Full Length Research Paper

ABSTRACT
The Russian wheat aphid (RWA), Diuraphis noxia (Kurdjumov) causes extensive economic damage to wheat (Triticum aestivum L.) in most wheat growing regions of the world. Control of RWA using systemic insecticides is expensive and pollutes the environment therefore the most effective method of RWA control is the development of RWA resistant cultivars. This study was initiated to determine inheritance of RWA resistance in a wheat resistance source KRWA9, and identify the chromosome location of the resistance gene. Inheritance was studied in parent materials, F1 populations, F2 populations and F2:3 families of a cross between resistant line KRWA9 and a susceptible variety NjoroBW2. Seedlings were infested with RWA then scored for damage on a visual scale of 1 to 9 after 21 days of infestation. The segregation data from NjoroBW2 × KRWA9 population depicted monogenic dominant inheritance of the resistance gene with phenotypic ratios of 3:1 in F2 populations and 1:2:1 in F2:3 families. Bulk segregant analysis approach was used for the mapping of resistance. Nine simple sequence repeat (SSR) primers were tested between parental lines and bulks, and only chromosome 7DS SSR marker Xgwm111 produced clear polymorphism between the parental lines and the resistant and susceptible bulks. Detailed analysis of this marker with the full population revealed very close linkage to resistance with a coefficient of determination (R2) value of 85%. This marker provides good opportunities for the marker-assisted breeding towards improving Russian wheat aphid resistance.
Key words: Russian wheat aphid, resistance, susceptibility, simple sequence repeat (SSR) markers.
INTRODUCTION
The Russian wheat aphid (RWA), Diuraphis noxia (Kurdjumov), a pest of wheat and barley, is indigenous to southern Russia, Iran, Afghanistan and countries bordering the Mediterranean Sea (Hewitt et al., 1984). The pest has spread widely and is now found in all the continents except Australia (Ennahli et al., 2009), and causes economic damage to wheat in many parts of the world. In Ethiopia, Miller and Haile (1988) reported 68% yield loss in wheat. In South Africa, 21−92% yield losses were reported (Du Toit and Walters, 1984). In Kenya, it can cause losses of up to 90% in wheat (Malinga, 2007) and sometimes up to 100% due to prolonged drought conditions. RWA attacks the plant by infesting the young growing tip, deep in the leaf whorls where it feeds from the phloem of longitudinal veins. Symptoms of RWA attack appear as chlorotic spots that coalesce to form white, yellow or purple streaks running parallel to the mid rib of leaves (Botha and Matsiliza, 2006). In young plants, heavy infestation leads to prostate tillers while adult plants show trapped ears within the flag leaf looking like a fish hook. Severe infestation may lead to head sterility and death of host plant.
Insecticide use and particularly contact foliar applications are ineffective because of the feeding nature of the aphid. The aphid feeds within the rolled leaf whorl so cannot be easily reached by contact foliar sprays. This necessitates the use of more expensive systemic insecticides which apart from being harmful to the environment promote development of resistant biotypes and destroys biological agents. RWA resistant cultivars have been observed to have a yield advantage as compared to susceptible cultivars (Tolmay et al., 2000) and resistant cultivars have low cost as seed is usually the least expensive component in the production system besides being environment friendly. Host plant resistance is therefore, the most desirable alternative that could form part of an integrated pest management programme (IPM).
The first RWA resistant cultivar, TugelaDn (containing resistance gene Dn1), was released in South Africa in 1992 (Van Niekerk, 2001). A new biotype-designated RWASA2 was identified in 2005 virulent to Dn1, Dn2, Dn3 and Dn9 (Jankielsohn, 2011). Most of the RWA resistant cultivars available for commercial production in South Africa (Tolmay et al., 2007) were overcome by RWASA2. Similarly, resistant cultivar Halt (containing Dn4) was released in the United States in 1994 (Quick et al., 1996), but a new biotype, USARWA2 with virulence to resistance genes Dn4 and Dny was reported in 2004 (Haley et al., 2004), also overcoming the majority of commercially available resistant cultivars. Although RWA resistance expression is known to be influenced by genetic background (Randolph et al., 2005; Tolmay and Van Deventer, 2005), it is nonetheless assumed to function on a gene-for-gene basis in terms of the resistance/biotype interaction (Ricciardi et al., 2010). Recently a third biotype, RWASA3 virulent to Dn1, Dn2, Dn3, Dn4 and Dn9 was reported in South Africa by Jankielsohn (2011). Notably, neither Dn4 nor Dny had been deployed against RWA in South Africa. In Kenya, two biotypes with genetic differences have been discovered in the major wheat growing areas, that is, Njoro and Timau (Malinga et al., 2007a). Amplified fragment length polymorphism markers used to detect genetic differences showed that the Njoro biotype may contain more virulent populations as compared to Timau biotype (Malinga et al., 2007a). This was the first confirmatory report on biotypes in Kenya and it raised great challenges to resistance breeding programs for Russian wheat aphid.
