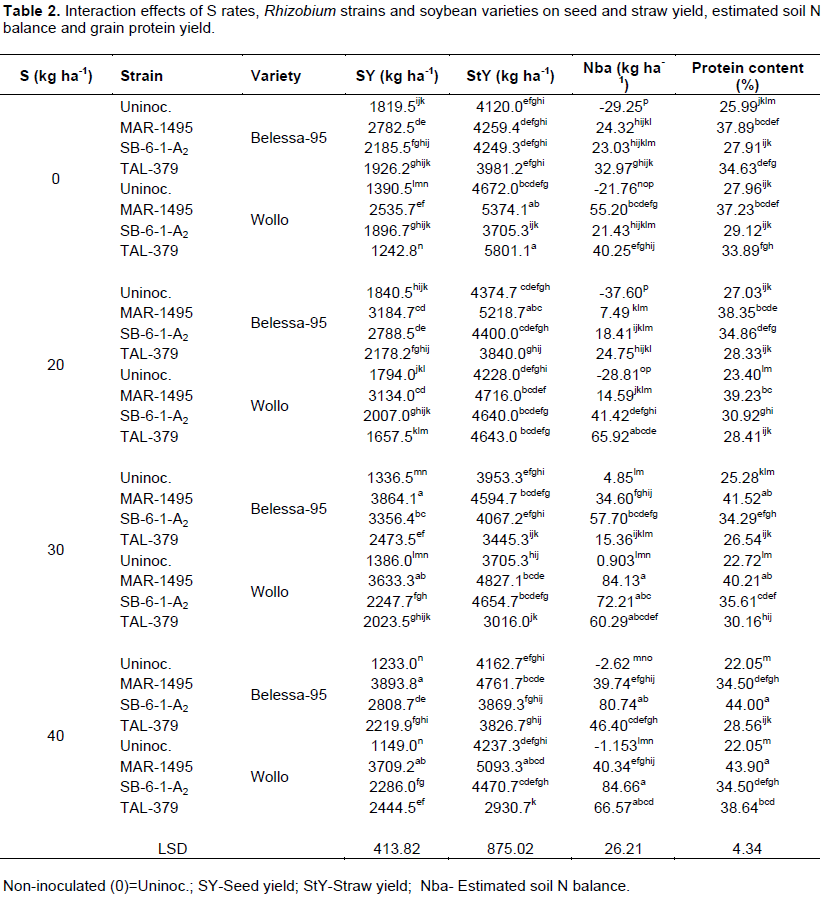
In addition, application of different doses of S increased straw yield of soybean significantly (p < 0.001) (Table 1). The highest straw yield of soybean (4507 kg ha-1) was recorded with 20 kg S ha-1 which was statistically at similar content with S0 treatment (4487 kg ha-1), whereas, the lowest straw yield were recorded in the highest S rates (30 and 40 kg S ha-1). Consequently, straw yield was higher in the inoculated treatments than uninoculated, with and/or without S application (Table 1).
N, P, K and S uptakes in soybean seed and straw were increased significantly with application of S application and inoculation of Rhizobium strains individually as well as in combination (Tables 3 and 4). The maximum increase in N uptake (3-folds) was obtained for Wollo at 40 kg S ha-1 along with inoculation of MAR-1495, while at 30 kg S ha-1 for Bellessa-95 inoculated with MAR-1495 was obtained an N uptake (256.6 kg ha-1) increased by two and half folds over control. The same is true for P and K seed uptakes which were increased significantly by addition of combined treatments of S fertilizer and inoculation of Rhizobium strain. Like N uptake, significantly the highest increase in S uptake (seventeen folds) was obtained for Wollo at 40 kg S ha-1 along with inoculation of MAR-1495, while at 30 kg S ha-1 for Bellessa-95 inoculated with MAR-1495 induced an S uptake of 39.02 kg ha-1, increased by about fifteen folds over control (Table 4).
