Full Length Research Paper

ABSTRACT
Tamarindus indica (Tamarind) is a socio-economically important tree species in the Sudanian and Sahelian zone of Africa. Few studies have shown that the species is pollinated by bees. In this article, first we studied the impact of the wind pollination. Secondly, we determined the visitors insect of the flowers of tamarind, studied the production of nectar and pollen. Thirdly, we established relation between nectar production and visit time of insect. The study was undertaken in Sudanian zone of Burkina Faso. The insects were captured using a net on the flowers. The determination of the insects visiting flowers is made using the Chenery key. The results show that wind pollination of tamarind is very low. We determined two long distance pollinators of Hymenoptera group (Xylocopa olivacea and Megachille sp.) and five short distance pollinators represented by Apis mellifera and Trigona sp. (Hymenoptera group), Syrphida sp. and Bombylius sp. (Diptera group). The wasp visitor, Polistes fastidiosus (Hymenoptera group)’s role in pollination is badly established. Production of nectar and pollen at the flower level occurs over short periods. We identified two major guilds of plants: one guild of plant for A. mellifera and one for Xylocopa violacae.
Key words: Tamarindus indica, pollen, nectar, plants visitors, pollination.
INTRODUCTION
Sahelian smallholder farmers depend on many trees and shrubs, primarily indigenous species, for a range of essential products, and for environmental services that help improve food security and crop production. They are harvested by rural population for local consumption and commercialisation on a small scale as well as for supplying small industries such as manufacturers of juice (Lamien and Bayala, 1996). With increasing recognition of their importance, the fruit tree species are beginning to attract attention as renewable natural resources that are possibly under threat (Diallo et al., 2008). Managing their populations, and improving the quality and regularity of fruit production are priorities for the economic development of rural populations (Bonkoungou et al., 1998). However, the main limiting factor is the insufficiency of knowledge on the population biology of most of these locally important fruit trees. Genetic diversity at local level and population processes such as mating systems, pollination biology, seed dispersal and establishment of juveniles, are poorly studied particularly in Sahel countries. Thus, little information is available on the factors that limit fruit production, the potential for genetic improvement via selective breeding, the degree of genetic and ecological vulnerability and many other aspects important in the management of these resources. According to Loveless and Hamrick (1984), Hamrick et al. (1992), Schemske and Horwistz (1984) and Levêque (1997), the reproductive success of a population is known to be the primary factor which determines its natural dynamics and the evolution of its genetic diversity. Hamrick and Godt (1989), Hamrick and Murawski (1990) and Diallo et al. (2008) noted that the reproduction system of plants and the pollinators’ behavior play a predominant role in the genetic structure of populations.
Tamarind, Tamarindus indica L. (Leguminosae: Caesalpinioïdae) ranks fourth on the list of 15 species considered most important by rural populations in the Sudanian and Sahelian countries of Africa (Bonkoungou et al., 1998). The tree has multiple uses, including uses in traditional medicine (Tybirk, 1993). Due to the sweet and acid flavour of fruit pulp of tamarind, it is widely used for food and beverage preparation. Tamarind is an example of an economically important fruit tree to be little studied in Sahel. It is especially important in the semi-arid countries of Africa and South Asia, where it is present long time ago. Its origin remains controversial (Diallo et al., 2007).
In the Sahel countries, all trees appear to result from natural regeneration, and there is little or no management of individual trees apart from keeping them from other destructive uses (Diallo et al., 2010). Any young tree found in farms is systematically spared and allowed to grow. This dependence on natural regeneration further underscores the interest of understanding the tree’s reproductive ecology in order to maintain not only fruit production in the short term, but also for the long-term management of the tree’s populations.
Despite its economic importance, very little is known about the reproductive ecology of tamarind. In the Sahel countries, the main problem is the low fruit production of many trees. Another concern mentioned by local people is the small size of many fruits containing few seeds and hence little pulp.
Erratic and low productivity are also cited by El-Siddig et al. (1999) as limiting the scope for commercial cropping. Caesalpinioid legume trees are often self-incompatible or at least preferentially allogamous (Gibbs et al., 1999; Lewis and Gibbs, 1999; Arista et al., 1999). Although, at the tamarind, self-incompatibility is partiality and the consequences of selfing both for fruiting and seeds production are known to be low (Diallo et al., 2008).