Breeding for RWA resistant cultivars requires a reliable method of selecting plants containing a resistant gene. While phenotype based selection method is straight-forward, it has several limitations like the environmental influence on symptoms of damage expression. It is therefore highly desirable to employ a screening technique that is based on molecular markers linked to the resistance genes. Aside from overcoming the problems associated with phenotypic screening, marker-assisted selection (MAS) would enable gene pyramiding which is the combination of two or more resistance genes efficiently. This will expedite the process of breeding for multiple and durable resistance.
Most of the known wheat genes conferring resistance to RWA, have been mapped using microsatellite markers. Nine of these resistance genes are located on the D genome of wheat and one on the 1RS/1BL translocation (McIntosh et al., 2003). A study by Liu et al. (2001) revealed that the locus for wheat microsatellite GWM111 (Xgwm111), located on wheat chromosome 7DS (short arm), is tightly linked to RWA resistance genes Dn1, Dn2 and Dn5, as well as Dnx in wheat resistance source PI 220127. The segregation data indicated that RWA resistance in PI 220127 is also conferred by a single dominant resistance gene (Dnx) (Liu et al., 2001). These results by Liu et al. (2001) confirmed that Dn1, Dn2 and Dn5 are tightly linked to each other, and this provided new information about their location, being 7DS, near the centromere, instead of as previously reported on 7DL. According to Miller et al. (2001), the marker Xgwm437 is closely linked to Dn2 at 2.8cM. Xgwm106 and Xgwm337 flanked Dn4 on chromosome 1DS at 7.4 and 12.9 cM, respectively (Liu et al., 2002). Nkongolo et al. (1991a) reported RWA resistance gene dn3 in Triticum tauschii. Dn5 is located on wheat chromosome 7DS rather than 7DL and microsatellite marker Xgwm635 shows close linkage to the gene (Liu et al., 2001). The markers Xgwm44 and Xgwm111 are linked to Dn6 near the centromere on chromosome 7DS at 14.6 and 3.0 cM, respectively (Liu et al., 2002). This was the first report of the chromosome location of Dn6, which is either allelic or tightly linked to Dn1, Dn2, Dn5 and Dnx. Xgwm635 (near the distal end of 7DS) clearly marked the location of a previously suggested resistance gene in PI 294994, which was designated as Dn8 (Liu et al., 2001). Xgwm642, in a defense gene-rich region of chromosome 1DL, marked another new gene Dn9 from PI 294994 (Liu et al., 2001). A third new gene Dny from the Chinese wheat Lin-Yuan207 was localized on chromosome 1DL between Xgwm111 and Xgwm337 (Liu et al., 2001). A study was carried out with PCR markers for Russian Wheat Aphid Resistance Gene Dn7 on Chromosome 1RS/1BL and two markers which amplified rye-specific fragments proved to be useful for MAS. Xrems1303 amplified a 320-bp band only in cultivars with high-level resistance to USA biotype 2 and was effective for MAS of Dn7. Xib267 was found to be linked to the susceptible locus and amplified a fragment specific for rye Petkus 1RS (Lapitan et al., 2007).
Most of the Kenyan commercial wheat varieties are susceptible to RWA (Kiplagat, 2005) and since breeding of RWA resistant cultivars is further complicated due to presence of RWA biotypes, rapid breeding for and deployment of additional wheat cultivars resistant to RWA is urgently needed to reduce further losses from RWA outbreaks. This study was carried out to determine the inheritance and chromosome location of RWA resistance gene in the wheat source KRWA9.
MATERIALS AND METHODS
Plant materials and population development
Seeds were obtained from the Kenya Agricultural Research Institute, Njoro and planted in the crossing block in a row spacing of 30 cm. Crossing was carried out between resistant line ‘KRWA9’ and susceptible commercial variety ‘Njoro BW2’ to obtain F1 progeny. The F1 progeny was planted the following season and selfed to obtain F2 seeds. F2:3 families were obtained by planting seeds harvested from individual F2 plants. Plants grew under normal rainfall regime with occasional irrigation supplement.