Seed protein yield
Significant variation (p<0.001) was observed in the seed protein yield of soybean with different doses of S application, resulted increased with increasing S rates (Table 2). Application of S at 40 kg ha-1 resulted with significantly the highest protein content,ranged from 31.3% forun fertilized treatment to 33.5%, corresponding to an average increase of 7.0%. In addition, inoculation of MAR-1495 significantly increased protein content, ranged from 24.5% for uninoculted treatment to 39.1%, corresponding to an average increase by 59.2% over control. But, results showed that combined application of S with Rhizobium strain further increased protein yield for the two soybean varieties, resulted with highest protein content (Table 2). Consequently, Wollo inoculated with MAR-1495 along with S application at 40 kg ha-1 showed the highest seed protein content (43.9%) which was statistically at similar content with Wollo inoculated with SB-6-1-A2 at similar S rate (40 kg ha-1). While Belessa-95 produced higher percentage of protein when inoculated with MAR-1495 along with S at 30 kg ha-1. Whereas, the lowest protein content of soybean (27.9 and 25.9%) were recorded in the uninoculated unfertilized (R0S0) treatment combination for Wollo and Belessa-95, respectively.
Estimated soil N balance (Nba)
Nitrogen fixed by legumes represents a key contribution to nutrient cycling in legume-based farming systems by increasing soil organic N status. Soil N balance (Nba) was negative in the control treatments. Application of S had significantly increased soil N balance over the control (unfertilized) except S at 20 kg ha-1 (Table 1). On the other hand, response of inoculation of Rhizobium strain was accompanied by significant increase in estimated soil N balance compared to uninoculated control.
Inoculation of SB-6-1-A2 had showed maximum increase in N balance with corresponding variation from -14.43 kg N ha-1 (uninoculated) to 50.0 kg N ha-1. Further benefit of estimated N in soil increased with both S fertilization and inoculation Rhizobium strain for the two soybean varieties (Table 2). Deficit N balance (Nba) of soybean was lowest (-29.25 kg ha-1) in control and maximum (80.74 kg ha-1) for Bellessa-95 treated with S application at 40 kg ha-1 along with inoculation of SB-6-1-A2. For Wollo, lowest (-21.76 kg ha-1) in control and maximum (84.66 kg N ha–1) along with S application at 40 kg ha-1 and inoculation of SB-6-1-A2 (Table 2).
The results revealed that the interactive positive effect of S fertilization and inoculation of Rhizobium strain on grain and straw yield and nutrients uptake and quality protein of soybean varieties, grown as a major crop on Nitisols of Assosa area, Ethiopia. Application ofS with Rhizobium strains plays an important role in physiological and developmental processes in plant life and the favorable effect of these important nutrients combination (N and S) might accelerate the growth processes, which ultimately resulted in increased seed yield and quality of the crop. Increased in root nodulation due to Rhizobium inoculation and S fertilization resulted in absorption of higher concentration of mineral nutrients from soil and hence increased shoot dry weight by soybean. The highest nutrient in shoot was due to total N accumulation enhanced biomass yield and showed the synergistic effect of nutrients in shoot. In addition, the positive effect of S fertilization with Rhizobium strain on nitrogen fixing potential and shoot dry weight attributed to seed yield production. Rhizobium strain treatments to soybean significantly increased seed weight per plant either alone or in combination with S. Each Rhizobium strain has own synergetic effect on production of seed weight as seed weight also increased when Rhizobium strains (significantly greater for MAR-1495) were applied individually but greater in combination with S. That shows S application is important nutritional element to get better soybean yield. Similarly, results for beneficial effect of S and Rhizobium strain application on yield and yield attributing characteristics have also been recorded by other workers (Habtemichial et al., 2007; Scherer et al., 2008).
This study revealed that Rhizobium inoculation with S fertilizer increased sufficient yield of quality soybean seed. The synergistic effect of N and S may be due to utilization of high quantities of nutrients through their well-developed root system and nodules which might have resulted in better growth and yield. These results confirm the earlier findings reported by Hussain et al. (2011). The application of S might have increased the availability of nutrient to soybean plant due to improved nutritional environment, which in turn, favorably influenced the energy transformation activation of enzymes, chlorophyll synthesis as well as increased carbohydrate metabolism (Dhage et al., 2014). It constitutes the main element of amino acids such as cysteine and methionine, which are of essential nutrient value and can increase seed yield. Therefore, soybean protein quality could be significantly improved by increasing the concentration of the sulfur-containing amino acids through S fertilization. The yield and quality of legume seeds are limited by the amount of S (S) partitioned to the seeds (Tan et al., 2010). The role of S in the seed production of soybean has also been reported by (Jamal et al., 2005). Dhage et al. (2014) found that soybean seed yield and straw yield increased significantly due to application of 60 kg ha-1 S followed by 40 kg ha-1 S over control.