As in other Caesalpinioid legume trees, tamarind flowers are mainly bee-pollinated (Radhamani et al., 1993; Nagarajan et al., 1997). Despite the fact that little is known about its pollinators and their behaviour on the pollination process, tamarind populations in the Sahel Country of West Africa are usually small and the individual trees are often isolated from each other. This suggests that pollinators’ insects play a fundamental role in fructification success. In zoophilous pollination systems plants and pollinators share mutual interest, each being useful to the other (Kearns et al., 1998; Herrera and Pellmyr, 2002; Dafani et al., 2005). Sahli and Conner (2007) highlighted that plant-pollinator interactions are one of the most important and reciprocal variables in nature. Plant-pollinator interactions have a significant effect on reproductive success (Janzen et al., 1980). Thus, Pesson (1984) noted that the relationships between angiosperms and their pollinators have evolved and diversified on the basis of reciprocal benefit, that is, food for the pollinators and pollen dispersal for the plants. Pollinator importance, visits rate and pollinator effective-ness are descriptive parameters of the ecology and evolution of plant-pollinator interactions (Reynolds and Fenster, 2008). The structural organization of mutualism networks, typified by inter-specific positive interactions, is important to maintain community diversity (Bartomeus et al., 2008).
Therefore, in order to understand the evolution of reproductive ecology in Tamarind populations, we must identify the pollinators and their diet to know how these intervene in pollination and in addition examine the interactions between these pollinators and their plants-hosts in the Sudano-Sahelian forest ecosystems. To generate crucial information for management of fruit production and maintenance of viable populations of this valuable and poorly studied tree, we undertook a study of Tamarind pollinators, focusing on the following: (1) what is the importance of wind in tamarind pollination (2) which insects intervene in tamarind pollination (3) Relationship between reward production by flowers for visitors and the visits time (4) if considering Tamaind to be central plant, which is the visitors guilds in the Sudanian ecosystems forest?
MATERIALS AND METHODS
Study site
Potential pollinators’ insects were collected on tamarind population. Trees were localised in the agro forestry parkland (10 ha) of Souroukoudinga (11°14’N, 4°26’W), in western Burkina Faso. The climate is Sudanian (Fontès and Guinko, 1995), that is, less arid than the Sahelian climate. There are two well-marked seasons which are: (i) the dry season which lasts approximately 5 months during which there is hot and dry wind. It includes/understands one dry and cold season and a dry and hot season; (ii) the rainy season, 4 months during which there is a wet wind called monsoon. These two great seasons are separated by two one month inter season, each one characterized by an alternation between the two types of wind. The differences between the temperatures of day labourers and seasonal are very high. The grounds ferruginous tropical are strongly washed low in nitrogen and phosphorus. The characteristics of the site are shown in the Table 1.

Field observations of potential pollinators and nectar production
In tamarind, visitors’ insects were collected with a net. The insects were sampled all day (6 to 18 h) for the successive 10 days, and then at fixed hours (6, 9, 12 , 14, 18 h) during 30 days. Pollinators were identified by the INERA entomology laboratory using the Delvare and Arbelenc (1985) key for family and Chenery key for the genus level and when possible to the species level. To determine the role of wind in tamarind pollination, we sampled 5 tamarind trees. 20 inflorescences per tree were randomly selected and 10 inflorescences were protected from the insects’ visits and the10 others are left without protection. We used the method of Goldingay et al. (1991) which consists of using a mosquito net with sufficiently small mesh to prevent insect penetration but allows ventilation. The observations were made from April 22th to May 22th. Nectar production was examined in flowers at each of the five phenological stages: (1) stage A: flower bud; (2) stage B: elongated flower; (3) stage C: open flower with closed anthers; (4) stage D: open flower with dehiscent anthers; and (5) stage E: fully opened flower at the point of wilting. For stages C, D, and E, we noted absence (0) or presence (1) of nectar within the corolla at regular time intervals (6, 9, 14, 18 h). Nectar production on tamarind flowers was estimated during 7 non rainy days on 25 flowers per stage on three trees. To determine the floral stage at which pollen viability is highest, we performed viability tests using carmine red (Kearns and Inouye, 1993; Diallo, 2001). For each floral stage, these tests were performed on 100 anthers collected on a total of 40 flowers from 3 different individuals (n = 400, that is, 100 x 40 flowers per individual). Anthers were cut with a razor blade. Sections were placed on microscope slides and pollen grains were counted and scored for viability. Cytoplasm of viable pollen grains appeared stained red whereas unviable grains appeared orange as the staining fluid simply filled up the empty cells.