Phenotyping
The parents, F1 plants, 100 F2 plants and F2:3 families were screened for RWA resistance under greenhouse conditions. Parents, F1 and F2 seedlings were grown in 20-cm-diameter pots containing sterilized forest soil and sand at a ratio of 3:1 mixed with 5 g Di-ammonium phosphate (18-46-0) fertilizer. Each pot contained two to four seedlings. Fifteen (15) seeds from each F2:3 family were planted on evaluation flats (1.5 × 1.0 × 0.75 m) containing sterilized forest soil and sand at a ratio of 3:1 mixed with 75 g Di-ammonium phosphate (18-46-0) fertilizer. Due to poor germination, screening data was collected from ten plants of each F2:3 family. The plants were watered regularly to ensure that they did not suffer moisture stress.
The virulent RWA colony that had earlier been biotyped by Malinga et al. (2007b) was collected from symptomatic bread wheat in the screenhouse and multiplied in preparation for infestation. The aphid colony was established on ‘Kenya Kwale’, a wheat variety that is highly susceptible to RWA and maintained in the greenhouse with temperatures 25:18°C, photoperiod (LD 12:12) and relative humidity varying between 60-80%. The pots and evaluation flats were caged with a 60 cm high wire cage and covered with a polyester screen mesh (68 meshes per square cm) to prevent aphids from getting in or escaping. Five adult aphids (3 - 5 instar stage) were placed on the whorls of seedlings at the two leaf stage using a camel hair brush. Five aphids were used for each plant to ensure maximum infestation pressure was achieved. RWA infes-tation was rated at twenty one days after infestation and scoring done according to a modified 1 - 9 visual scale (Malinga, 2007). Plants showing damage scale of 1 - 5 were grouped as resistant and 6 - 9 susceptible.
Statistical analysis
The data of RWA reaction for individual F2 plants was tested against an expected phenotypic segregation ratio of 3:1 using the Chi square (χ2) goodness of fit test, to confirm the mode of inheritance at probability level of P = 0.05. The data on RWA reaction for individual F2:3 families was tested against an expected phenotypic segregation ratio of 1:2:1 using the Chi square (χ2) test to also confirm the mode of inheritance at probability level of P = 0.05. The segregation of F2:3 families was expected to confirm the segregation ratios observed in F2 populations and aid in the classification of F2 lines for the bulk segregant analysis.
Genotyping using microsatellite markers
DNA was isolated from parents and 100 F2 plants following the protocol by Dellaporta and Woods (1983) with some modifications. Approximately 500 mg of leaf tissue was ground with liquid nitrogen before adding and mixing with 500 µl of extraction buffer (0.1 M Tris-HCl pH 8.0, 0.05 M ethylenediaminetetraacetic acid (EDTA), 0.5 M NaCl, 1% polyvinylpyrolidone, 1.6% sodium dodecyl sulphate (SDS). This was followed by the addition of 50 µl of 20% SDS, and after mixing by inversion the tubes were incubated for 15 min at 65°C. The samples were removed from incubator and 250 ml of potassium acetate (-20°C) followed by incubation in freezer for 10 min at -20°C. The samples were then centrifuged at 13,000 rpm for 5 min and 500 µl of isopropanol (at -20°C) was added to the supernatant in new tubes. The mixture was incubated for 10 min at -20°C followed by centrifugation at 13,000 rpm for 5 min. The supernatant was discarded, DNA pellet washed with 500 µl of 70% ethanol (at -20°C) followed by air-drying. The DNA pellet was resuspended in 100 ul of 10:1 TE (10 mM Tris:1 mM EDTA) buffer. The samples were RNase treated by adding 2.25 μL of 10 mg ml-1 RNase and incubating for 30 min at 65°C followed by storing at -20°C till further use.
DNA was quantified spectrophotometrically and quality checked by 1% agarose gel electrophoresis, against lambda DNA of known quantity. Presence of DNA was confirmed by visualizing the bands on the gel under a UV transilluminator (Alpha Innotech, Taiwan). Comparison of the concentration of DNA was done against known standards of 100, 125, 250 500 and 1000 ng/µl lambda DNA to determine quantity. DNA was diluted to a working stock of 30 ng/μl for PCR reactions.