Each Rhizobium strain has its own synergetic effect on seed yield and nutrients uptake increased substantially when applied in combination with S. Generally, the two soybean varieties, Belessa-95 and Wollo showed highest seeds N,P, K and S uptakes when inoculated with MAR-1495 at S rate of 30 and 40 kg ha−1, respectively, indicated that balanced S application is important nutritional aspect for increasing soybean yield and nutrients uptake efficiency along with Rhizobium strain. Islam et al. (2012) also reported that S fertilization enhanced crop yield and uptake of macronutrients such as nitrogen, phosphorus and potassium especially nitrogen. According to Cazzato et al. (2012), S fertilization rates were increased nutrient levels in lupin seeds when compared with the control unfertilized treatment. Seed yield and N, P, K and S uptakes of seeds were significantly improved by Rhizobium inoculation and S fertilization. Results indicated a significant positive correlation (Table 5) between seed yield and N, P, K and S uptakes (r2=0.96, r2=0.70, r2=0.93, and r2=0.84, respectively) showing the importance of nutrients uptake on improvement in growth and seed yield of soybean. The magnitude of response on straw yield of soybean was more in case of inoculated with or without S fertilization. This may be due to the enhanced shoot nutrient uptake and N2 fixation due to combined application of Rhizobium strain with S during the vegetative growth; therefore most of the absorbed forms are re-translocated in seed than to straw during the reproductive stages. Therefore, S input modifies N allocation more to storage organs (seeds) than to straw.
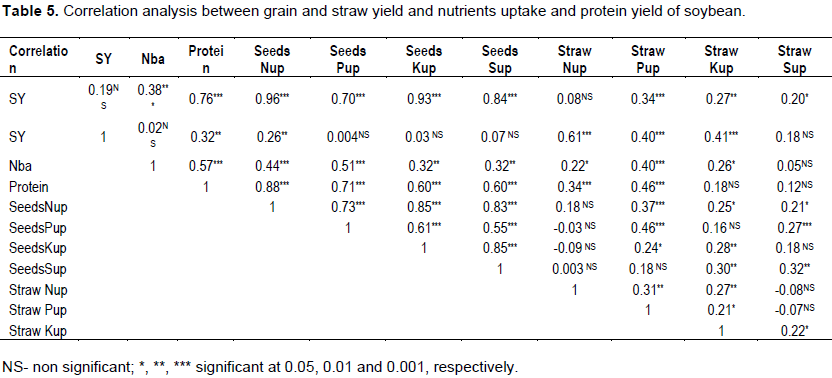
Results also revealed that Rhizobium strain alone or combined with S application increased protein content, while inoculation of strain with S increased the protein content up to 44.0%. The increased in protein content with increasing level of S may be because in the absence of S, amino acids cannot be transformed into proteins, which results in reduced N acquisition (Zuber et al., 2013). Because of central role of S and N in the synthesis of proteins, the supplies of these nutrients in plants are highly inter-related (Jamal et al., 2005), which suggested that an insufficient S supply can affect yield and quality of crops. In fact, there is close link between S supply and N requirement of plant in addition. There was accumulation of one nutrient in plant when other nutrient was lacking and accumulated nutrient was used in protein synthesis when treatments were reversed (Jamal et al.,2010).The least protein content was produced in the uninoculated plants (Tables 1 and 2); resulted with decreasing protein content with increasing S application. This may be due to nutrient imbalance between the two nutrients. Fismes et al. (2000) also found that their interactions, reflected by plant uptake, are synergistic at optimum rates and antagonistic at excessive levels of one of the both. But when applied in combination known to create a more synergism, which was helpful in improved plant growth, including nutrient uptake in the plants and the improved protein content. In legumes, previous studies showed that S deficiency decreases N assimilation and fixation (Scherer et al., 2006). In addition, modifies seed protein composition by decreasing the abundance of storage proteins with the highest content of S–containing amino acid, while increasing the level of S–poor globulins (Zuber et al., 2013). Similarly, Sharma and Sharma (2014) also reported that a significant increase in S-containing amino acids such as methionine and cysteine obtained by combined application of N and S for soybean. Therefore, the results of this study indicated that S fertilization is required to improve N-use efficiency and thereby maintaining protein quality (Fismes et al., 2000). The increase in seed nutrients uptake was correlated with the enhancement of N2 fixation due to S increased the nitrogenenase activity when applied with Rhizobium inoculation. A significant positive correlation between seed protein content and N, P, K and S uptakes of seeds (r2=0.88, r2=0.71, r2=0.60, and r2=0.60, respectively) may be due to fertilization of S facilitated the growth of plants by improving the uptake of nutrients in shoot and seeds and stimulating seeds and straw production. Accordingly, the present observations strongly support the view that seed yield, nutrients uptake and quality protein of soybean varieties are improved with combined application of Rhizobium strain and S-fertilization.