Guilds constitution
Data analysis
The number of flowers that produced nectar at each stage of the flower was recorded at different hours of the day and the numbers were plotted against the observation times. The pollen production was analyzed by the Khi-deux test.
RESULTS
Impact of visit on the flowering and fruiting
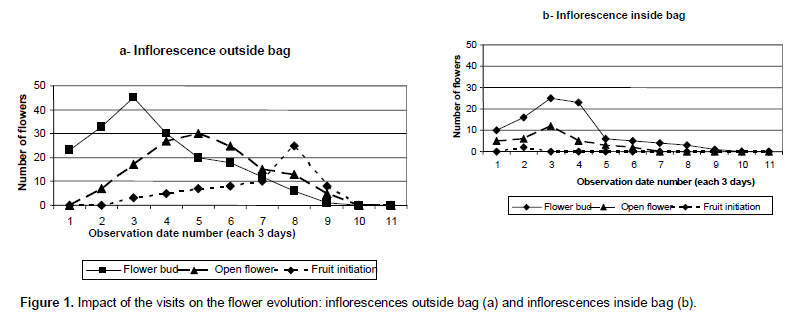
Figure 1a and b express respectively the flowering evolution on the non-protected and protected inflo-rescences for all trees. The number of closed flowers increases over a short period and decreases progressively from April 22th to May 22th both for protected branches and those put in sacks. For initiated fruits, we noticed a difference between inflorescences inside bag and outside of bags. On inflorescences inside bags, there is no initiated fruit.
Visitors/pollinators
Insects belonging to two groups were collected on tamarind flowers. Figures 2 and 3 shows six insects of T. indica flowers visitors in the study area.
Hymenoptera group
Apoidea super family composed of 4 families: (i) Apidae family: Apis mellifera (Figure 2a), subspecies Adansonii, is a honey bee belonging to the sub-family of Apinae. Trigona sp. is a small bee devoid of a stinger and belonging to the Meliponinae sub-family; (ii) Megachilidae family: Megachile sp. (Figure 2b) is a solitary bee with very vigorous flight belonging to the Megachiles sub-family. It is called "the lazy's bee" in Fulani jargon; (iii) Vespoidea family: It is a wasp belonging to the Vespidea sub-family. We found only one species, Polistes fastidiosus (Figure 2c), visiting tamarind flowers within this family; (iv) Anthophoridae family: In this family, we collected Xylocopa violacea (Figure 2d). It is a solitary bee in the Anthophorinae sub-family. They make their nest in the trees or in deadwood (carpenter bee). They are known to be pollen eaters.

Diptera group
In this group, we collected two families. (i); the family of the Syrphidae represented by two species Syrphida sp. (Figure 3a and b); (ii) the family of Bombyliidae represented by Bombilius sp. (Figure 3c).

Nectar and pollen production and insects visit periods
Nectar production
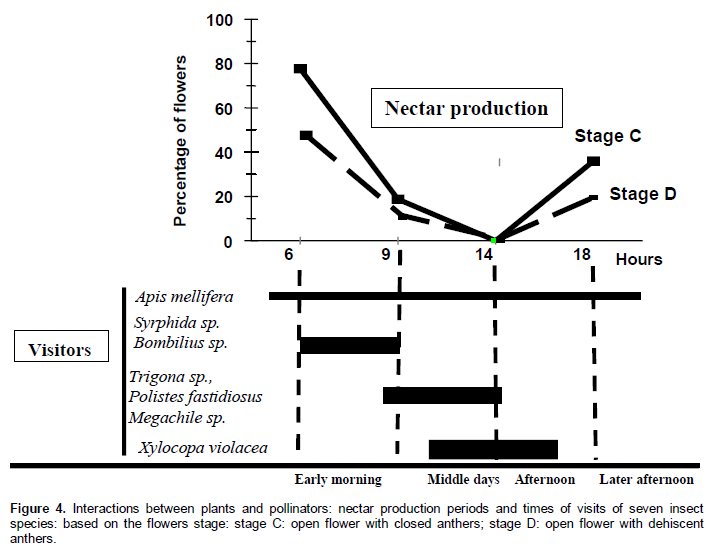
Figure 4 summarizes the periods of nectar production and insect visit time. Most of the flowers at stage C and a small number of flowers at stage D produce nectar early in the morning and later in the afternoon. During the mid-day heat, there is no nectar production during all the floral stage. The Khi-deux Test show there is no statistically significant difference (P > 0.5) between the three trees in the number of flowers that produced nectar.