Bulk segregant analysis (BSA) with microsatellite markers was used to identify DNA markers associated with RWA resistance. Nine primers for Xgwm microsatellites were used in this study. These microsatellite markers have been mapped in wheat chromosome 7D. They included Xgwm30, Xgwm44, Xgwm46, Xgwm56, Xgwm111, Xgwm297, Xgwm333, Xgwm437 and Xgwm644 (Roder et al., 1998). BSA was done using DNA from KRWA9, NjoroBW2, resistant homozygous plants, resistant heterozygous (segregating) plants, homozygous susceptible plants and control resistance sources PI 137739 (Dn1), PI 262660 (Dn2), USA9 (Dn7) and PI 294994 (Dn5, Dn8 and Dn9). DNA solution was bulked into their respective resistant and susceptible bulks. The resistant bulk consisted of equal amounts of DNA 10 μl from eight homozygous resistant plants. The susceptible bulk contained DNA from eight susceptible plants. The third bulk contained DNA from segregating plants. There were two more bulks with equal amounts of DNA 10 μl from each parent NjoroBW2 and KRWA9. All PCR reactions were performed in 13 μl reaction volumes containing 1.25 μl of 10X PCR buffer, 8.5 μl of ddH2O, 0.5 μl of 10 mM dNTPs, 0.75 µl of 50 mM MgCl2, 0.25 μl of 10 mM each of forward and reverse primer and 0.05 μl of Invitrogen Taq DNA polymerase recombinant (5 U/µl) and 1.5 μl template DNA. PCR amplifications were carried out on PCR machine (Applied Biosystems 2720 Thermal Cycler, Singapore). The microsatellite products were resolved on 2.0% agarose gels in TAE buffer. The bands were visualized under a UV transilluminator (Alpha Innotech, Taiwan). The electrophoresis products were captured on a camera and transferred to a computer.
Once a specific polymorphism between resistant and susceptible bulks had been identified by BSA screening, individual co-segregation analysis, based on the associations between marker genotype and RWA reaction phenotype, was carried out on the total F2 segregating population to determine the genetic linkage between a RWA resistance gene and a marker.
Microsatellite marker, Xgwm111 (linked to RWA resistance) and weighted at 210 bp was used to confirm the presence of RWA resistance gene in NjoroBW2 × KRWA9 F2 population. The PCR profile was a follows: an initial denaturing step at 94°C for 3 min followed by 45 cycles at 94°C for 1 min, annealing for primer Xgwm111 at 55°C for 1 min.
This was followed by primer elongation at 72°C for 2 min and final 10 min primer extension at 72°C. The simple sequence repeat (SSR) products were resolved on 2.0% agarose gels in TAE buffer and bands visualized under a UV transilluminator (Alpha Innotech, Taiwan). The electrophoresis products were captured on a camera and transferred to a computer.
Marker analysis
Informative bands were scored as present (+) or absent (-) and since SSRs are co-dominant markers, it was expected that alleles from both parents would be observed in some samples. Single marker analysis was done using the JoinMap software (Stam and Van Ooijen 1995) to detect QTL associated with Xgwm111. Linear regression was done to obtain coefficient of determination (R2) that explains the phenotypic variation arising from QTL linked to a marker. Chi-square goodness-of-fit test was carried out to test conformity to Mendelian segregation patterns. The Chi square (χ2) value and segregation ratios from gel data were later compared against Chi square (χ2) value and phenotypic segregation ratios resulting from RWA reactions of individual F2 populations and F2:3 families.
RESULTS
Inheritance analysis
The resistant parent KRWA9 showed resistance reactions having minimal levels of chlorosis and rolling, with damage scores of 1 - 3. This indicated high levels of resistance in the resistant parent. The susceptible parent NjoroBW2 showed a susceptible reaction with damage scores of 7 - 9. Most NjoroBW2 seedlings had severe leaf chlorosis, streaking and rolling leading to death after 21 days of infestation. The F1 population of cross NjoroBW2 × KRWA9 showed resistance reaction with damage scores of 1 - 4. The resistance reaction of F1 population was not significantly different from the reaction of KRWA9 indicating that the resistance gene in KRWA9 is dominant. The χ2 statistics for NjoroBW2 × KRWA9 F1 population was significant at P<0.05 with a fit in ratio of 1:0 (Table 1). In NjoroBW2 × KRWA9 F2 generation, the hybrids segregated and were classified into their respective phenotypic classes. The F2 population showed both susceptible and resistant reactions with damage scores of 1 - 9. The χ2 statistics was significant at P<0.05 with a fit in ratio of 3:1 (Table 1). The F2:3 progenies were classified as homozygous resistant and heterozygous resistant (segregating) based on the seedling reactions to RWA. The F2:3 homozygous resistant progenies showed damage scores of 1 - 5, indicating resistance. Heterozygous resistant progenies showed damage scores of 1 - 9 indicating both resistance and susceptible reactions. The χ2 statistics for F2:3 population of NjoroBW2 × KRWA9 was significant at P<0.05 (Table 1) with a fit in ratio of 1:2:1. These results confirmed the model of 3:1 at F2 populations with a fit of 1:2:1 at F2:3 families for monohybrid inheritance.