Amount of N taken by plant from soil and fertilizer was significantly higher due to inoculation and S application as compared to control. Even though the estimation only considers the chemical nature of the plant residues, it should be further estimated considering the magnitude and timing of N and S release from residues, and any subsequent immobilization. The subsequent decomposition of N rich residues replenish N removed by harvesting without the addition of fertilizer N and contribute fixed N to subsequent crops (Jensen and Hauggaard, 2003). Nitrogen inputs from grain legumes are highly dependent on the crop N harvest index, that is, the proportion of total above-ground N production removed as grain (Ravuri and Hume, 1993). Some studies show a positive effect of grain legumes on the soil N balance due to S fertilization and Rhizobium inoculation (Habtemichial et al., 2007), whereas other study show a negative effect (Hussain et al., 2011). However, most of these N balances have probably underestimated the below-ground input of fixed N by legumes due to problems of root sampling and quantifying root exudates or rhizodeposition (Khan et al., 2003). Negative N balance in the uninoculated and unfertilized control may be due to the fact that nitrogen input (starter N at 18 kg ha-1 fertilizer) was not enough to meet crop demand. Therefore, large amounts of N moved away from the soil for uninoculated as well as unfertilized control. (Amanuel et al., 2000) also reported that N balance after legume harvest is positive when crop residues are returned to soil and only seed or grain is removed, which was not included in this study. Habtemichial et al. (2007) reported positive soil N balance in range of 12 to 52 kg ha-1 after harvest of faba bean crop in Northern Ethiopia, but major difference was that crop residues were returned to soil. Higher positive soil N balance estimated with S application and inoculation might be due to the fact that amount of N fixed increased from 22.4 to 300 kg ha-1 and 28.3 to 273.2 kg ha-1 for Wollo and Belessa-95, respectively. Therefore, the estimated soil nitrogen balance became more positive, because the estimation only considers the chemical nature of the plant residues (straw nutrients content) that soybean straw can be a benefit for soil nutrients replenishment. Therefore, soybean inoculation with S fertilizer can be beneficial to farmers whose aim is to increase N balance in the soil in addition to maximize soybean yields since most of the N is removed in seeds as well as in straw harvested. Because, the amount of straw N in soybean varieties depended on how completely N was translocated to seed, total amount of N remaining in the field after soybean has been harvested for seed (Ravuri and Hume, 1993).
In this experiment the two soybean varieties performed better when inoculation of Rhizobium strain was applied with S fertilization than when both were applied alone. The combined application increased the availability and uptake of N, P, K and S by seeds and straw, and the growth and yield of the two varieties of soybean. The most beneficial effect of S with inoculation on N2 fixation, plant growth and yield can be understood given the fact that the experimental soil was severely deficient in available S as well as Rhizobium strain, which can be considered major limiting factors for quality soybean production on acidic soils of Assosa area. Hence, the integrated application of Rhizobium strain with S could be a viable strategy to improve the yield and quality of soybean, in soils containing suboptimal S and N.
The two varieties responded to the combined application of Rhizobium strain MAR-1495with S fertilizer at 30 and 40 kg ha-1. Even though with the highest yield achieved at the highest S rate (40 kg ha-1) and indicated that the yield of soybean can still be improved at further higher S rate, but were significantly at par for most parameters. Therefore, S at 30 and 40 kg ha-1 with MAR-1495was the most profitable interaction between inorganic S levels and Rhizobium strain treatments for N2 fixation, yield and quality attributes of the two soybean varieties. In addition, strain SB-6-1-A2with S fertilization also suggesting a promising way for enhancing the growth and yield of soybean. Therefore, proper fertilization programs including S integrated with inoculation of Rhizobium strain should be implemented to improve the productivity of food legumes and thereby increase total food production, enhance the supply of good quality proteins in Ethiopia.