Pollen production
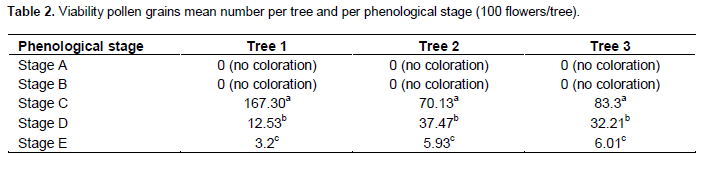
No pollen grains were observed in the anthers at the floral bud stage. At stage B and C, the many intense-red coloration of pollen grains showed that viable pollen grains were abundant (about 80% of all the grains were observed in each anther) (Figure 5).
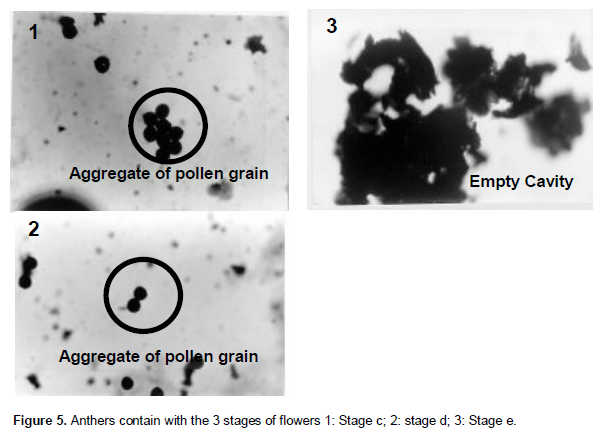
They are joined together in the shape. At stage D, they become less abundant (40%). They disap-pear entirely at stage E where the anthers are empty necrosis cavities. Table 2 shows the number of viable pollen grains for each phenological stage.
Visit time of insects
Visits occurred from 6 to 18 h and each species appeared at specific periods of time during the day. The visitors were classified into four groups based on the time of their visit: (i) the “early insects” (6-9 h) composed by A. mellifera (Hymenoptera), Syrphida sp. and Bombilius sp. (Diptera); (ii) the "second hour insects” (8-11 h) dominated by Polistes fastidiosus, Trigona sp., Megachile sp, A. mellifera; (Hymenoptera), Bombilius sp. (Diptera); (iii) the “warm hour insects” (11-15 h), characterized by the appearance of a species so far absent from the cloud of visitors, Xylocopa violacea. We noted again the presence of A. mellifera during this period. Around 15 h, P. fastidiosus, Megachile sp. and A. mellifera appeared again; (iv) the “twin light insects” (15-18 h) was dominated by A. mellifera until sunset, and then it is the only species present.
Plant and visitors guilds
Some insects have several host plants. For example, in the Sudanian eco-zones, A. mellifera adansoii, X. violacea and Megachile sp. have been captured on other plant species. However, P. fastidiosus has only been observed on T. indica during our study. Table 3 shows the insect visitors and their host plants throughout the year in Sudanian area.

DISCUSSION
Our study indicates that the wind pollination is low. This put the previous work of Oswald (1984) into perspective by which plants with pollen grains of size less than 20 µ (14-16 µ for tamarind pollen) are mainly pollinated by the wind. However, the lack of visits does not affect the subsequent evolution of flowering, but affect the fruit initiation. This shows that in the tamarind: (i) there are no cleistogamous (no fecondation before the opening of the flowers); (i) pollination by the wind is weak.
The collection and identification of insects on T. indica show that the first species group (Hymenoptera) belong to the same group as those identified in India (Rhadamani et al., 1993). This confirms partially Leppik (1956)’s observations that leguminous species are pollinated mainly by Hymenoptera group (especially bees). The second group is represented by the species of Diptera; there is completed information on the tamarind insect’s visitors, Frankies et al. (1990) also underlined the dominance of bees as pollinators in most dry tropical areas.