.png)
Genotypic analysis
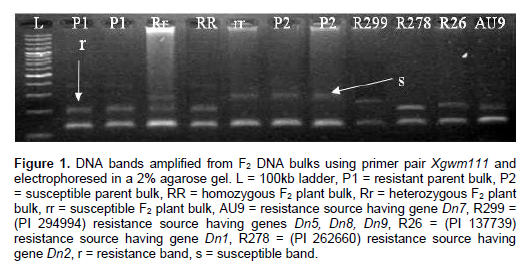
Nine primers (Xgwm30, Xgwm44, Xgwm46, Xgwm56, Xgwm111, Xgwm297, Xgwm333, Xgwm437 and Xgwm644) were screened for polymorphism and only chromosome 7DS primer Xgwm111 produced a distinguishing polymorphism. Primer Xgwm111 produced a band that clearly and consistently differentiated the parents, resistant and susceptible bulks (Figure 1). A band was produced on control resistance source PI 137739 which was similar to the one on resistance source KRWA9. The band was approximately 210 bp and was subsequently tested on F2 population individuals. Other bands were produced on resistance sources PI 262660 (Dn2), PI 294994 (Dn5, Dn8 and Dn9) and AUS9 (Dn7). Figure 1 shows the banding patterns for KRWA9, NjoroBW2, homozygous resistant plants, heterozygous resistant plants, homozygous susceptible plants and control resistance sources “R299”, “R278”, “R26” and “AU9”. KRWA9 showed two distinctive bands; one was 210 bp while the other was 160 bp. The susceptible parent NjoroBW2 showed two distinctive bands; one was 280 bp while the other was 160 bp (Figure 1). It was observed that both parents had a common 160 bp band. The 210 bp band was present in the resistant parent but absent in the susceptible parent. This band was designated as the band of interest. The inclusion of different resistant sources helped to accurately identify the DNA markers for gene of interest. The primer Xgwm111 also produced a 210 bp band that clearly and consistently differentiated the parents, resistant, heterozygous and susceptible plants in the F2 population (Figure 2). Based on the banding patterns observed in the F2 population, 28 plants were homozygous resistant, 49 heterozygous and 23 homozygous susceptible (Table 3). This ratio did not differ from the expected 1:2:1 segregation ratio (χ2 = 5.991, df = 2, P ≤ 0.05).

Linkage analysis
The F2 population of NjoroBW2 × KRWA9 cross showed a wide range of segregation for response to infestation by RWA. The frequency distribution of RWA feeding damage on the F2 population was somewhat bimodal, indicating the presence of one major resistance gene in KRWA9 (Figure 3). Simple regression analysis identified marker Xgwm111 to be highly significantly associated with resistance in KRWA9. The marker had an LOD score of 40.1 and high R2 value of 85% indicating that it is a very significant marker for the resistance in KRWA9 (Table 2). Genetic data for Xgwm111 marker showed a complete co-segregation with the disease data in the mapping population indicating a very tight linkage to the RWA resistance gene in KRWA9.



