Full Length Research Paper

ABSTRACT
The present study was carried out to study the in vitro conservation of potato genotypes at different temperatures and aerophilic and micro aerophilic conditions. A total of 31 genotypes were conserved at different incubated temperature ranging from 10 to 25°C. At lower temperature (10°C) plant growth was slowest as compared to plants incubated at high temperature (25°C). The results revealed that aerophilic condition was optimum for the growth of all potato genotypes. Data were collected on plant height, number of roots and number of nodes. Maximum plant height, highest number of roots and number of nodes were observed in all genotypes grown at 25°C. In vitro microareophillic condition of the plant growth was very slow but conservation was maximum. It was concluded from the present investigation that low temperature and micro-aerophilic condition is best for in vitro conservation of International Potato Center (CIP) germplasm which can increase the period between sub culturing.
Key words: In vitro conservation, genotype, sub-culturing, aerophilic.
INTRODUCTION
MATERIALS AND METHODS
Plant materials
A total of 31CIP potato (Solanum tuberosum L.) variety was used for in vitro conservation through different incubating temperature 10, 16 and 25°C. For that, explants (1 to 2cm) were inoculated in MS media and incubated at different temperature 25, 16 and 10°C. Enhanced growth rate was observed in plants incubated at 25°C whereas, the slow growth rates were observed in plants incubated at 10°C.
In vitro conservation
To induce a shoot from explants and to cultivate cell in suspension various kind of media have been designed. For the sake of convenience, macro and micro nutrients necessary for plants growth were formerly combined in a definite proportion to form Murashige &Skoog MS (1962) media. One of the commonly used media for tissue culture was that developed by Murashige &Skoog for tobacco tissue culture. Both the over concentrated and poor concentrated media never show satisfactory result. For in vitro conservation,simple media was used without supplement of any plant growth regulators (PGRs) and agar is also added to solidified the media which provide support into the new explant. The previously multiplied explants were used as plant materials.
In vitromultiplication was carried out by culturing nodal segment of 31 CIP different genotypes. All the equipments (Forceps, Scalpels, Petri plates) were surface sterilized in an autoclave at 121°C temperature and 15 PSI Pressure for 1h. Under aseptic condition plantlets of the CIP genotypes were taken out in a sterile plate, with the help of sterile forceps. With the help of sterile scalpel, the rootsand leaves of these plantlets were removed and finally shoot part was cut into smallsegment, each segment having at least one node. Maintaining the proper polarity of the cut segment, and inoculated in the culture medium in test tubes (size 25×190 mm, containing 10ml of solidified media). After inoculation, explants cultures were incubated at three different temperatures 10, 16 and 25°C under the light of white fluorescent tubes for 3 weeks. And for the study of aerophilic (tubes covered with plugs) and micro aerophilic (tubes covered with tight Caps) study the explants culture were incubated at the same temperature, that is, 25ËšC.
Statistical analysis
All experiments were established in a completely randomized design. Experiments showing responsive treatments were repeated once. The data were recorded the length of plants and the number of nodes and root per plants with help of ruler and then plotted on the Excel sheet for measuring the mean value andstandard error value. The data were analyzed by descriptive statistics and both the mean andstandard values were used for graphs designing.