Pollen is available over a short period after the flower's opening. Our results are in conformity with those of Radhamani et al. (1993) which showed that the viability of tamarind-tree pollen lasts for 12 h and the difference between stage C, D and E confirm those of Stone et al. (1995) who noted that generally pollen viability decreases quickly with age of the opening flower. So, it is important that pollinators arrive at good instant to assure an efficient transfer of pollen.
The nectar is produced only during certain times of the day. Therefore, the two main resources which attract visitors to the tamarind tree (nectar and pollen) are not available throughout the entire life of the flower, and this can limit the visitors.
Only A. mellifera is present during the period of high nectar production. This insect group is known as short distance pollinators (Diallo, 2001) which are probability attracted by the deposit of honeydew. The other visitors appear when nectar production is low (Polistes, Trigona and Megachile) or nil (Xylocopa). A. mellifera is also the only species that visits the flowers throughout the day. The Polistes larva’s eats a coleopteran larva but their adults consummated pollen.
Despite the successive phenological stage of flower, the A. mellifera disperses pollen over short distances resulting in a lot of self-pollination. For self-incompatible (allogamous) plants with low tree densities (like tamarind), A. mellifera is not an efficient pollinator. This study also showed that on the Genista scorpius, honey bees rarely change trees (Diallo, 1995). Therefore, even if they visit several trees during the day (which is unlikely), only the first flowers visited receive pollen from a different tree whereas all the other flowers have a high chance of being self-pollinated. The worker bees are known to convey the distances, the quality and the food sources to each other through a dance which is a kind of language (Diallo, 2001). In contrast, X. violacea and Megachile sp. are not as common as A. mellifera but appear to disperse pollen over relatively long distances. They spend little time on any tree and can fly tens of meters, so they probably contribute to more cross pol-lination than self-pollination. The polyphagous characteristic and the vigorous fly of X. violacea allows it to reach other populations by day, thanks to "shift plants" composed by others host plants for other species trees (Da, 2003). In a study of pollination by shelter, Monty et al. (2006) showed that on 47 flowers visitors as potentials pollinators, only two species could be considered as efficient pollinator, among them is Xylocopa sp. on the basis of frequency of visits, visiting behaviour and pollen load.
By analyzing the visiting periods, we notice that the insects share the host’s resources (pollen and nectar) in a way that reduces inter-specific competition, that is, not all species visit the flowers at the same time of the day. For example, visit time of some insects species show that they are more interested in the pollen, in the sugar secreted on the flowers and leaves, or in the caterpillars that the flowers and leaves shelter. Polistes for example hunt Coleoptera larva’s or caterpillars to lay their eggs and serve as hosts for their larvae. However, the adults of these parasitic species which eat pollen are potential competitors with the species that disperse the pollen. A. mellifera is the only species that uses both pollen and nectar; but it has also diversified its supply sources by gathering pollen from several trees species. So, competition between visitors is probably low because of the diversification of resources used and the diversity of host plants.
Based on our observations, A. mellifera is the most generalist species in the Sudanian eco-zones. For this, Le Thomas (1997) noted that in the north of Ivory Coast under Sudanian south tropical climate, taxonomically close bees are attracted by the same flowers and Trigona insects seem to be the most generalist. It is necessary to note that all these species do not flourish at the same period. So, the relationship between plants and the insect visitors occurs through indirect mutualism (flowering at different periods in a year) and competition (same flowering time in a year).
CONCLUSION
Our data indicates that the wind influence is very restricted in the tamarind pollination. Tamarind potential pollinators are represented by two major groups: short pollinators and long distance pollinators. The honeybee is a short distance pollinator. Its morning arrival as well as its permanent presence disseminates hard on the auto pollen. This situation hampers the success of fruit formation according to pollinators most represented in every population of tamarind. The presence of predators (Polites) allows the diminishing rate of parasitism of fruits kernels by the Coleoptera larva. We also noted that the visits time were different in the course of the day. This shows that there is an organization to exploit the host rewards.
CONFLICT OF INTERESTS
The author(s) have declared that there is no conflict of interests.