DISCUSSION
KRWA9 was selected for this study because visual observations of RWA feeding damage on it suggested that this source of resistance has high level resistance (Pathak et al., 2007; Malinga et al., 2008). This resistance could be transferred to NjoroBW2 a popular commercial wheat variety which is susceptible to RWA. The F1 seedlings of the cross between NjoroBW2 and KRWA9 were all resistant indicating the resistance in KRWA9 is dominant. The segregation observed in the F2 population and the F2:3 families further confirmed the dominance of resistance in KRWA9. Most RWA resistant genotypes have single dominant genes located on chromosome 1D and 7D (Du toit, 1987; Nkongolo et al., 1991b; Saidi and Quick, 1996; Liu et al., 2001; Liu, 2001). Resistance sources reported to have single dominant genes include PI137739 (Dn1), PI262660 (Dn2), PI372129 (Dn4) and PI243781 (Dn6) (Du Toit, 1989; Nkongolo et al., 1991b; Saidi and Quick, 1994). The dominant nature of RWA resistance gene could be easily identified in the segregating populations. However, the major problem with single gene inheritance is that insect can develop biotypes very fast if the resistant cultivar is grown on a large scale. Colorado State University has developed several commercially available RWA resistant varieties of winter wheat such as Halt, Prairie Red, Prowers 99 and Yuma (Thomas et al., 2002). All these varieties have the Dn4 resistance gene derived from PI 372129 (Turcikum 57). It was later reported that RWA resistant cultivars with the Dn4 gene were susceptible to a new biotype designated as “Biotype 2” (Haley et al., 2004). This led to sourcing of more resistant materials. Gene Dn7 that was previously transferred from rye to wheat background via a 1 RS/1BL translocation had been reported to be resistant biotype 1 and 2 and depicts high levels of resistance as compared to other Dn genes (Collins et al., 2005; Turanli et al., 2012). However, part of the rye chromosome containing Dn7 has detrimental genes resulting to poor bread making quality (Graybosch et al., 1990). Breeding for resistance with Dn7 gene is no longer a desirable strategy and identification of diverse sources of resistance would be a highly desirable to keep ahead of biotype development in RWA. Pyramiding two or more resistance genes in a single cultivar will also increase the longevity of resistance.
The marker Xgwm111 has previously been found to be linked to genes Dn1, Dn2 and Dn5 in resistance sources PI 137739, PI 262660 and PI 294994, respectively (Liu et al., 2005). In their study, the marker Xgwm111 produced band sizes 210 bp in PI 137739 for gene Dn1, 200 bp in PI 262660 for gene Dn2 and 200 bp in PI294994 for gene Dn5 (Liu et al., 2005). The results are in agreement with Liu et al. (2001, 2002), who reported that Xgwm111 amplifies functional fragments from DNA of RWA-resistant wheat sources with expected sizes of 200 to 225 bp that are associated with RWA resistance.
In the F2 population, marker Xgwm111 followed the expected Mendelian segregation ratio of 3:1 or 1:2:1 (Table 3). These findings are consistent with Pathak et al. (2007) on a single dominant gene controlling resistance in KRWA9. The marker also completely co-segregated with the disease data and it is believed that the resistance gene in KRWA9 must be tightly linked to the marker. This offers a good opportunity for breeders to use this marker to select for resistance to RWA.
CONCLUSION
The usage of host plant resistance at the low cost is environmentally safe and is an ideal method to control the Russian wheat aphid. KRWA9 is a good source of resistance to RWA biotypes in Kenya and marker Xgwm111 could be used for marker assisted selection of resistance associated with this line. Similarity exists between KRWA9 and PI 137739, therefore there is a need to screen more markers in order to find more polymorphic markers in this region of chromosome 7DS. Most RWA resistance sources are monogenic and the challenge is that insects can develop biotypes very fast which could overcome the resistant cultivars. Identification of many sources of RWA resistance would be highly desirable to keep ahead of biotype development in the RWA by way of deploying multiple resistance genes to new breeding lines.
CONFLICT OF INTERESTS
The author(s) have not declared any conflict of interests.
ACKNOWLEDGEMENTS
We would like to thank the Kenya Agricultural Research Institute for facilitating the development, screening and molecular analysis breeding populations. We are grateful to Egerton University for technical support in carrying out this study. Funding of this research was provided by Enhanced Agricultural Productivity Programme (EAPP) through the Kenya Agricultural Research Institute and Murdoch University, Australia.