The authors have not declared any conflict of interests.
REFERENCES
|
Agricultural Transformation Agency (ATA) (2013). Status of soil resources in Ethiopia and priorities for sustainable management. GSP for Eastern and Southern Africa. Mar 25-27, 2013, Nairobi, Kenya.
|
|
|
|
Amanuel G, Kuhne RF, Tanner DG, Vlek PLG (2000) Biological N fixation in faba bean (Vicia faba L.) in the Ethiopian highlands as affected by P fertilization and inoculation. Biol. Fertil. Soils 32:353-359.
Crossref
|
|
|
|
|
Anandham R, Sridarb R, Nalayinic P, Poonguzhalia S, Madhaiyana M, Tongmin SAA (2007) Potential for plant growth promotion in groundnut (Arachis hypogaea L.) cv. ALR-2 by co-inoculation of sulfur-oxidizing bacteria and Rhizobium. Microbiol. Res. 162:139-153.
Crossref
|
|
|
|
|
Assosa Agricultural Research Center (AsARC) (2007) Assosa Agricultural Research Center Farming system survey. Assosa, Ethioipa.
|
|
|
|
|
Cazzato E, Laudadio V, Stellacci AM, Ceci E, Tufarelli V (2012). Influence of S application on protein quality, fatty acid composition and nitrogen fixation of white lupin (Lupinus albus L.). Eur. Food. Res. Technol. 235:963-969.
Crossref
|
|
|
|
|
Chianu JN, Nkonya EM, Mairura FS, Chianu JN Akinnifesi FK (2011). Biological nitrogen fixation and socioeconomic factors for legume production in sub-Saharan Africa: a review. Agron. Sustain. Dev. 31(1):139-154.
Crossref
|
|
|
|
|
Dhage SJ, Patil VD, Patange MJ (2014). Effect of various levels of phosphorus and sulphur on yield, plant nutrient content, uptake and availability of nutrients at harvest stages of soybean [Glycine max(L.)]. Int. J. Curr. Microbiol. Appl. Sci. 3(12):833-844.
|
|
|
|
|
Fismes J, Vong, PC, Guckert A, Frossard E (2002). Influence of sulfur on apparent N-use efficiency, yield and quality of oilseed rape (Brassica napusL.) grown on a calcareous soil. Eur. J. Agron. 12:127-141.
Crossref
|
|
|
|
|
Graham PH, Tlusty B, Beyhaut E (2004). Inoculated legumes and revegetation / roadside plantings-Final Report. Department of Soil, Water, and Climate, University of Minnesota, Department of Transportation Research Services Section. pp. 1-53.
|
|
|
|
|
Habtemichial KH, Singh BR, Aune JB (2007). Wheat response to N2 fixed by faba bean (Vicia fabaL.) as affected by sulfur fertilization and rhizobial inoculation in semi-arid Northern Ethiopia. J. Plant Nutr. Soil Sci. 170:412-418.
Crossref
|
|
|
|
|
Hussain K, Islam M, Siddique MT, Hayat R, Mohsan S (2011). Soybean growth and nitrogen fixation as affected by sulfur fertilization and inoculation under rain fed conditions in Pakistan. Int. J. Agric. Biol. 13:951-955.
|
|
|
|
|
Islam M, Mohsan S, Ali S (2012). Effect of different phosphorus and sulfur levels on nitrogen fixation and uptake by chickpea (Cicer arietinumL.). Agrociencia 46:1-13.
|
|
|
|
|
Jamal A, Fazli SI, Ahmad S, Abdin MZ, Yun SJ (2005). Effect of Sulfur and Nitrogen application on Growth Characteristics, Seed and Oil Yields of Soybean Cultivars. Korean J. Crop Sci. 50(5):340-345.
|
|
|
|
|
Jamal A, Moon YS, Abdin MZ (2010). Enzyme activity assessment of peanut (Arachis hypogeaL.) under slow-release S fertilization. Aust. J. Crop Sci. 4(3):169-174.
|
|
|
|
|
Jensen ES, Hauggaard NH (2003). How can increased use of biological N2 fixation in agriculture benefit the environment? Plant Soil 252:177-186.