REFERENCES
| Arista M, Ortiz PL, Talavera S (1999). Apical pattern of fruit production in raceme of Ceratonia siliqua (Leguminosae: Caesalpinioïdeae): Role of Pollinators 86(12):1708-1716. | ||||
|
Bartomeus I, Vilà M, Santamaría L (2008). Plant-Animal Interactions: Contrasting effects of invasive plants in plant–pollinator networks. Oecologia Publishe on line. Oecologia, 155:761-770. Crossref |
||||
| Bonkoungou GE, Djimdé M, Ayuk AT, Zoungrana I, Tchoundjeu Z (1998). Taking stock of agroforestry in the Sahel- harvesting results for the future. End of phase report for SALWA: 1989-1996. Nairobi, Kenya. | ||||
| Da N (2003). Fonctionnement d'une forêt soudanienne en cours d'aménagement : Cas du couple de guilde plantes/pollinisateurs. Mémoire d'ingénieur du développement rural. Université de Ouagadougou. 75p + annexes | ||||
| Dafani A, Kevan PG, Husband BC (2005). Practical pollination biology. Ecol. 67(4): 991-1001. | ||||
| Delvare G, Aberlenc HP (1989). Les insectes d'Afrique et d'Amérique tropicale; clés pour la reconnaissance des familles; 229-263. Prifas-CIRAD, Montpellier, France. | ||||
| Diallo BO (1995). Etude de la fructification et de la pollinisation chez Genista scorpius (Papilionacée). D.E.A. Evolution et Ecologie. Université Montpellier II. Science et Technique du Languedoc-Institut National de Paris-Grignon - Ecole Nationale Supérieure d'Agronomie de Montpellier. 22p. | ||||
| Diallo BO (2001). Biologie de la reproduction et Evaluation de la diversité génétique chez une légumineuse : Tamarindus indica L. (Caesalpinioideae). Thèse Univ. Montpellier II. Science et Technique du Languedoc. 119p. | ||||
|
Diallo BO, Joly HI, Mc Key D, Mc Key- Hossaert M, Chevallier MH (2010). Variation des caractères biométriques des graines et des plantules de neuf provenances de Tamarindus indica L. (Caesalpinioideae) Fruits, 65:153-167. DOI: 10.1051 Crossref |
||||
|
Diallo BO, Joly IH, Hossaert-McKey M, McKey D, Chevallier MH (2007). Genetic diversity of Tamarindus indica populations: Any clues on the origin from its current distribution? Afr. J. Biotech. 6(7):853-860. |
||||
|
Diallo BO, Mckey D, Chevallier M-H, Joly HI, Hossaert-Mckey M (2008). Breeding system and pollination biology of the semi domesticated fruit tree, Tamarindus indica L. (Leguminosae: Caesalpinioïdae): Implications for fruit production, selective breeding, and conservation of genetic resources. Afr. J. Biotech. 7(22):4068-4075. |
||||
| El-Siddig K, Ebert G, Lüdders P (1999). Tamarind (Tamarindus indica L.): a review on a multpurpose-tree with promising future in Sudan. J. Appl. Bot. Ang. Bot. 73:202-205. | ||||
| Fontès J, Guinko S (1995). Carte de la végétation et de l'occupation des sols du Burkina Faso. Note explicative, 67p. | ||||
| Frankie GW, Vinson SB, Newstrom LE, Barthell JF, Haber WA, Frankie JK (1990). Plant phenology, pollination ecology, pollinator behaviour and conservation of pollinators in Neotropical dry forest. In Reproductive ecology of tropical forest plants. Eds. K.S Bawa and M. Hadley 1990. Man and the biosphere series. UNESCO, Paris. | ||||
|
Gibbs PE, Oliveira PE, Bianchi MB (1999). Postzygotic control of selfing in Hymenaea stigonocarpa (leguminosae-caesalpinioïdeae), a batpollinated tree of Brazilian cerrados Int. J. Plants Sci. 160(1):72-78. Crossref |
||||
|
Goldingay RL, Cart SM, Whelan RJ (1991). The importance of non-flying mammals in pollination. Oikos 61:79-89. Crossref |
||||
| Hamrick JL, Godt MJW (1989). Allozyme diversity in plant species. In Plant population genetics, breeding and genetic resources, eds. A.H.D. Brown, M.T. Clegg, A. L. Kahler, and B.S. Weir : 43-63.Sinaurer, Sunderland, M.A.Hamrick J.L., and D.A. Murawski (1990). The breeding structure of tropical tree populations. Plant Spec. Biol. 5:157-165. | ||||