REFERENCES
| Botha CEJ, Matsiliza B (2006). Reduction in transport in wheat (Triticum aestivum L.) is caused by sustained phloem feeding by the Russian wheat aphid (Duraphis noxia Kudjumov). South Afr. J. Bot. 70:249-254. | ||||
|
Collins MB, Haley SD, Randolph TL, Peairs FB, Rudolph JB (2005). Comparison of Dn4- and Dn7-carrying spring wheat genotypes artificially infested with Russian wheat aphid (Homoptera: Aphididae) Biotype 1. J. Econ. Entomol. 98(5):1698-1703. Crossref |
||||
|
Dellaporta SL, Wood J, Hicks JB (1983). A plant DNA minipreparation: version II. Plant Mol. Biol. Rep. 1:19-21. Crossref |
||||
| Du Toit F, Walters MC (1984). Damage assessment and economic threshold values for chemical control of the Russian wheat aphid, Diuraphis noxia (Mordvilko) on winter wheat, pp. 58-62. In M. C. Walters [ed.] Progress in Russian Wheat Aphid (Diuraphis noxia) research in the Republic of South Africa. Technical communication 191. Department of Agriculture, Republic of South Africa. | ||||
| Du Toit F (1989). Inheritance of resistance in two Triticum aestivum lines to Russian wheat aphid (Homoptera: Aphididae). J. Econ. Entomol. 82:1251-1253. | ||||
|
Ennahli S, El Bouhssini M, Grando S, Anathakrishnan R, Niide T, Starkus L, Starkey S, Smith CM (2009). Comparison of categories of resistance in wheat and barley genotypes against biotype 2 of the Russian wheat aphid, Diuraphis noxia (Kurdjumov). Arthropod Plant Interact. 3:45-53. Crossref |
||||
| Graybosch RA, Peterson CJ, Hansen LW, Mattern PJ (1990). Relationships between protein solubility characteristics, 1BL/1RS, high molecular weight glutenin composition and end use quality in winter wheat germplasm. Cereal chem. 67:342-349. | ||||
| Haley SD, Peairs FB, Walker CB, Rudolph JB, Randolph TL (2004). Occurrence of a new Russian wheat aphid biotype in Colorado Crop Sci. 44:1589-1592. | ||||
| Hewitt PH, van Niekerk GJJ, Walters MC, Kriel CF, Fouche A (1984). Aspects of ecology of the Russian wheat aphid, Diuraphis noxia, in the Bloemfontein district. I. The colonization and infestation of sown wheat, identification of summer hosts and cause of infestation symptoms. In: M. C. Walters (eds) Progress in Russian wheat aphid. Research in the Republic of South Africa. Tech. Commun. Dept Agric Rep. S Africa 191:3-13. | ||||
|
Jankielsohn A (2011). Distribution and diversity of Russian wheat aphid (Hemiptera: Aphididae) biotypes in South Africa and Lesotho. J. Econ. Entomol. 104(5):1736-1741. Crossref |
||||
| Kiplagat OK (2005). The Russian wheat aphid (Diuraphis noxia Mord.). Damage on Kenyan wheat (Triticum aestivum L.) varieties and possible control through resistance breeding. PhD Thesis Wageningen University. ISBN 90-8504-175-9. | ||||
|
Lapitan NLV, Peng J, Sharma V (2007). A high-density map and PCR markers for russian wheat aphid resistance gene Dn7 on chromosome 1RS/1BL. Crop Sci. 47:811-820. Crossref |
||||
|
Liu XM, Smith CM, Friebe BR, Gill BS (2005). Molecular mapping and allelic relationships of Russian wheat aphid - resistance genes. Crop Sci. 45:2273-2280. Crossref |
||||
|
Liu XM, Smith CM, Gill BS (2002). Identification of microsatellite markers linked to Russian wheat aphid resistance genes Dn4 and Dn6. Theor. Appl. Genet. 104:1042-1048. Crossref |
||||
|
Liu XM, Smith CM, Gill BS, Tolmay V (2001). Microsatellite markers linked to six Russian wheat aphid resistance genes in wheat. Theor. Appl. Genet. 102:504-510. Crossref |
||||
| Malinga JN (2007). Studies on Russian wheat aphid (Diuraphis noxia: Kurdjumov; Homoptera) with special empasis to biotypes and host plant resistance in bread wheat (Triticum aestivum) in Kenya. PhD Thesis. Egerton University, Kenya. | ||||
| Malinga JN, Kinyua M, Wanjama J, Kamau A, Awalla J (2007a). Differential Population Increase, Damage and Polymorphism within Kenyan Russian wheat aphid Populations. 10th KARI Biennial Scientific Conference. 13-17 November, 2006. Nairobi, Kenya | ||||
|