Crossref
|
|
|
|
|
Khalid R, Khan KS, Akram Z, Qureshi R, Gulfraz M (2011). Relationship of plant available S with soil characteristics, rainfall and yield levels of oilseed crops in Pothwar Pakistan. Pak. J. Bot. 43(6):2929-2935.
|
|
|
|
|
Laswai HS, Mpagalile JJ, Silayo VCK, Ballegu WR (2005). Use of soybeans in food formulation in Tanzania. In. Myaka FA, Kirenga G, Malema B (Eds.). Proceedings of the First National Soybean Stakeholders Workshop, 10th-11th November 2005, Morogoro, Tanzania. Pp. 52-59.
|
|
|
|
|
Motsara MR, Roy RN (2008). 'Guide to laboratory establishment for plant nutrient analysis'. Food and Agriculture Organization of the United Nations (FAO) Fertilizer and plant nutrition bulletin-19, Rome.
|
|
|
|
|
Nigussie M, Girma A, Anchala C, Kirub A (Eds.) (2009). Improved technologies and resource management for Ethiopian Agriculture. A Training Manual. RCBP-MoARD, Addis Ababa, Ethiopia.
|
|
|
|
|
Ravuri V, Hume DJ (1993) Soybean Straw Nitrogen Affected by Dinitrogen Fixation and Cultivars. Agron. J. 85:328-333.
Crossref
|
|
|
|
|
SAS Institute Inc. (2004) SAS/STAT User's Guide: Version 9.1th edition. SAS Institute Inc., Cary, North Carolina.
|
|
|
|
|
Scherer HW, Pacyna S, Spoth K, Schulz M (2006). Sulphur supply to peas (Pisum sativum L.) influences symbiotic N2 fixation. Plant Soil Environ. 52(2):72-77.
|
|
|
|
|
Scherer HW, Pacyna S, Spoth K, Schulz M (2008). Low levels of ferredoxin, ATP and leghemoglobin contribute to limited N2 fixation of peas (Pisum sativum L.) and alfalfa (Medicago sativa L.) under S deficiency conditions. Biol. Fertil. Soils 44:909-916.
Crossref
|
|
|
|
|
Sharma A, Sharma S (2014).Effect of nitrogen and S nutrition on yield parameters and protein composition in soybean [Glycine max(L.)]. J. Appl. Nat. Sci. 6(2):402-408.
|
|
|
|
|
Solomon D, Lehmann J, Tekalign M, Fritzsche F, Zech W (2001). Sulfur fractions in particle-size separates of the sub-humid Ethiopian highlands as influenced by land use changes. Geoderma 102:41-59.
Crossref
|
|
|
|
|
Solomon T, Pant LM, Angaw T (2012). Effects of Inoculation by Bradyrhizobium japonicum Strains on Nodulation, Nitrogen Fixation, and Yield of Soybean (Glycine max L.) Varieties on Nitisols of Bako, Western Ethiopia. ISRN Agronomy (261475):1-8.
|
|
|
|
|
Somasegaran P, Hoben HJ (1994). Handbook for Rhizobia–Methods in Legume-Rhizobium Technology. Springer-Verlag, Heidelberg, Germany. pp. 10-52.
Crossref
|
|
|
|
|
Tan Q, Zhang L, Grant J, Cooper P, Tegeder M (2010). Increased Phloem Transport of S-Methylmethionine Positively Affects Sulfur and Nitrogen Metabolism and Seed Development in Pea Plants. Plant Physiol. 15:1886-1896.
Crossref
|
|
|
|
|
Vanlauwe B, Descheemaeker K, Giller KE, Huising J, Merckx R, Nziguheba G, Wendt J, Zingore S (2015). Integrated soil fertility management in sub-Saharan Africa: unravelling local adaptation. Soil 1:491-508.
Crossref
|
|
|
|
|
Zeleke G, Getachew A, Dejene A, Shahidur R (2010). Fertilizer and Soil Fertility Potential in Ethiopia: Constraints and Opportunities for Enhancing the system. Working Paper, International Food Policy Research Institute. Pp. 1-20.
|
|
|
|
|
Zuber H, Poignavent G, Le Signor C, Aime D, Vieren E, Tadla C, Lugan R, Maya Belghazi M, Labas V, Santoni AL, Wipf D, Julia BJ, Avice JA, Salon C, Karine GK (2013). Legume adaptation to sulfur deficiency revealed by comparing nutrient allocation and seed traits in Medicago truncatula. Plant J. 76:982-996.
Crossref
|
|