RESULTS AND DISCUSSION
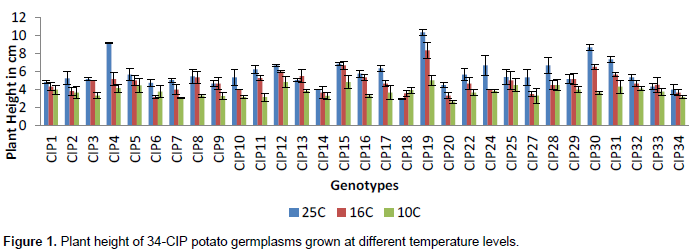

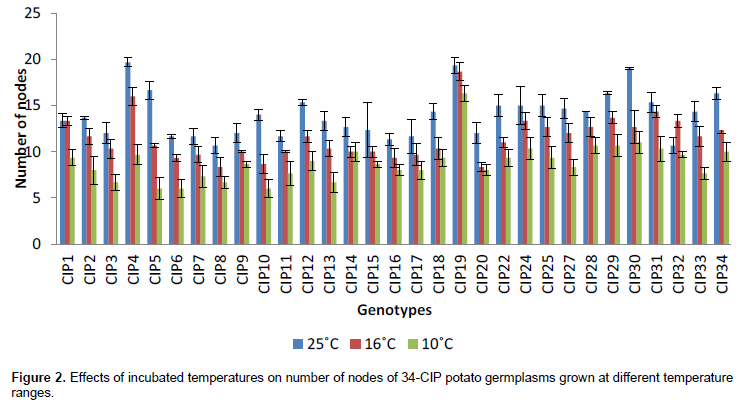


CONCLUSION
The study thus revealed that temperature of 25°C is the most favorable for multiplication of potato germplasm because at this temperature the growth rate is very fast, irrespective of the variety and time interval but for conservation of in vitro germplasm, 10°C was found optimal. Establishing the proper conditions for slow growth in potato will aid in preserving their germplasm for the purpose of later reintroduction and sustainable use. In micro-areophillic condition, the growth rate was very slow as compared to areophillic which shows its suitability for the conservation of plantlets.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interest.
REFERENCES
|
Acu-a R, Gil I, Bonachela S, Magán JJ (2008). Oxyfertigation of a greenhouse melon crop grown in rockwool slabs in a mediterranean area. Acta Hort. 779:447-454. |
|
|
Afrasiab H, Iqbal J (2010). In vitro techniques and mutagenesis for the genetic improvement of potato Cvs. Desiree and Diamant. Pak. J. Bot. 42(3):1629-1637. |
|
|
Arrigoni-Blank, Tavares FF, Blank AF, Menezes TSA, de Santana AD (2014). In Vitro Conservation of Sweet Potato Genotypes. Sci. World J. P 7. |
|
|
Badoni A, Chauhan JS (2009). Effect of growth regulators on meristem-tip development and in vitro multiplication of potato cultivar 'KufriHimalini'. Nat. Sci. 7(9):31-34. |
|
|
Bhattarai SP, Pendergast L, Midmore Nature and Sci., 7 (9): 31-34.DJ (2006). Root aeration improves yield and water use efficiency of tomato in heavy clay and saline soils. Sci. Hortic. 108: 278-288. |
|
|
Bosese SR, Huner NPA (1990). Effects of growth temperature and temperature shift on spinach leaf morphology and photosynthesis. Plant physiol. 94:1830-1836. |
|
|
Caron J, Nkongolo NV (2004). Assessing gas diffusion coefficients in growing media from in situ water flow and storage measurements. Vadose Zone J. 3:300-311. |
|
|
Chang WW, Huang L, Shen M, Webster C, Burlingame AL, Roberts JK (2000). Patterns of protein synthesis and tolerance of anoxia in root tips of maize seedlings acclimated to a low-oxygen environment, and identification of proteins by mass spectrometry. Plant Physiol. 122:295-318. |
|
|
Cherif M, Tirilly Y, Belanger RR (1997). Effect of oxygen concentration on plant growth, lipidperoxidation and receptivity of tomato roots to Pythium F under hydroponic conditions. Eur. J. Plant Pathol. 103:255-264. |
|
|
Ciobaniui IB, Constantinovici D (2012). The effects of sorbital and conservation period on the in vitro evaluation of Solanum tubersoum L. plantlets. Cercetări Agronomiceîn Moldova. 45:(3):151. |
|
|
Ciobanu IB, Constantinovici D, Creţu L (2011). Influence of genotype and cultivation conditions on vitro plantlets evolution of Solanum tuberosum L. local varieties. pp. 13-20. |
|
|
Factor TL, De Araujo JAC, Kawakami FPC, Iunck V (2007). Potato basic minitubers production in three hydroponic systems. Hortic. Bras. 25:82-87. |
|
|
Geigenberger P (2003). Response of plant metabolism to too little oxygen. Curr. Opin. Plant Biol. 6:247-256. |
|
|
Gopal J, Chamainl A, Sarker D (2002). Slow growth in vitro propagation of potato germplasm at normal propagation temperature. Potato Res. 45:203-213. |
|
|
Greenway H, Gibbs J (2003). Mechanisms of anoxia tolerance in plants. II. Energy requirements for maintenance and energy distribution to essential processes. Funct. Plant Biol. 30:999-1036. |
|
|