Malinga JN, Kinyua MG, Kamau AW, Wanjama JK, Awalla JO, Pathak RS (2007b). Biotyping and Genetic Variation within Tropical Population of Russian Wheat Aphid, Duraphis noxia Kurdjumov (Homoptera: Aphididae) in Kenya. J. Entomol. 4:350-361. Crossref |
||||
| Malinga JN, Kinyua MG, Kamau AW, Wanjama JK, Pathak RS (2008). Characterisation of Bread Wheat Genotypes Resistant To Russian Wheat Aphid in Kenya. E. Afr. Agric. For. J. 74(1-2):51-58. | ||||
| McIntosh RA, Yamazaki Y, Devos K, Dubcovsky J, Rogers WJ, Appels R (2003). Catalogue of gene symbols for wheat. In: Pogna NE, Romano M, Pogna EA, Galterio G (eds) Proceedings of the 10th International Wheat Genetics Symposium. 4:1-34. | ||||
|
Miller CA, Altinkut A, Lapitan NLV (2001). A microsatellite marker for tagging Dn2, a wheat gene conferring resistance to the Russian wheat aphid. Crop Sci. 41:1584-1589. Crossref |
||||
| Miller RH, Haile A (1988). Russian wheat aphid on barley in Ethiopia. Rachis 7:51-53. | ||||
|
Nkongolo KK, Quick JS, Limin AE, Fowler DB (1991a). Sources and inheritance of resistance to Russian wheat aphid in Triticum species amphiploids and Triticum tauschii. Can. J. Plant. Sci. 71:703-708. Crossref |
||||
|
Nkongolo KK, Quick JS, Peairs FB, Meyer, WL (1991b). Inheritance of resistance of PI 372129 to the Russian wheat aphid. Crop Sci. 31:905-907. Crossref |
||||
|
Quick JS, Ellis GE, Normann RM, Stromberger JA, Shanahan JF, Peairs FB, Rudolph JB, Lorenz K (1996). Registration of Halt wheat. Crop Sci. 36(1):210-210. Crossref |
||||
|
Ricciardi M, Tocho E, Tacaliti MS, Vasicek A, Gime DO, Paglione A, Simmonds J, Snape JW, Cakir M, Castro AM (2010). Mapping quantitative trait loci for resistance against Russian wheat aphid (Diuraphis noxia) in wheat (Triticum aestivum L.) Crop Pasture Sci. 61:970-977. Crossref |
||||
| Roder MS, Korzun V, Wendehake K, Plaschke J, Tixier MH, Leroy P, Ganal MW (1998). A microsatellite map of wheat. Genet. 149(4):2007-2023. | ||||
| Saidi A, Quick JS (1994). Inheritance of Russian wheat aphid resistance in four winter wheats. In F.B. Peairs, M.K. Kroening and C.L. Simmons (comps.) Proceedings of the Sixth Russian Wheat Aphid Workshop, Colorado State University, Ft. Collins, CO. pp.126-132. | ||||
|
Saidi A, Quick JS (1996). Inheritance and allelic relationships among Russian wheat aphid resistance genes in winter wheat. Crop Sci. 36:256-258. Crossref |
||||
| Stam P, Van Ooijen JW (1995). JOINMAP version 2.0: software for the calculation of genetic linkage maps. CPRO-DLO, Wageningen. | ||||
| Thomas J, Hein G, Baltensperger D, Nelson L, Haley S (2002). Managing the Russian wheat aphid with resistant wheat varieties. NebFacts (September 2002) Nebraska Cooperative Extension NF96-307. Publ. of Cooperative Extension Institute of agriculture and Natural Resources, University of Nebraska, Lincoln. | ||||
|
Tolmay VL, Lindeque RC Prinsloo GJ (2007). Preliminary evidence of a resistance-breaking biotype of the Russian wheat aphid, Diuraphis noxia (Kurdjumov) (Homoptera: Aphididae), in South Africa. Afr. Entomol. 15:228-230. Crossref |
||||
| Tolmay VL, Prinsloo G, Hatting J (2000). Russian wheat resistant wheat cultivars as the main component of an integrated control program. In: Proceedings of the eleventh regional Wheat workshop for Eastern, Central, and Southern Africa. Addis Ababa, Ethiopia: CIMMYT pp. 190-194. | ||||
|
Tolmay VL, Van Deventer CW (2005). Yield retention of resistant wheat cultivars, severely infested with Russian wheat aphid, Diuraphis noxia (Kurdjumov), in South Africa. S. Afr. J. Plant Soil. 22:246-250. Crossref |
||||
| Turanli F, Ilker E, Ersin Dogan F, Askan L, Istipliler D (2012). Inheritance of Resistance to Russian wheat aphid (Diuraphis noxia Kurdjumov) in bread wheat (Triticum aestivum L.). Turk J. Field Crops. 17(2):171-176. | ||||
| Van Niekerk HA (2001). Southern Africa wheat pool. In: The world wheat book: the history of wheat breeding. Ed. by AP B, WJ A. Lavoisier Publishing, Paris, FR, pp. 923-936. | ||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