|
Hamrick JL, Murawski DA (1990). The breeding structure of tropical tree populations. Plant Spec. Biol. 5:157-165. Crossref |
||||
|
Hamrick JL, Godt MJ, Sherman-Broyles SL (1992). Factors influencing levels of genetic diversity in woody plant species. New For. 6:95-124. Crossref |
||||
| Herrera CM, Pellmyr O (2002). Plants animal interaction an evolutionary approach. Black Well Science. Oxford. | ||||
|
Janzen DH, Devries P, Gladstorn ED, Higgins ML, Lewinsohn TM (1980). Self and cross pollination of Encyclia cordigera (Orchidaceae) in Santa Rosa National Park, Costa Rica. Biotropica. 12:72-74. Crossref |
||||
| Kearns CA, Inouye DW (1993). Techniques for pollination biologists. Univ. Press of Colorado. USA, 583p. | ||||
|
Kearns CA, Inouye DW, Waser NM (1998). Endangered Mutualism: The conservation of plant-pollinator interactions Ann. Rev. Ecol. Syst. 29:83-112. Crossref |
||||
| Lamien N, Bayala J (1996). Rôle social et économique de l'arbre dans le milieu rural. Aspect utilisation et commercialisation de quelques produits forestiers non ligneux dans l'Ouest du Burkina. Rapport analytique de la campagne (1995-1996) ; INERA, BF, p. 28. | ||||
|
Leppik EE (1956). The form and function of numeral patterns in flowers. Am. J. Bot. 43:445-455. Crossref |
||||
| Levêque C (1997). La biodiversité. Presse universitaire de France 126p. | ||||
|
Lewis G, Gibbs P (1999). Reproductive biology of Caesalpinia calycina and C. pulviosa (leguminossae) of the Caatinga of north-castern Brazil, Plants Syst. Evol. 217(1-2):43-53. Crossref |
||||
|
Loveless MD, Hamrick JL (1984). Ecological determinants of genetic structure in plant population. Ann. Rev. Ecol. Syst. 15:65-95. Crossref |
||||
|
Monty A, Saad L, Mahy G (2006). Bimodal pollination system in rare endemic Oncocyclus irises (iridaceae) of Lebanon. Can. J. Bot. 84:1327-1338. Crossref |
||||
| Nagarajan B, Nicodemus A, Mandal AK, Verma RK, Gireesan K, Mahadevan NP (1997). Phenology and controlled pollination study in Tamarind. Silv. Genet. 47:237-241. | ||||
| Oswald H (1984). Floraison, pollinisation et fructification chez le hêtre (Fagus silvatica L.). In : Pollinisation et productions végétales. Edit. Institut National Recherche Agronomique. Grenelle, Paris. pp. 243-258 | ||||
| Pesson P (1984). Transport du pollen par les animaux : Zoogamie. In : Pollinisation et productions végétales. Edit. P. Pesson and J. Louveaux, Institut National de Recherche Agronomique, Grenelle, Paris. pp. 97-139. | ||||
| Radhamani A, Nicodemus A, Narajan B, Subramanian KN (1993). Reproductive biology and breeding system studies in Tamarindus indica L. In Pollination in Tropics. Eds. Veeresh G.K, Shaanker R.U, Ganeshaiah K.N, 33-35. International Union for the study of social insects, India Chapter. Bangalore, India. | ||||
|
Reynolds RJ, Fenster CB (2008). Plant-Animal Interactions: Point and interval estimation of pollinator importance: a study using pollination data of Silene caroliniana. Ecol. 90:2077-2087. Crossref |
||||
|
Sahli HF, Conner JK (2007). Visitation, effectiveness, and efficiency of 15 genera of visitors to wild radish, Raphanus raphanistrum (Brassicaceae). Am. J. Bot. 94(2):203-209. Crossref |
||||
| Schemske DW, Horvitz CC (1984). Variation among floral visitor in pollination ability: a precondition for mutualism specialization. Am. Ass. Adv. Sc. Vol. 225 N° 4661. | ||||
|
Stone JL, Thomson JD, Dent-Acosta SJ (1995). Assessment of pollen viability in hand-pollination experiments: review. Am. J. Bot. 82:1186-1197. Crossref |
||||
|
Tybirk K (1993). Pollination, breeding system and seed abortion in some African acacias. Bot. J. Linnean Soc. 112:107-137. Crossref |
||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