Klok EJ, Wilson IW, Wilson D, Chapman SC, Ewing RM, Somerville SC, Peacock WJ, Dolferus ES (2002). Dennis. Expression profile analysis of the low-oxygen response in Arabidopsis root cultures. Plant Cell 14:2481-2494. |
|
|
Kotkas, K (2004). Influence of culture medium composition on in vitro preservation of potato varieties by means of meristemplants. Analele I.C.D.S.Z., BraÅŸov 31:97-108. |
|
|
Liu F, VanToai T, Moy LP, Bock G, Linford J (2005). Quackenbush. Global transcription profiling reveals comprehensive insights into hypoxic response in Arabidopsis. Plant Physiol. 137:1115-1129. |
|
|
Maltaris T, Seufert R, Fischl F, Schaffrath M, Pollow K, Koelbl H, Dittrich R (2007). The effect of cancer treatment on female fertility and strategies for preserving fertility. Eur. J. Obstet. Gynecol. Reprod. Biol. 130(2):148-155. |
|
|
Mobini SH, Ismail MR, Arouiee H (2009). Influence of ventilation and media on potato (Solanum tuberosum L.) tuberization and its growth characteristics. Afr. J. Biotechnol. 8(10):2232-2241. |
|
|
Mohamed MAH, Alsadon AA (2009). Influence of ventilation and sucrose on growth and leaf anatomy of micropropagated potato plantlets. Sci. Hortic. 123(3):295-300. |
|
|
Morar P, Silvestre J (1996). Plant injury due to oxygen deficiency in the root environment of soilless culture: A review. Plant Soil 184:243-254. |
|
|
Murashige T, Skoog F (1962). A revised medium for rapid growth and bio assays with tobacco tissue cultures. Plant Physiol. 15:473-497. |
|
|
Nisar M, Khan N, Ahmad Z, Ghafoor A (2011). Genetic diversity and disease response of rust in bread wheat collected from Waziristan Agency, Pakistan." Int. J. Biodivers. Conserv. 3(1):10-18. |
|
|
Pitman NCA, Jorgensen PM (2002). Estimating the size of the world's threatened flora. Sci. 298:989. |
|
|
Rahman MH, Islam R, Hossain M, Islam MS (2010). Role of sucrose, glucose and maltose on conventional potato micropropagation. J. Agric. Technol. 6(4):733-739. |
|
|
Ramanatha Rao V, Riley R (1994). The use of biotechnology for conservation and utilization of plant genetic resources. Plant Genet Resour Newsl. 97:3-20. |
|
|
Rao NK (2004) Plant genetic resources: Advancing conservation and use through biotechnology. Afri. J. Biotech. 3(2):136-145. |
|
|
Ritter E, Angulo B, Riga P, Herran C, Relloso J, Sanjose M (2001).Comparison of hydroponic and aeroponic cultivation systems for the production of potato minitubers. Potato Res. 44:127-135. |
|
|
Roca WM, Espinoza NO, Roca MR, Bryan JE (1989). A tissue culture method for the rapid propagation of potatoes. Amer. Potato J. 55:691-701. |
|
|
Rolletschek H, Radchuk R, Klukas C, Schreiber F, Wobus U, Borisjuk L (2005a). Evidence of a key role for photosynthetic oxygen release in oil storage in developing soybean seeds. New Phytol. 167:777-786. |
|
|
Rolletschek H, Weber H, Borisjuk L (2003). Energy status and its control on embryogenesis of legumes. Embryo photosynthesis contributes to oxygen supply and is coupled to biosynthetic fluxes. Plant Physiol.132:1196-1203. |
|
|
Sarkar D, Naik PS (1999). Factors effecting minimal growth conservation of potato microplant in vitro. Euphytica. 102:275-280. |
|
|
Withers LA (1991). In vitro conservation. Biol. J. Linn. Soc. 43:31-42. |
|
|
Withers, Lyndsey A, Florent E (1997). In vitro conservation of plant genetic resources. Biotech. Agric. 1997:57-88. |
|
|
Yousef AAR, Suwwan MA, Musa AM, Abu-Qaoud HA (2001). In vitro culture and microtuberization of spunta potato (Solanum tuberosum). Dirasat. Agric. Sci. 24:173-181. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


